
Identification of Ossnrk1a−1 Regulated Genes Associated with Rice Immunity and Seed Set

 , and
, and 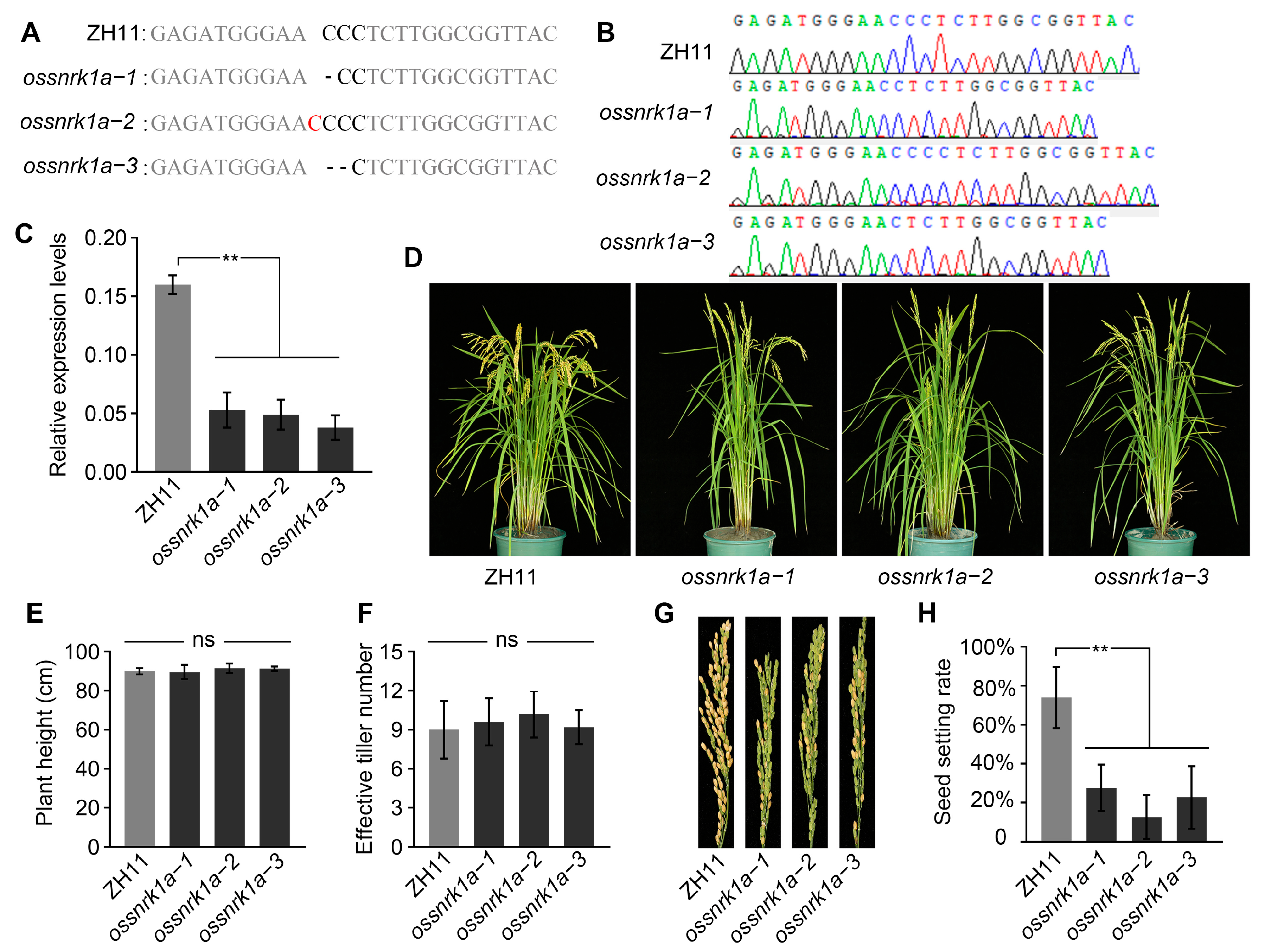{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Deletion of OsSnRK1a Decreased Rice Seed Setting Rate
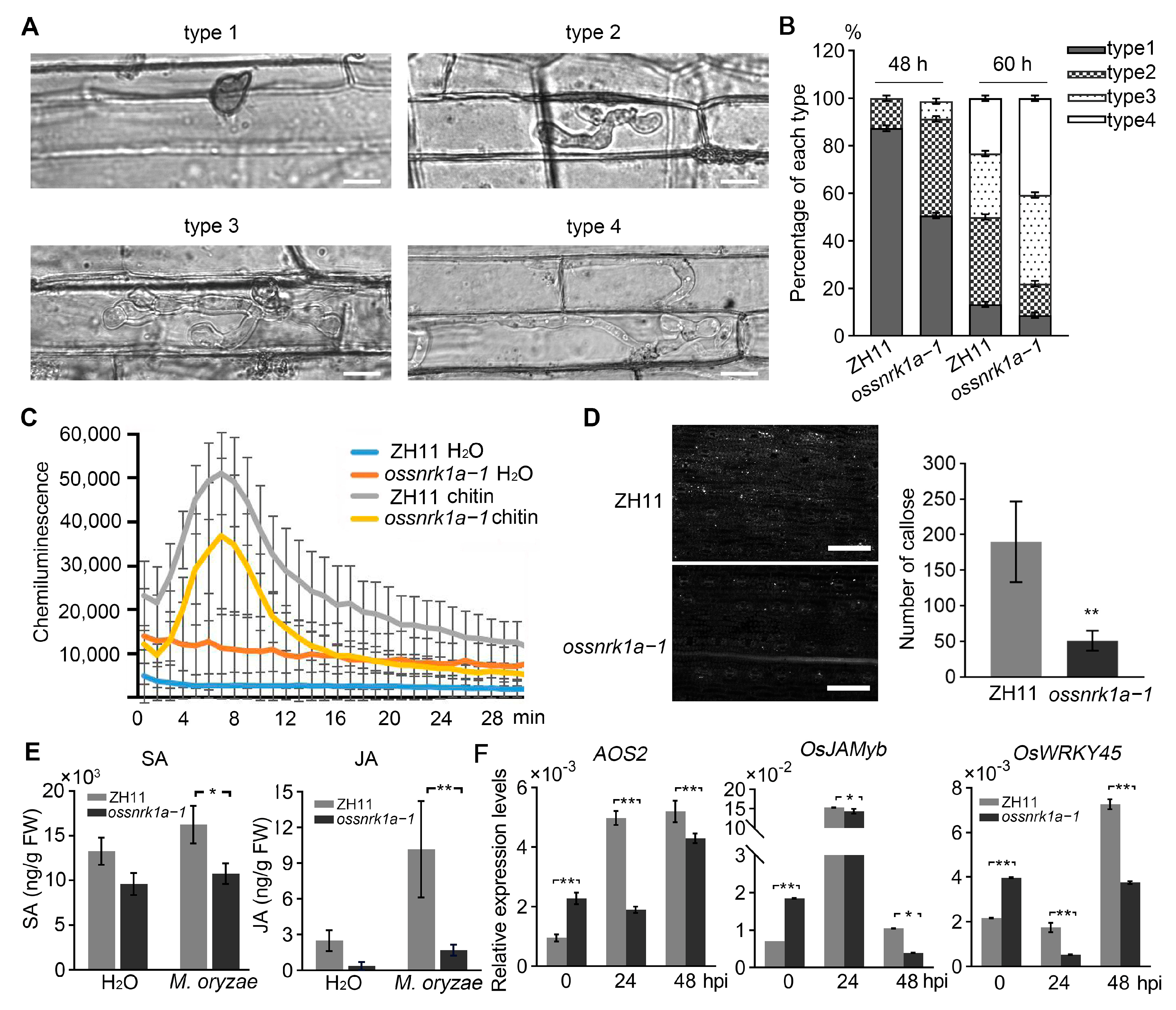
2.2. Knockout of OsSnRK1a Undermined Rice Resistance to Blast Fungus
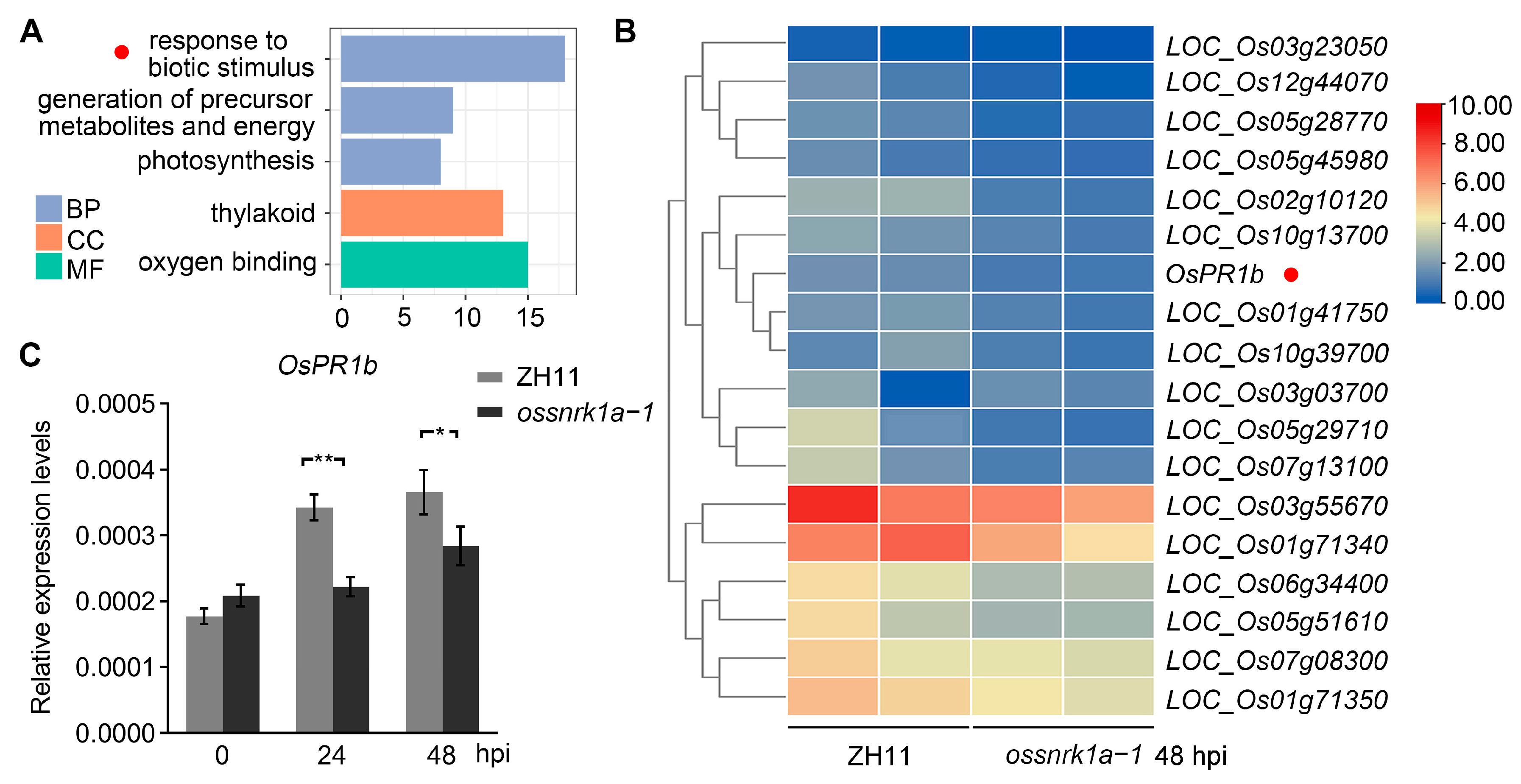
2.3. Transcriptome Analyses Showing Defective OsPR1b Induction by M. oryzae Infection in ossnrk1a−1
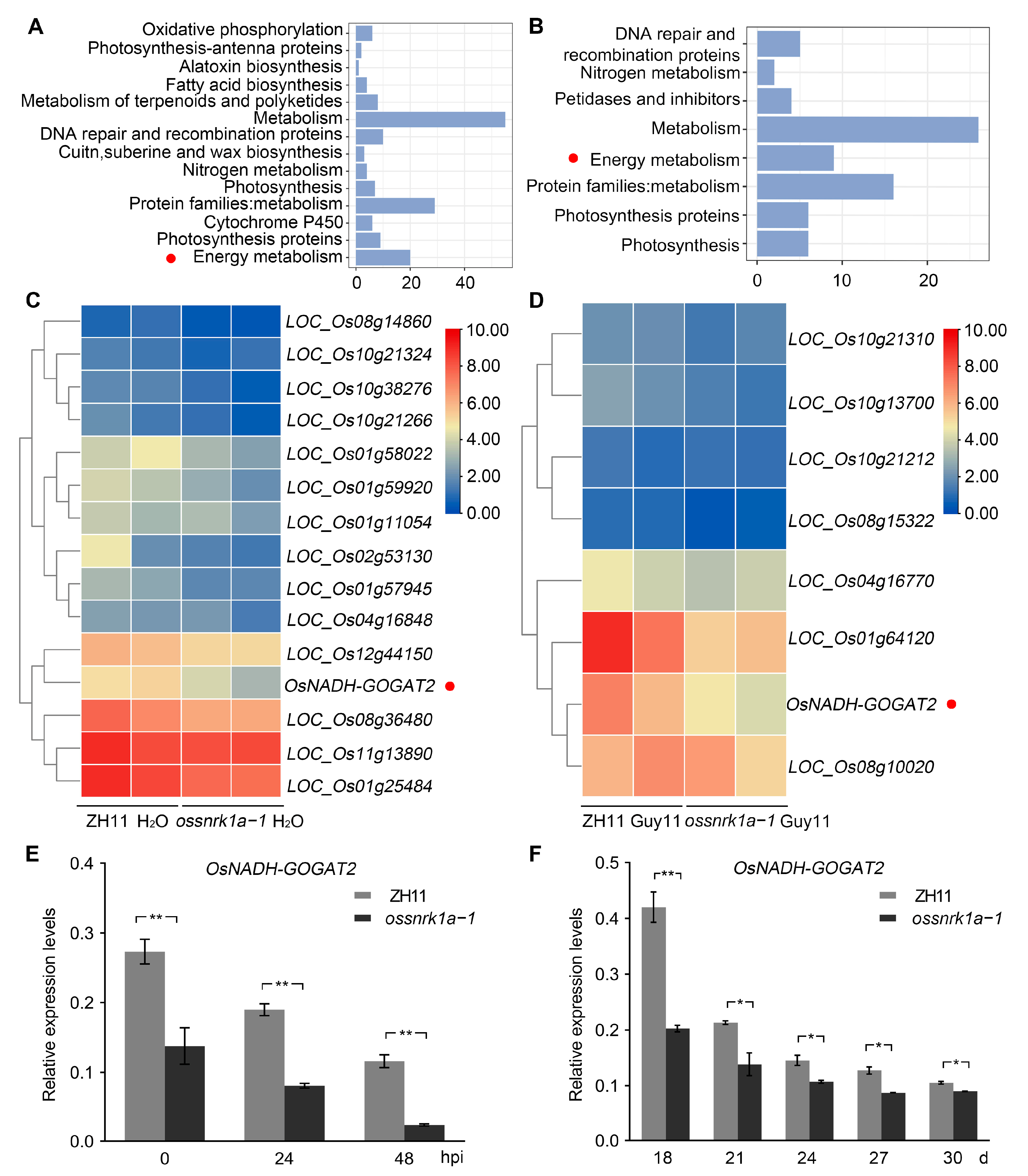
2.4. OsSnRK1a Is a Positive Regulator in OsNADH-GOGAT2 Expression
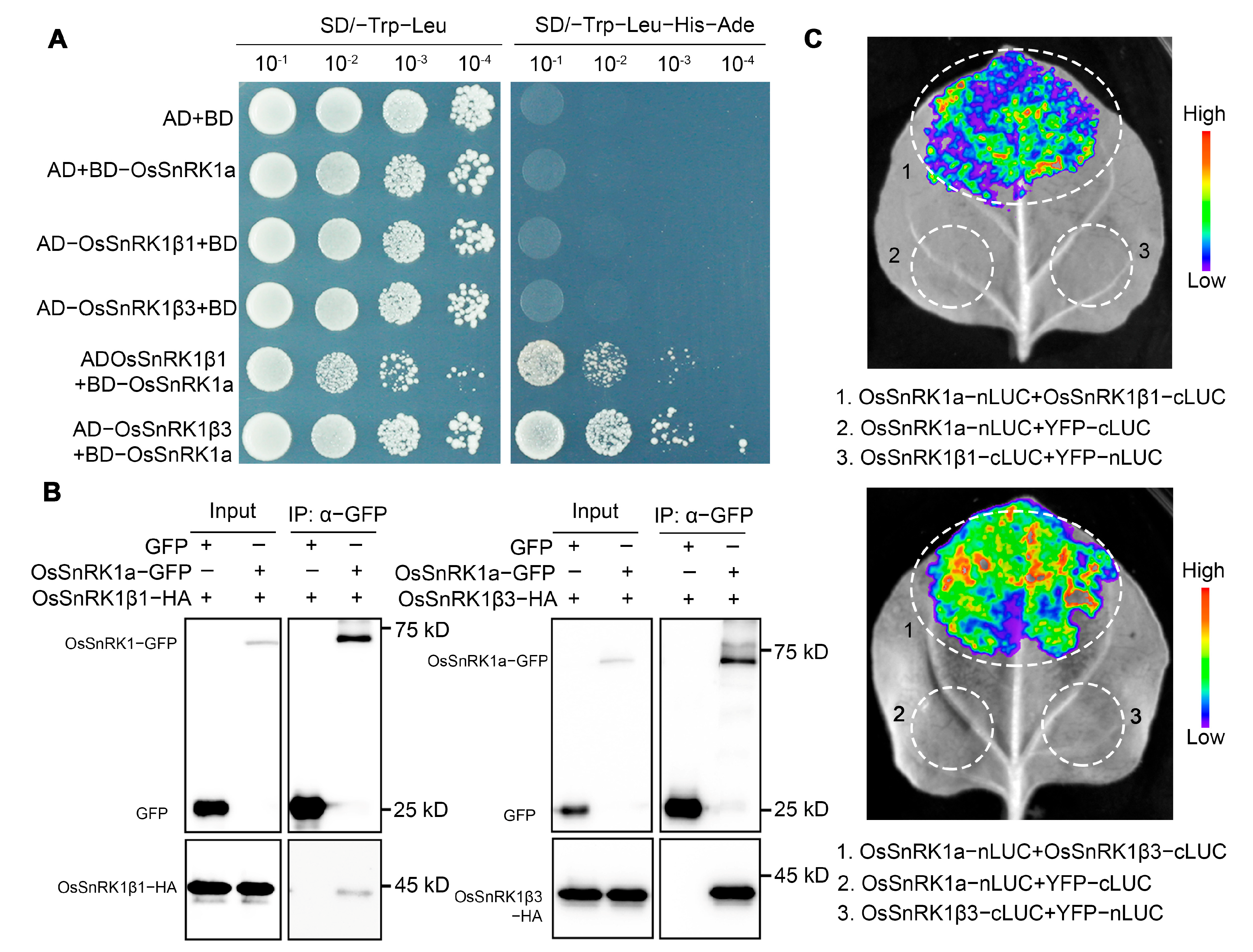
2.5. OsSnRK1a Interacts with the Regulatory Subunits OsSnRK1β1 and OsSnRK1β3
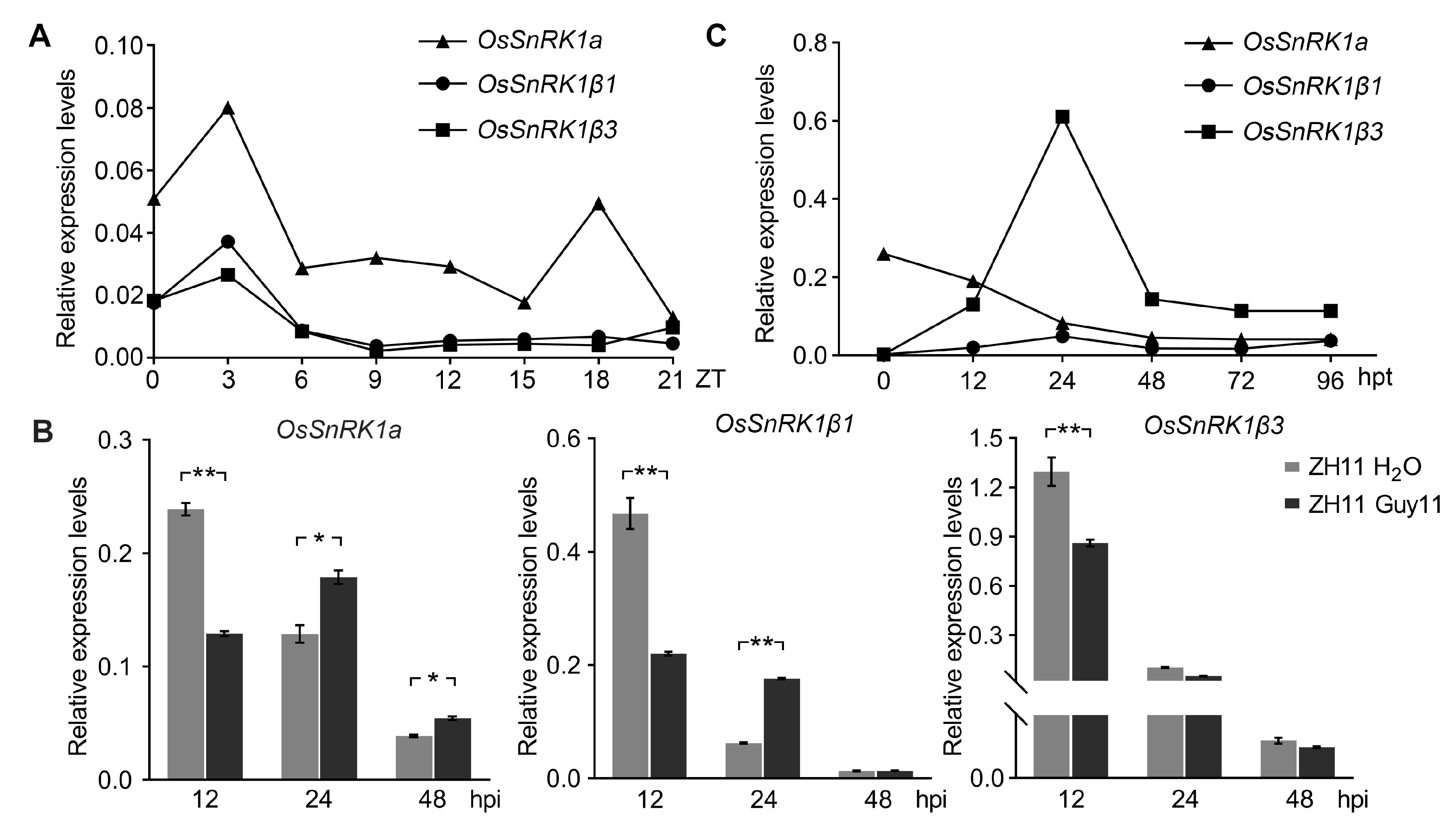
2.6. Transcriptional Pattern Analysis for the SnRK1 Subunits
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Gene Expression Analyses
4.3. Blast Fungus Inoculation
4.4. Measurement of SA and JA Contents
4.5. Rice Sheath Inoculation Assays
4.6. ROS Burst Detection
4.7. Callose Deposition Assay
4.8. Yeast Two-Hybrid Analyses
4.9. Split-Luciferase Complementation Assay
4.10. Co-IP Assays
4.11. Transcriptome Deep Sequencing and Data Analysis
4.12. Starvation Treatment of Seedlings
4.13. Statistical Analyses
4.14. Primers and Plasmid Constructs
4.15. Gene Accession Numbers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| AOS2 | Allene oxide synthase 2 |
| BP | Biological processes |
| CC | Cellular components |
| Co-IP | Co-immunoprecipitation |
| DEGs | Differently expressed genes |
| GFP | Green fluorescence protein |
| GO | Gene Ontology |
| ha. | Hectare |
| hpi | Hours post inoculation |
| IH | Invasive hyphae |
| JA | Jasmonic acid |
| MF | Molecular functions |
| PAMP | Pathogen-associated molecular pattern |
| PTI | PAMP-triggered immunity |
| qRT-PCR | Real-time quantitative reverse transcription PCR |
| ROS | Reactive oxygen species |
| SA | Salicylic acid |
| SnRK1 | Sucrose non-fermenting-1-related protein kinase-1 |
| Y2H | Yeast two-hybrid |
| YFP | Yellow fluorescent protein |
| ZT | Zeitgeber time |
References
- Zhang, M.; Zhang, S. Mitogen-activated protein kinase cascades in plant signaling. J. Integr. Plant Biol. 2022, 64, 301–341. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, K.; Wang, J.; Gong, Z.; Li, S.; Deng, X.; Li, Y. Integrated transcriptomic and metabolomic analyses uncover the differential mechanism in saline–alkaline tolerance between Indica and Japonica rice at the seedling stage. Int. J. Mol. Sci. 2023, 24, 12387. [Google Scholar] [CrossRef]
- Nadarajah, K.K. ROS homeostasis in abiotic stress tolerance in plants. Int. J. Mol. Sci. 2020, 21, 5208. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Singh, S.K. Chapter 18—Role of plant hormones in combating biotic stress in plants. In Plant Perspectives to Global Climate Changes, 1st ed.; Aftab, T., Roychoudhury, A., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 373–391. [Google Scholar]
- Alderson, A.; Sabelli, P.A.; Dickinson, J.R.; Cole, D.; Richardson, M.; Kreis, M.; Shewry, P.R.; Halford, N.G. Complementation of snf1, a mutation affecting global regulation of carbon metabolism in yeast, by a plant protein kinase cDNA. Proc. Natl. Acad. Sci. USA 1991, 88, 8602–8605. [Google Scholar] [CrossRef] [PubMed]
- Crozet, P.; Margalha, L.; Confraria, A.; Rodrigues, A.; Martinho, C.; Adamo, M.; Elias, C.A.; Baena-Gonzalez, E. Mechanisms of regulation of SNF1/AMPK/SnRK1 protein kinases. Front. Plant Sci. 2014, 5, 190. [Google Scholar] [CrossRef] [PubMed]
- Crepin, N.; Rolland, F. SnRK1 activation, signaling, and networking for energy homeostasis. Curr. Opin. Plant Biol. 2019, 51, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Emanuelle, S.; Doblin, M.S.; Stapleton, D.I.; Bacic, A.; Gooley, P.R. Molecular insights into the enigmatic metabolic regulator, SnRK1. Trends Plant Sci. 2016, 21, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.-F.; Cui, N.; Wang, L.; Zhao, X.-C.; Qu, B.; Li, T.-L.; Zhang, G.-L. The SnRK protein kinase family and the function of snRK1 protein kinase. Int. J. Agric. Biol. 2012, 4, 575–579. [Google Scholar]
- Margalha, L.; Valerio, C.; Baena-Gonzalez, E. Plant SnRK1 kinases: Structure, regulation, and function. Exp. Suppl. 2016, 107, 403–438. [Google Scholar] [CrossRef]
- Kulik, A.; Wawer, I.; Krzywinska, E.; Bucholc, M.; Dobrowolska, G. SnRK2 protein kinases-key regulators of plant response to abiotic stresses. OMICS 2011, 15, 859–872. [Google Scholar] [CrossRef]
- Belda-Palazon, B.; Adamo, M.; Valerio, C.; Ferreira, L.J.; Confraria, A.; Reis-Barata, D.; Rodrigues, A.; Meyer, C.; Rodriguez, P.L.; Baena-Gonzalez, E. A dual function of SnRK2 kinases in the regulation of SnRK1 and plant growth. Nat. Plants 2020, 6, 1345–1353. [Google Scholar] [CrossRef]
- Coello, P.; Hey, S.J.; Halford, N.G. The sucrose non-fermenting-1-related (SnRK) family of protein kinases: Potential for manipulation to improve stress tolerance and increase yield. J. Exp. Bot. 2011, 62, 883–893. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.H.; Hong, J.W.; Kim, E.C.; Yoo, S.D. Regulatory functions of SnRK1 in stress-responsive gene expression and in plant growth and development. Plant Physiol. 2012, 158, 1955–1964. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.R.; Lee, K.W.; Chen, C.Y.; Hong, Y.F.; Chen, J.L.; Lu, C.A.; Chen, K.T.; Ho, T.H.; Yu, S.M. SnRK1a–interacting negative regulators modulate the nutrient starvation signaling sensor SnRK1 in source-sink communication in cereal seedlings under abiotic stress. Plant Cell 2014, 26, 808–827. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.-A.; Lin, C.-C.; Lee, K.-W.; Chen, J.-L.; Huang, L.-F.; Ho, S.-L.; Liu, H.-J.; Hsing, Y.L.; Yu, S.M. The SnRK1a protein kinase plays a key role in sugar signaling during germination and seedling growth of rice. Plant Cell 2007, 19, 2484–2499. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, B.; Moraes, T.A.; Mengin, V.; Margalha, L.; Vicente, R.; Feil, R.; Hohne, M.; Sousa, A.; Lilue, J.; Stitt, M.; et al. Impact of the SnRK1 protein kinase on sucrose homeostasis and the transcriptome during the diel cycle. Plant Physiol. 2021, 187, 1357–1373. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, B.; Baena-Gonzalez, E. Management of plant central metabolism by SnRK1 protein kinases. J. Exp. Bot. 2022, 73, 7068–7082. [Google Scholar] [CrossRef] [PubMed]
- Jossier, M.; Bouly, J.P.; Meimoun, P.; Arjmand, A.; Lessard, P.; Hawley, S.; Grahame, H.D.; Thomas, M. SnRK1 (SNF1-related kinase 1) has a central role in sugar and ABA signalling in Arabidopsis thaliana. Plant J. 2009, 59, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Primavesi, L.F.; Jhurreea, D.; Andralojc, P.J.; Mitchell, R.A.; Powers, S.J.; Schluepmann, H.; Delatte, T.; Wingler, A.; Paul, M.J. Inhibition of SNF1-related protein kinase1 activity and regulation of metabolic pathways by trehalose-6-phosphate. Plant Physiol. 2009, 149, 1860–1871. [Google Scholar] [CrossRef]
- Wurzinger, B.; Nukarinen, E.; Nagele, T.; Weckwerth, W.; Teige, M. The SnRK1 kinase as central mediator of energy signaling between different organelles. Plant Physiol. 2018, 176, 1085–1094. [Google Scholar] [CrossRef]
- Piattoni, C.V.; Bustos, D.M.; Guerrero, S.A.; Iglesias, A.A. Nonphosphorylating glyceraldehyde-3-phosphate dehydrogenase is phosphorylated in wheat endosperm at serine-404 by an SNF1-related protein kinase allosterically inhibited by ribose-5-phosphate. Plant Physiol. 2011, 156, 1337–1350. [Google Scholar] [CrossRef]
- Meng, D.; Cao, H.; Yang, Q.; Zhang, M.; Borejsza-Wysocka, E.; Wang, H.; Dandekar, A.M.; Fei, Z.; Cheng, L. SnRK1 kinase-mediated phosphorylation of transcription factor bZIP39 regulates sorbitol metabolism in apple. Plant Physiol. 2023, 192, 2123–2142. [Google Scholar] [CrossRef] [PubMed]
- Kulma, A.; Villadsen, D.; Campbell, D.G.; Meek, S.E.; Harthill, J.E.; Nielsen, T.H.; MacKintosh, C. Phosphorylation and 14-3-3 binding of Arabidopsis 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase. Plant J. 2004, 37, 654–667. [Google Scholar] [CrossRef]
- Hu, Y.; Liu, J.; Lin, Y.; Xu, X.; Xia, Y.; Bai, J.; Yu, Y.; Xiao, F.; Ding, Y.; Ding, C.; et al. Sucrose nonfermenting-1-related protein kinase 1 regulates sheath-to-panicle transport of nonstructural carbohydrates during rice grain filling. Plant Physiol. 2022, 189, 1694–1714. [Google Scholar] [CrossRef] [PubMed]
- Lehti-Shiu, M.D.; Shiu, S. Diversity, classification and function of the plant protein kinase superfamily. Philos. Trans. R. SOC. B 2012, 367, 2619–2639. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Nie, B.; Yu, B.; Xu, F.; Zhang, Q.; Wang, Y.; Xu, W. Rice ubiquitin-conjugating enzyme OsUbc13 negatively regulates immunity against pathogens by enhancing the activity of OsSnRK1a. Plant Biotechnol. J. 2023, 21, 1590–1610. [Google Scholar] [CrossRef]
- Han, X.; Zhang, L.; Zhao, L.; Xue, P.; Qi, T.; Zhang, C.; Yuan, H.; Zhou, L.; Wang, D.; Qiu, J.; et al. SnRK1 phosphorylates and destabilizes WRKY3 to enhance barley immunity to powdery mildew. Plant Commun. 2020, 1, 100083. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Hanley-Bowdoin, L. SnRK1: A versatile plant protein kinase that limits geminivirus infection. Curr. Opin. Virol. 2021, 47, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Li, Y.; Yan, R.; Ren, L.; Liu, F.; Zeng, L.; Sun, S.; Yang, H.; Chen, K.; Xu, L.; et al. SnRK1.1-mediated resistance of Arabidopsis thaliana to clubroot disease is inhibited by the novel Plasmodiophora brassicae effector PBZF1. Mol. Plant Pathol. 2021, 22, 1057–1069. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, G. Plant innate immunity in rice: A defense against pathogen infection. Natl. Sci. Rev. 2016, 3, 295–308. [Google Scholar] [CrossRef]
- Takano, M.; Kajiya-Kanegae, H.; Funatsuki, H.; Kikuchi, S. Rice has two distinct classes of protein kinase genes related to SNF1 of Saccharomyces cerevisiae, which are differently regulated in early seed development. Mol. Gen. Genet. 1998, 260, 388–394. [Google Scholar] [CrossRef]
- Filipe, O.; De Vleesschauwer, D.; Haeck, A.; Demeestere, K.; Hofte, M. The energy sensor OsSnRK1a confers broad-spectrum disease resistance in rice. Sci. Rep. 2018, 8, 3864. [Google Scholar] [CrossRef]
- Kim, C.; Vo, K.T.X.; An, G.; Jeon, J. A rice sucrose non-fermenting-1 related protein kinase 1, OSK35, plays an important role in fungal and bacterial disease resistance. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 669–675. [Google Scholar] [CrossRef]
- Nukarinen, E.; Nagele, T.; Pedrotti, L.; Wurzinger, B.; Mair, A.; Landgraf, R.; Bornke, F.; Hanson, J.; Teige, M.; Baena-Gonzalez, E.; et al. Quantitative phosphoproteomics reveals the role of the AMPK plant ortholog SnRK1 as a metabolic master regulator under energy deprivation. Sci. Rep. 2016, 6, 31697. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.B.; Lee, B.C.; Lee, D.E.; Kuk, Y.I.; Lee, A.Y.; Han, O.; Back, K. Molecular characterization of the gene encoding rice allene oxide synthase and its expression. Biosci. Biotechnol. Biochem. 2002, 66, 2719–2722. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.; Liu, H.; Qiu, D.; Zhou, Y.; Li, X.; Xu, C.; Wang, S. A pair of allelic WRKY genes play opposite roles in rice-bacteria interactions. Plant Physiol. 2009, 151, 936–948. [Google Scholar] [CrossRef] [PubMed]
- Shimono, M.; Koga, H.; Akagi, A.; Hayashi, N.; Goto, S.; Sawada, M.; Kurihara, T.; Matsushita, A.; Sugano, S.; Jiang, C.J.; et al. Rice WRKY45 plays important roles in fungal and bacterial disease resistance. Mol. Plant Pathol. 2012, 13, 83–94. [Google Scholar] [CrossRef]
- Cao, W.; Chu, R.; Zhang, Y.; Luo, J.; Su, Y.; Xie, L.; Zhang, H.; Wang, J.; Bao, Y. OsJAMyb, a R2R3-type MYB transcription factor, enhanced blast resistance in transgenic rice. Physiol. Mol. Plant Pathol. 2015, 92, 154–160. [Google Scholar] [CrossRef]
- Agrawal, G.K.; Rakwal, R.; Jwa, N.S. Rice (Oryza sativa L.) OsPR1b gene is phytohormonally regulated in close interaction with light signals. Biochem. Biophys. Res. Commun. 2000, 278, 290–298. [Google Scholar] [CrossRef]
- Luan, Z.H.; Zhou, D.W. Screening of rice (Oryza sativa L.) OsPR1b-interacting factors and their roles in resisting bacterial blight. Genet. Mol. Res. 2015, 14, 1868–1874. [Google Scholar] [CrossRef]
- Tabuchi, M.; Abiko, T.; Yamaya, T. Assimilation of ammonium ions and reutilization of nitrogen in rice (Oryza sativa L.). J. Exp. Bot. 2007, 58, 2319–2327. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhao, J.; Wang, X.; Ma, L.; Ma, Z.; Meng, X.; Fan, H. SnRK1 signaling regulates cucumber growth and resistance to Corynespora cassiicola. Plant Sci. 2023, 332, 111716. [Google Scholar] [CrossRef] [PubMed]
- Tamura, W.; Kojima, S.; Toyokawa, A.; Watanabe, H.; Tabuchi-Kobayashi, M.; Hayakawa, T.; Yamaya, T. Disruption of a novel NADH-glutamate synthase2 gene caused marked reduction in spikelet number of rice. Front. Plant Sci. 2011, 2, 57. [Google Scholar] [CrossRef]
- Ljung, K.; Nemhauser, J.L.; Perata, P. New mechanistic links between sugar and hormone signalling networks. Curr. Opin. Plant Biol. 2015, 25, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Sugden, C.; Donaghy, P.G.; Halford, N.G.; Hardie, D.G. Two SNF1-related protein kinases from spinach leaf phosphorylate and inactivate 3-hydroxy-3-methylglutaryl-coenzyme a reductase, nitrate reductase, and sucrose phosphate synthase in vitro. Plant Physiol. 1999, 120, 257–274. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Chao, Y.C.; Tseng, T.W.; Huang, C.K.; Lo, P.C.; Lu, C.A. Two MYB-related transcription factors play opposite roles in sugar signaling in Arabidopsis. Plant Mol. Biol. 2017, 93, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Hulsmans, S.; Rodriguez, M.; De Coninck, B.; Rolland, F. The SnRK1 energy sensor in plant biotic interactions. Trends Plant Sci. 2016, 21, 648–661. [Google Scholar] [CrossRef]
- Bhargava, A.; Mansfield, S.D.; Hall, H.C.; Douglas, C.J.; Ellis, B.E. MYB75 functions in regulation of secondary cell wall formation in the Arabidopsis inflorescence stem. Plant Physiol. 2010, 154, 1428–1438. [Google Scholar] [CrossRef]
- Gonzalez, A.; Zhao, M.; Leavitt, J.M.; Lloyd, A.M. Regulation of the anthocyanin biosynthetic pathway by the TTG1/bHLH/Myb transcriptional complex in Arabidopsis seedlings. Plant J. 2008, 53, 814–827. [Google Scholar] [CrossRef]
- Emanuelle, S.; Doblin, M.S.; Gooley, P.R.; Gentry, M.S. The UBA domain of SnRK1 promotes activation and maintains catalytic activity. Biochem. Biophys. Res. Commun. 2018, 497, 127–132. [Google Scholar] [CrossRef]
- Wang, C.; Shen, L.; Fu, Y.; Yan, C.; Wang, K. A simple CRISPR/Cas9 system for multiplex genome editing in rice. J. Genet. Genom. 2015, 42, 703–706. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Yang, F.; Niu, Y.; Lin, Y.; Chen, Z.; Li, G.; Luo, Q.; Wang, F.; Wang, M. Loss function of SL (sekiguchi lesion) in the rice cultivar Minghui 86 leads to enhanced resistance to (hemi)biotrophic pathogens. BMC Plant Biol. 2020, 20, 507. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Huang, X.; He, Z.; Zhang, Q.; Meng, H.; Shi, H.; Feng, B.; Zhou, Y.; Zhang, J.; Lu, G.; et al. Phosphorylation of OsTGA5 by casein kinase II compromises its suppression of defense-related gene transcription in rice. Plant Cell 2022, 34, 3425–3442. [Google Scholar] [CrossRef]
- Yang, C.; Yu, Y.; Huang, J.; Meng, F.; Pang, J.; Zhao, Q.; Islam, M.A.; Xu, N.; Tian, Y.; Liu, J. Binding of the Magnaporthe oryzae chitinase MoChia1 by a rice tetratricopeptide repeat protein allows free chitin to trigger immune responses. Plant Cell 2019, 31, 172–188. [Google Scholar] [CrossRef]
- Schneider, C.; Kudla, G.; Wlotzka, W.; Tuck, A.; Tollervey, D. Transcriptome-wide analysis of exosome targets. Mol. Cell 2012, 48, 422–433. [Google Scholar] [CrossRef]
- Mahmood, T.; Yang, P.C. Western blot: Technique, theory, and trouble shooting. N. Am. J. Med. Sci. 2012, 4, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Y.; Lu, M.; Chen, J.; Li, W.; Wang, M.; Chen, F. Identification of Ossnrk1a−1 Regulated Genes Associated with Rice Immunity and Seed Set. Plants 2024, 13, 596. https://doi.org/10.3390/plants13050596
Cao Y, Lu M, Chen J, Li W, Wang M, Chen F. Identification of Ossnrk1a−1 Regulated Genes Associated with Rice Immunity and Seed Set. Plants. 2024; 13(5):596. https://doi.org/10.3390/plants13050596
Chicago/Turabian StyleCao, Yingying, Minfeng Lu, Jinhui Chen, Wenyan Li, Mo Wang, and Fengping Chen. 2024. "Identification of Ossnrk1a−1 Regulated Genes Associated with Rice Immunity and Seed Set" Plants 13, no. 5: 596. https://doi.org/10.3390/plants13050596