
The Effects of Light Spectrum and Intensity, Seeding Density, and Fertilization on Biomass, Morphology, and Resource Use Efficiency in Three Species of Brassicaceae Microgreens


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Context
2.2. Light Conditions
2.3. Environmental Conditions
2.4. Measurements and Calculations
2.5. Statistical Analysis
3. Results
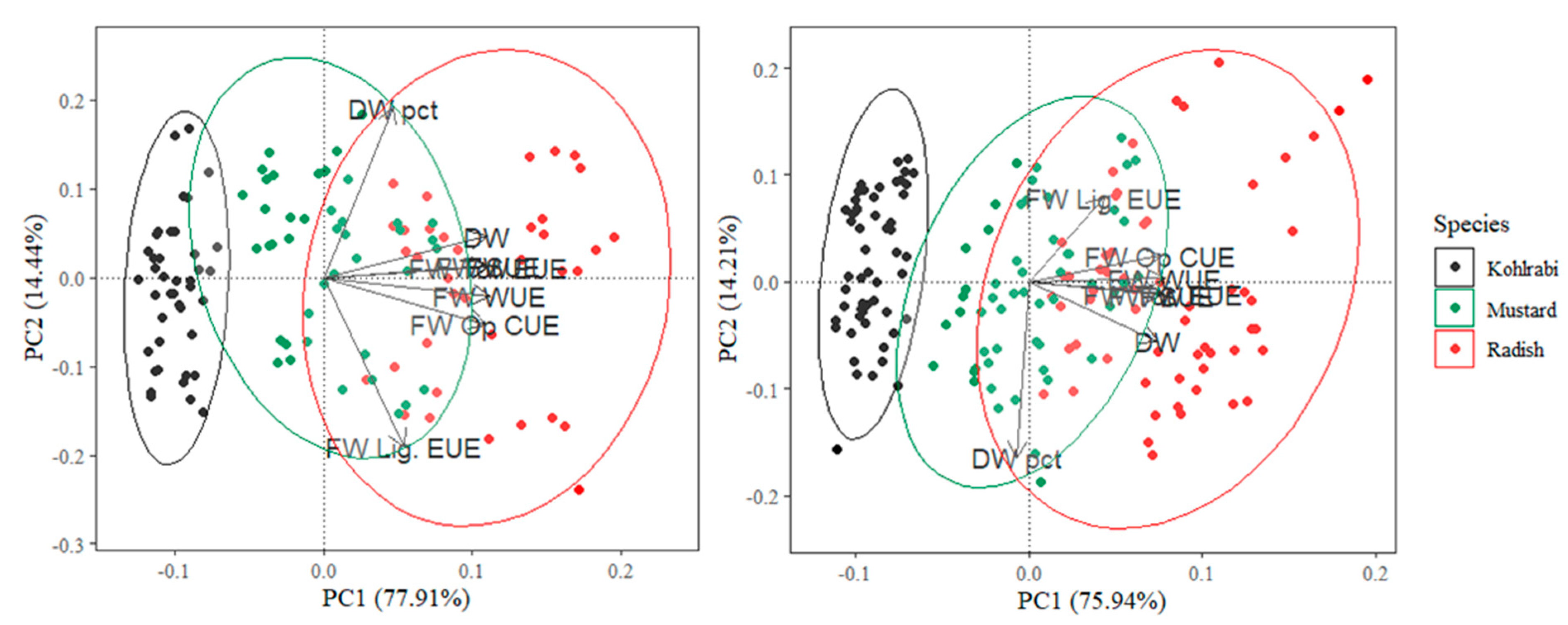
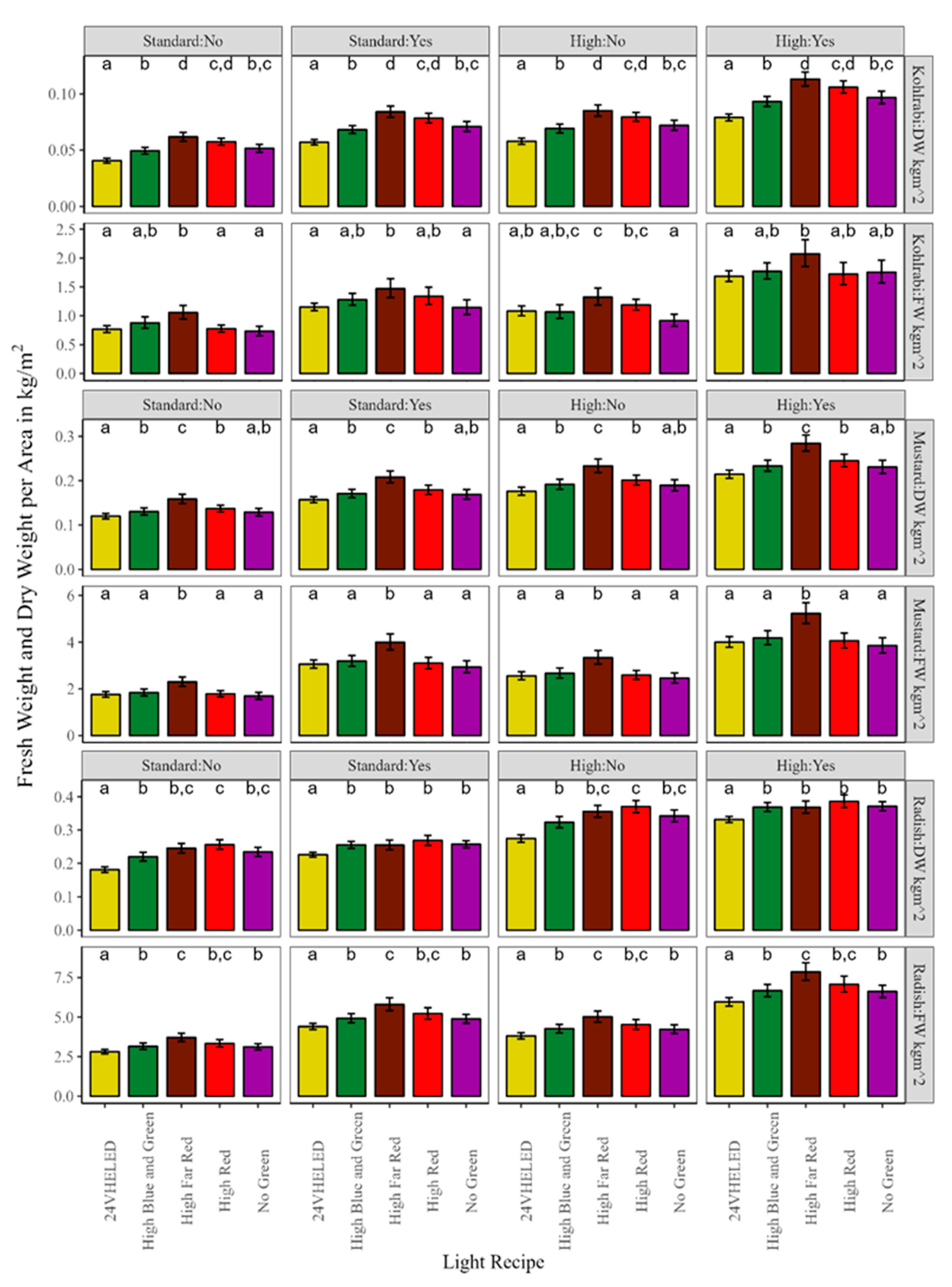
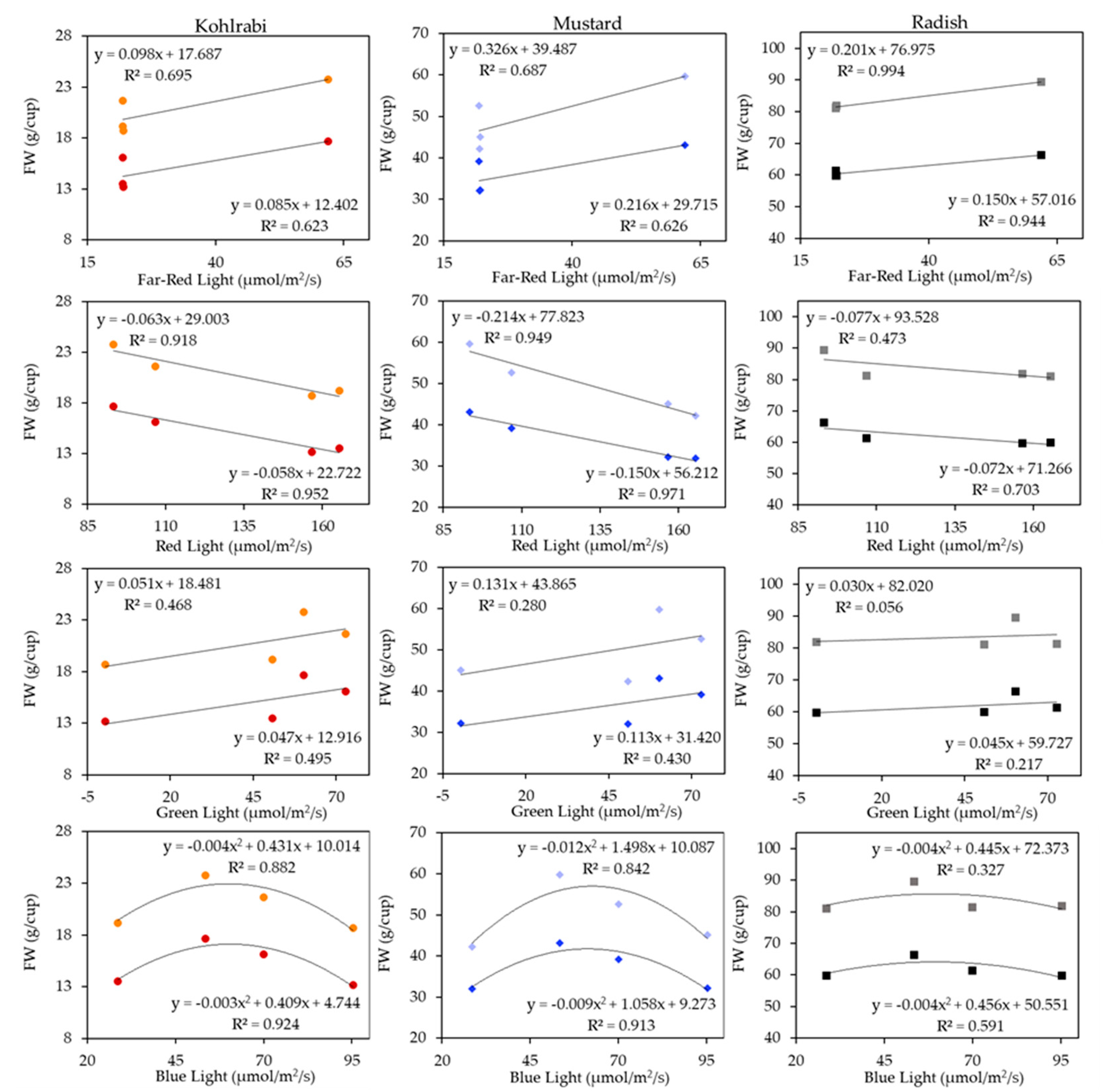
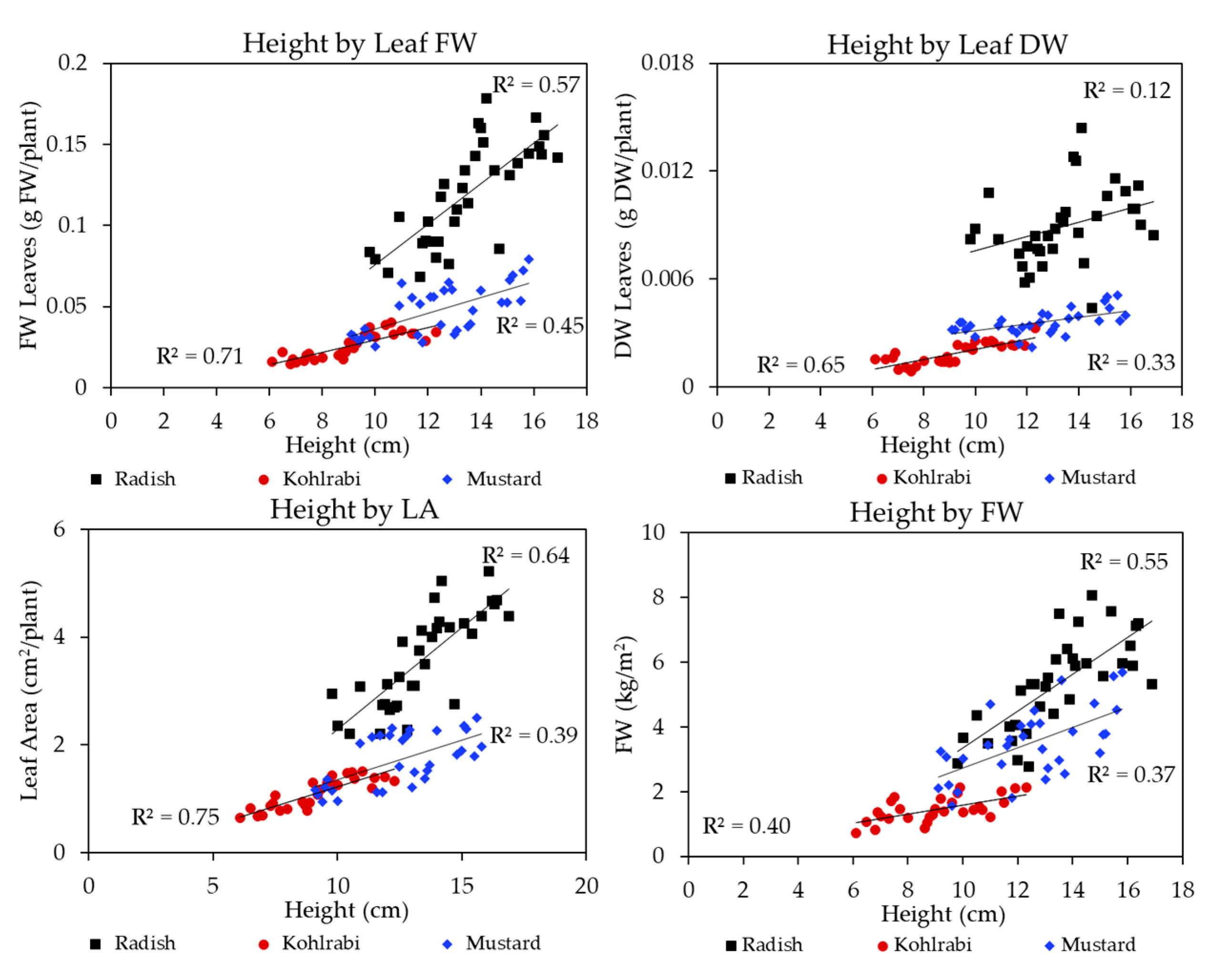
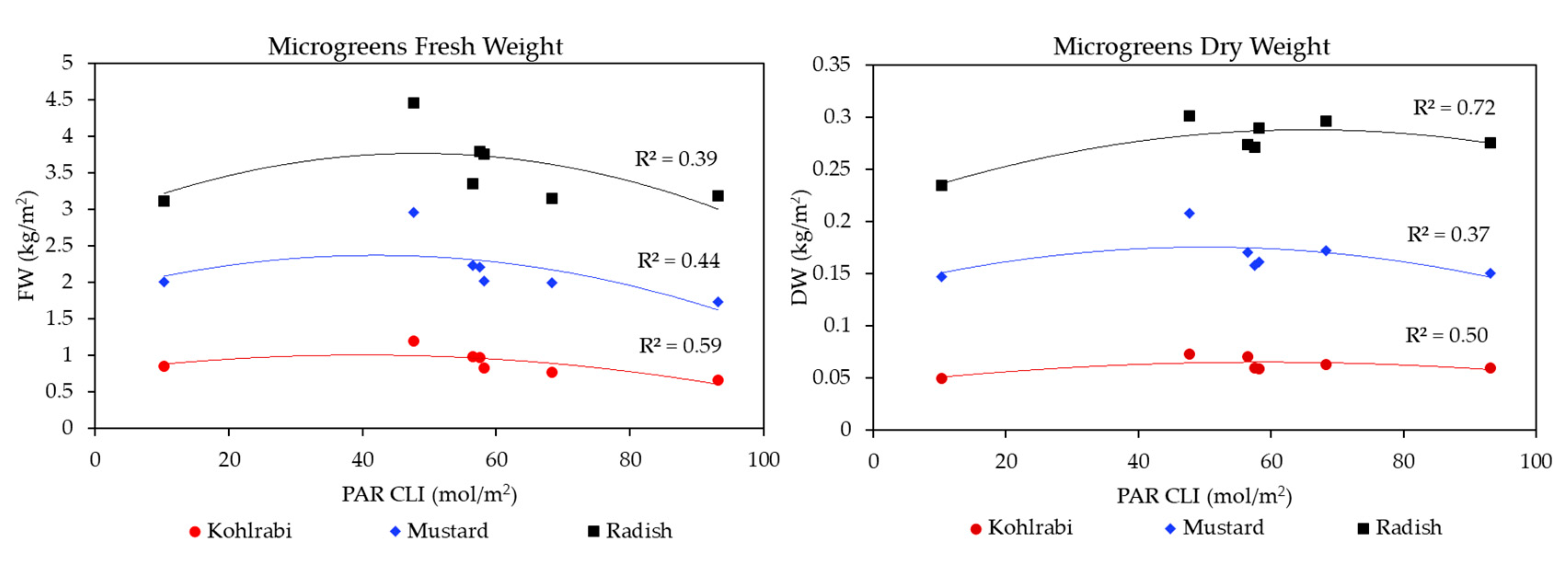
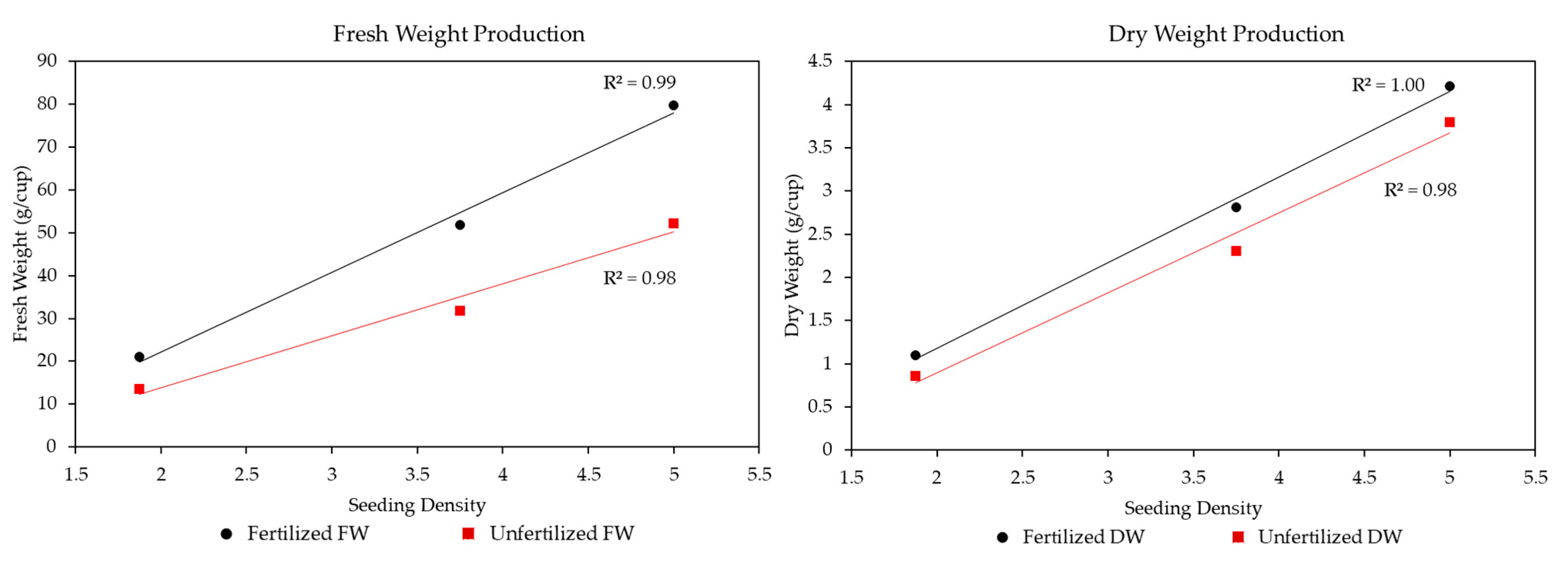
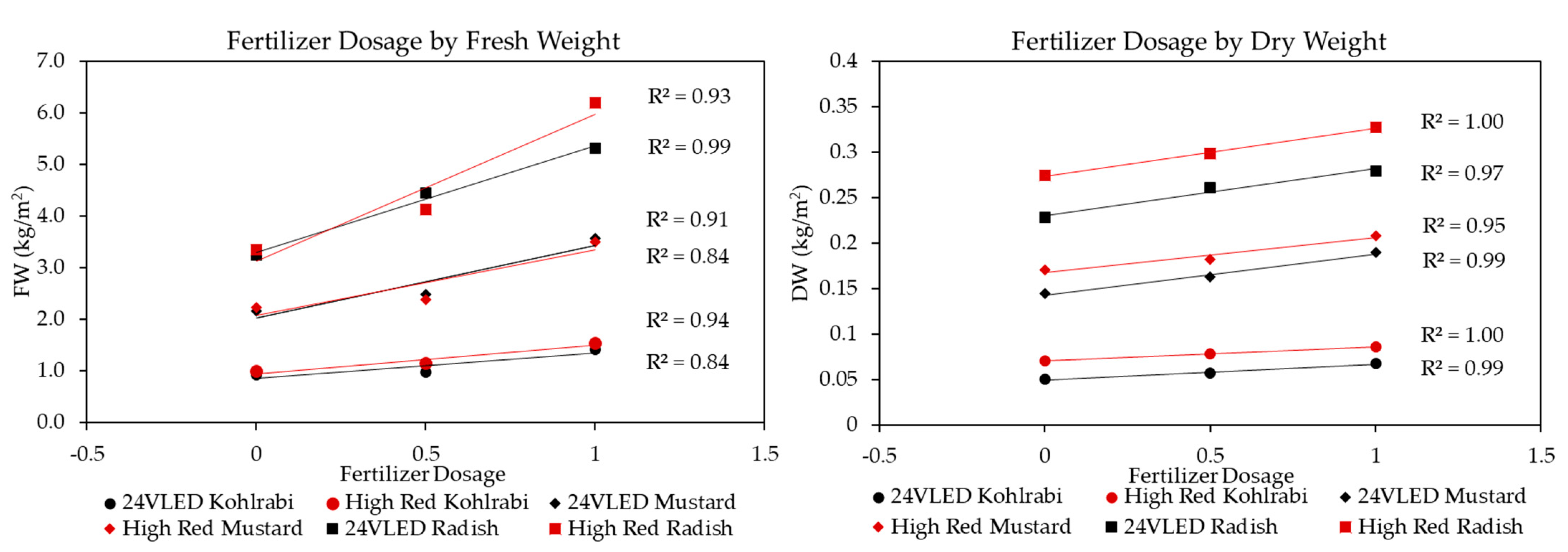
3.1. Microgreen Biomass Production and Morphology
3.2. Resource Use Efficiencies
4. Discussion
4.1. Effects of the Light Spectrum on Microgreens
4.2. Microgreen Fertilizer and Seeding Density
4.3. Microgreen Resource Use Efficiency
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Acronym | Full Statement |
| HFR | High Far-Red |
| NG | No Green |
| 24VHELED | 24-volt high efficiency LED |
| HBG | High Blue Green |
| HR | High Red |
| DLI | Daily Light Integral |
| TPFD | Total Photon Flux Density |
| Fert. | Fertilizer |
| PFD | Photon Flux Density |
| DW | Dry Weight |
| FW | Fresh Weight |
| SD | Seeding Density |
| SE | Standard Error |
| WUE | Water Use Efficiency |
| CUE | Cost Use Efficiency |
| EUE | Energy Use Efficiency |
| SUE | Land Surface Use Efficiency |
| LUE | Light Use Efficiency |
| PPE | Phytochrome Photo-Equilibrium |
References
- FAO. FAO Emissions Due to Agriculture: Global, Regional and Country Trends; FAO: Rome, Italy, 2020. [Google Scholar]
- Patle, G.T.; Kumar, M.; Khanna, M. Climate-Smart Water Technologies for Sustainable Agriculture: A Review. J. Water Clim. Chang. 2020, 11, 1455–1466. [Google Scholar] [CrossRef]
- Lal, R. Saving Global Land Resources by Enhancing Eco-Efficiency of Agroecosystems. J. Soil. Water Conserv. 2018, 73, 100A–106A. [Google Scholar] [CrossRef]
- Cordell, D.; Drangert, J.O.; White, S. The Story of Phosphorus: Global Food Security and Food for Thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Potassium: A Neglected Nutrient in Global Change. Glob. Ecol. Biogeogr. 2015, 24, 261–275. [Google Scholar] [CrossRef]
- Bolduc, A.R. The Use of Mycorrhizae to Enhance Phosphorus Uptake: A Way out the Phosphorus Crisis. J. Biofertil. Biopestic. 2011, 2, 1–5. [Google Scholar] [CrossRef]
- Gregory, P.J.; Ingram, J.S.I.; Brklacich, M. Climate Change and Food Security. Philos. Trans. R. Soc. B Biol. Sci. R. Soc 2005, 360, 2139–2148. [Google Scholar] [CrossRef]
- Vermeulen, S.J.; Campbell, B.M.; Ingram, J.S.I. Climate Change and Food Systems. Annu. Rev. Environ. Resour. 2012, 37, 195–222. [Google Scholar] [CrossRef]
- Brown, M.E.; Antle, J.M.; Backlund, P.; Carr, E.R.; Easterling, W.E.; Walsh, M.K.; Ammann, C.; Attavanich, W.; Barrett, C.B.; Bellemare, M.F.; et al. Climate Change, Global Food Security, and the U.S. Food System; United States Department of Agriculture: Washington, DC, USA, 2015. [Google Scholar]
- IPCC Climate Change 2022: Impacts, Adaptation and Vulnerability; Pörtner, H.-O.; Roberts, D.C.; Tignor, M.; Poloczanska, E.S.; Mintenbeck, K.; Alegría, A.; Craig, M.; Langsdorf, S.; Löschke, S.; Möller, V.; et al. (Eds.) Cambridge University Press: Cambridge, UK, 2022. [Google Scholar]
- Altieri, M.A.; Nicholls, C.I.; Henao, A.; Lana, M.A. Agroecology and the Design of Climate Change-Resilient Farming Systems. Agron. Sustain. Dev. 2015, 35, 869–890. [Google Scholar] [CrossRef]
- Sun, J.; Xiao, Z.; Lin, L.; Lester, G.E.; Wang, Q.; Harnly, J.M.; Chen, P. Profiling Polyphenols in Five Brassica Species Microgreens by UHPLC-PDA-ESI/HRMSn. J. Agric. Food Chem. 2013, 61, 10960–10970. [Google Scholar] [CrossRef]
- Kyriacou, M.C.; Rouphael, Y.; Di Gioia, F.; Kyratzis, A.; Serio, F.; Renna, M.; De Pascale, S.; Santamaria, P. Micro-Scale Vegetable Production and the Rise of Microgreens. Trends Food Sci. Technol. 2016, 57, 103–115. [Google Scholar] [CrossRef]
- Ying, Q.; Kong, Y.; Zheng, Y. Growth and Appearance Quality of Four Microgreen Species under Lightemitting Diode Lights with Different Spectral Combinations. HortScience 2020, 55, 1399–1405. [Google Scholar] [CrossRef]
- Zhang, X.; Bian, Z.; Li, S.; Chen, X.; Lu, C. Comparative Analysis of Phenolic Compound Profiles, Antioxidant Capacities, and Expressions of Phenolic Biosynthesis-Related Genes in Soybean Microgreens Grown under Different Light Spectra. J. Agric. Food Chem. 2019, 67, 13577–13588. [Google Scholar] [CrossRef] [PubMed]
- Samuolienė, G.; Brazaitytė, A.; Viršilė, A.; Miliauskienė, J.; Vaštakaitė-Kairienė, V.; Duchovskis, P. Nutrient Levels in Brassicaceae Microgreens Increase under Tailored Light-Emitting Diode Spectra. Front. Plant Sci. 2019, 10, 1475. [Google Scholar] [CrossRef] [PubMed]
- Ying, Q.; Kong, Y.; Jones-Baumgardt, C.; Zheng, Y. Responses of Yield and Appearance Quality of Four Brassicaceae Microgreens to Varied Blue Light Proportion in Red and Blue Light-Emitting Diodes Lighting. Sci. Hortic. 2020, 259, 108857. [Google Scholar] [CrossRef]
- Treadwell, D.; Hochmuth, R.; Landrum, L.; Laughlin, W. Microgreens: A New Specialty Crop. Edis 2020, 2020, 1–3. [Google Scholar] [CrossRef]
- Tan, L.; Nuffer, H.; Feng, J.; Kwan, S.H.; Chen, H.; Tong, X.; Kong, L. Antioxidant Properties and Sensory Evaluation of Microgreens from Commercial and Local Farms. Food Sci. Hum. Wellness 2020, 9, 45–51. [Google Scholar] [CrossRef]
- Xiao, Z.; Lester, G.E.; Luo, Y.; Wang, Q. Assessment of Vitamin and Carotenoid Concentrations of Emerging Food Products: Edible Microgreens. J. Agric. Food Chem. 2012, 60, 7644–7651. [Google Scholar] [CrossRef]
- Craver, J.K.; Gerovac, J.R.; Lopez, R.G.; Kopsell, D.A. Light Intensity and Light Quality from Sole-Source Light-Emitting Diodes Impact Phytochemical Concentrations within Brassica Microgreens. J. Am. Soc. Hortic. Sci. 2017, 142, 3–12. [Google Scholar] [CrossRef]
- Vaštakaitė-Kairienė, V.; Brazaitytė, A.; Viršilė, A.; Samuolienė, G.; Miliauskienė, J.; Jankauskienė, J.; Duchovskis, P. Pulsed Light-Emitting Diodes for Higher Contents of Mineral Elements in Mustard Microgreens. Acta Hortic. 2020, 1271, 149–154. [Google Scholar] [CrossRef]
- Zhang, Y.; Xiao, Z.; Ager, E.; Kong, L.; Tan, L. Nutritional Quality and Health Benefits of Microgreens, a Crop of Modern Agriculture. J. Future Foods 2021, 1, 58–66. [Google Scholar] [CrossRef]
- Rajan, P.; Lada, R.R.; MacDonald, M.T. Advancement in Indoor Vertical Farming for Microgreen Production. Am. J. Plant Sci. 2019, 10, 1397–1408. [Google Scholar] [CrossRef]
- EFSA Panel on Contaminants in the Food Chain (CONTAM). Statement on Possible Public Health Risks for Infants and Young Children from the Presence of Nitrates in Leafy Vegetables; Wiley-Blackwell Publishing Ltd.: Hoboken, NJ, USA, 2010; Volume 8. [Google Scholar]
- Singh, N.; Rani, S.; Mishra, A. Cruciferous Microgreens: Growing Performance and Their Scope as Super Foods at High Altitude Locations. Progress. Hortic. 2019, 51, 41. [Google Scholar] [CrossRef]
- Treftz, C.; Omaye, S.T. Hydroponics: Potential for Augmenting Sustainable Food Production in Non-Arable Regions. Nutr. Food Sci. 2016, 46, 672–684. [Google Scholar] [CrossRef]
- Orsini, F.; Pennisi, G.; Zulfiqar, F.; Gianquinto, G. Sustainable Use of Resources in Plant Factories with Artificial Lighting (PFALs). Eur. J. Hortic. Sci. 2020, 85, 297–309. [Google Scholar] [CrossRef]
- Whitelam, G.C.; Halliday, K.J. Light and Plant Development; Blackwell Pub: New York City, NY, USA, 2007; ISBN 9781405145381. [Google Scholar]
- Zhen, S.; Kusuma, P.; Bugbee, B. Toward an Optimal Spectrum for Photosynthesis and Plant Morphology in LED-Based Crop Cultivation. In Plant Factory Basics, Applications and Advances; Elsevier: Amsterdam, The Netherlands, 2022; pp. 309–327. [Google Scholar]
- Yang, F.; Liu, Q.; Cheng, Y.; Feng, L.; Wu, X.; Fan, Y.; Raza, M.A.; Wang, X.; Yong, T.; Liu, W.; et al. Low Red/Far-Red Ratio as a Signal Promotes Carbon Assimilation of Soybean Seedlings by Increasing the Photosynthetic Capacity. BMC Plant Biol. 2020, 20, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ballaré, C.L.; Pierik, R. The Shade-Avoidance Syndrome: Multiple Signals and Ecological Consequences. Plant Cell Environ. 2017, 40, 2530–2543. [Google Scholar] [CrossRef] [PubMed]
- Folta, K.M.; Childers, K.S. Light as a Growth Regulator: Controlling Plant Biology with Narrow-Bandwidth Solid-State Lighting Systems. HortScience 2008, 43, 1957–1964. [Google Scholar] [CrossRef]
- McCree, K.J. The Action Spectrum, Absorptance and Quantum Yield of Photosynthesis in Crop Plants. Agric. Meteorol. 1971, 9, 191–216. [Google Scholar] [CrossRef]
- Evans, J. The Dependence of Quantum Yield on Wavelength and Growth Irradiance. Funct. Plant Biol. 1987, 14, 69. [Google Scholar] [CrossRef]
- Inada, K. Action Spectra for Photosynthesis in Higher Plants. Plant Cell Physiol. 1976, 17, 355–365. [Google Scholar] [CrossRef]
- Meng, Q.; Kelly, N.; Runkle, E.S. Substituting Green or Far-Red Radiation for Blue Radiation Induces Shade Avoidance and Promotes Growth in Lettuce and Kale. Environ. Exp. Bot. 2019, 162, 383–391. [Google Scholar] [CrossRef]
- Emerson, R.; Chalmers, R.; Cederstrand, C. Some Factors Influencing the Long-Wave Limit of Photosynthesis. Proc. Natl. Acad. Sci. USA 1957, 40, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Emerson, R. The Quantum Yield of Photosynthesis. Annu. Rev. Plant Physiol. 1958, 9, 1–24. [Google Scholar] [CrossRef]
- Liu, H.; Fu, Y.; Yu, J.; Liu, H. Accumulation and Primary Metabolism of Nitrate in Lettuce (Lactuca sativa L. Var. Youmaicai) Grown Under Three Different Light Sources. Commun. Soil. Sci. Plant Anal. 2016, 47, 1994–2002. [Google Scholar] [CrossRef]
- Saengtharatip, S.; Goto, N.; Kozai, T.; Yamori, W. Green Light Penetrates inside Crisp Head Lettuce Leading to Chlorophyll and Ascorbic Acid Content Enhancement. Acta Hortic. 2020, 1273, 261–269. [Google Scholar] [CrossRef]
- Liu, J.; van Iersel, M.W. Photosynthetic Physiology of Blue, Green, and Red Light: Light Intensity Effects and Underlying Mechanisms. Front. Plant Sci. 2021, 12, 328. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, S.D.; Folta, K.M. Green Light Control of Anthocyanin Production in Microgreens. Acta Hortic. 2016, 1134, 13–18. [Google Scholar] [CrossRef]
- Snowden, M.C.; Cope, K.R.; Bugbee, B. Sensitivity of Seven Diverse Species to Blue and Green Light: Interactions with Photon Flux. PLoS ONE 2016, 11, 1–32. [Google Scholar] [CrossRef]
- Kamal, K.Y.; Khodaeiaminjan, M.; El-Tantawy, A.A.; Moneim, D.A.; Salam, A.A.; Ash-shormillesy, S.M.A.I.; Attia, A.; Ali, M.A.S.; Herranz, R.; El-Esawi, M.A.; et al. Evaluation of Growth and Nutritional Value of Brassica Microgreens Grown under Red, Blue and Green LEDs Combinations. Physiol. Plant 2020, 169, 625–638. [Google Scholar] [CrossRef]
- Dou, H.; Niu, G.; Gu, M. Photosynthesis, Morphology, Yield, and Phytochemical Accumulation in Basil Plants Influenced by Substituting Green Light for Partial Red and/or Blue Light. HortScience 2019, 54, 1769–1776. [Google Scholar] [CrossRef]
- Orlando, M.; Trivellini, A.; Incrocci, L.; Ferrante, A.; Mensuali, A. The Inclusion of Green Light in a Red and Blue Light Background Impact the Growth and Functional Quality of Vegetable and Flower Microgreen Species. Horticulturae 2022, 8, 217. [Google Scholar] [CrossRef]
- Zhen, S.; Bugbee, B. Far-Red Photons Have Equivalent Efficiency to Traditional Photosynthetic Photons: Implications for Redefining Photosynthetically Active Radiation. Plant Cell Environ. 2020, 43, 1259–1272. [Google Scholar] [CrossRef] [PubMed]
- Zhen, S.; van Iersel, M.W. Far-Red Light Is Needed for Efficient Photochemistry and Photosynthesis. J. Plant Physiol. 2016, 209, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Legendre, R.; van Iersel, M.W. Supplemental Far-Red Light Stimulates Lettuce Growth: Disentangling Morphological and Physiological Effects. Plants 2021, 10, 166. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Kusuma, P.; Carvalho, S.D.; Li, Y.; Folta, K.M. Manipulation of Seedling Traits with Pulsed Light in Closed Controlled Environments. Environ. Exp. Bot. 2019, 166, 103–108. [Google Scholar] [CrossRef]
- Kozai, T.; Sakaguchi, S.; Akiyama, T.; Yamada, K.; Ohshima, K. Design and Management of PFAL. In Plant Factory: An Indoor Vertical Farming System for Efficient Quality Food Production; Elsevier Inc.: Amsterdam, The Netherlands, 2015; pp. 295–312. ISBN 9780128017753. [Google Scholar]
- Kalantari, F.; Tahir, O.M.; Joni, R.A.; Fatemi, E. Opportunities and Challenges in Sustainability of Vertical Farming: A Review. J. Landsc. Ecol. 2018, 11, 35–60. [Google Scholar] [CrossRef]
- Yokoyama, R. Energy Consumption and Heat Sources in Plant Factories. In Plant Factory Using Artificial Light: Adapting to Environmental Disruption and Clues to Agricultural Innovation; Elsevier: Amsterdam, The Netherlands, 2018; pp. 177–184. ISBN 9780128139745. [Google Scholar]
- Kozai, T. Role and Characteristics of PFALs. In Plant Factory Basics, Applications and Advances; Elsevier: Amsterdam, The Netherlands, 2022; pp. 25–55. ISBN 9780323851527. [Google Scholar]
- Xu, Y. Nature and Source of Light for Plant Factory. In Plant Factory Using Artificial Light: Adapting to Environmental Disruption and Clues to Agricultural Innovation; Elsevier: Amsterdam, The Netherlands, 2018; pp. 47–69. ISBN 9780128139745. [Google Scholar]
- Zhen, S.; van Iersel, M.; Bugbee, B. Why Far-Red Photons Should Be Included in the Definition of Photosynthetic Photons and the Measurement of Horticultural Fixture Efficacy. Front. Plant Sci. 2021, 12, 1158. [Google Scholar] [CrossRef]
- Sager, J.C.; Smith, W.; Edwards, J.L.; Cyr, K.L. Photosynthetic Efficiency and Phytochrome Photoequilibria Determination Using Spectral Data. Trans. ASAE 1988, 31, 1882–1889. [Google Scholar] [CrossRef]
- Puccinelli, M.; Maggini, R.; Angelini, L.G.; Santin, M.; Landi, M.; Tavarini, S.; Castagna, A.; Incrocci, L. Can Light Spectrum Composition Increase Growth and Nutritional Quality of Linum usitatissimum L. Sprouts and Microgreens? Horticulturae 2022, 8, 98. [Google Scholar] [CrossRef]
- Runkle, E.S.; Meng, Q.; Park, Y. LED Applications in Greenhouse and Indoor Production of Horticultural Crops. Proc. Acta Hortic. Int. Soc. Hortic. Sci. 2019, 1263, 17–29. [Google Scholar] [CrossRef]
- R Core Team. R A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Lenth, R. Emmeans: Estimated Marginal Means, Aka Least-Squares Means; R Package: Vienna, Austria, 2023. [Google Scholar]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix; R Package: Vienna, Austria, 2021. [Google Scholar]
- Park, Y.; Runkle, E.S. Far-Red Radiation Promotes Growth of Seedlings by Increasing Leaf Expansion and Whole-Plant Net Assimilation. Environ. Exp. Bot. 2017, 136, 41–49. [Google Scholar] [CrossRef]
- Lee, M.J.; Son, K.H.; Oh, M.M. Increase in Biomass and Bioactive Compounds in Lettuce under Various Ratios of Red to Far-Red LED Light Supplemented with Blue LED Light. Hortic. Environ. Biotechnol. 2016, 57, 139–147. [Google Scholar] [CrossRef]
- Tan, T.; Li, S.; Fan, Y.; Wang, Z.; Ali Raza, M.; Shafiq, I.; Wang, B.; Wu, X.; Yong, T.; Wang, X.; et al. Far-Red Light: A Regulator of Plant Morphology and Photosynthetic Capacity. Crop J. 2022, 10, 300–309. [Google Scholar] [CrossRef]
- Zhen, S.; Haidekker, M.; van Iersel, M.W. Far-Red Light Enhances Photochemical Efficiency in a Wavelength-Dependent Manner. Physiol. Plant 2019, 167, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Kalaitzoglou, P.; van Ieperen, W.; Harbinson, J.; van der Meer, M.; Martinakos, S.; Weerheim, K.; Nicole, C.C.S.; Marcelis, L.F.M. Effects of Continuous or End-of-Day Far-Red Light on Tomato Plant Growth, Morphology, Light Absorption, and Fruit Production. Front. Plant Sci. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Tao, Y.; Lim, J.; Shaw, A.; Chory, J. Regulation of Phytochrome B Nuclear Localization through Light-Dependent Unmasking of Nuclear-Localization Signals. Curr. Biol. 2005, 15, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Hooks, T.; Sun, L.; Kong, Y.; Masabni, J.; Niu, G. Adding UVA and Far-Red Light to White LED Affects Growth, Morphology, and Phytochemicals of Indoor-Grown Microgreens. Sustainability 2022, 14, 8552. [Google Scholar] [CrossRef]
- Ying, Q.; Kong, Y.; Zheng, Y. Applying Blue Light Alone, or in Combination with Far-Red Light, during Nighttime Increases Elongation without Compromising Yield and Quality of Indoor-Grown Microgreens. HortScience 2020, 55, 876–881. [Google Scholar] [CrossRef]
- Kong, Y.; Schiestel, K.; Zheng, Y. Pure Blue Light Effects on Growth and Morphology Are Slightly Changed by Adding Low-Level UVA or Far-Red Light: A Comparison with Red Light in Four Microgreen Species. Environ. Exp. Bot. 2019, 157, 58–68. [Google Scholar] [CrossRef]
- Kong, Y.; Schiestel, K.; Zheng, Y. Maximum Elongation Growth Promoted as a Shade-Avoidance Response by Blue Light Is Related to Deactivated Phytochrome: A Comparison with Red Light in Four Microgreen Species. Can. J. Plant Sci. 2020, 100, 314–326. [Google Scholar] [CrossRef]
- Hogewoning, S.W.; Wientjes, E.; Douwstra, P.; Trouwborst, G.; van Ieperen, W.; Croce, R.; Harbinson, J. Photosynthetic Quantum Yield Dynamics: From Photosystems to Leaves. Plant Cell 2012, 24, 1921–1935. [Google Scholar] [CrossRef] [PubMed]
- Wientjes, E.; Philippi, J.; Borst, J.W.; van Amerongen, H. Imaging the Photosystem I/Photosystem II Chlorophyll Ratio inside the Leaf. Biochim. Et Biophys. Acta (BBA)—Bioenerg. 2017, 1858, 259–265. [Google Scholar] [CrossRef] [PubMed]
- van Amerongen, H.; Wientjes, E. Harvesting Light. In Photosynthesis in Action: Harvesting Light, Generating Electrons, Fixing Carbon; Elsevier: Amsterdam, The Netherlands, 2022; pp. 3–16. ISBN 9780128237823. [Google Scholar]
- Gerovac, J.R.; Craver, J.K.; Boldt, J.K.; Lopez, R.G. Light Intensity and Quality from Sole-Source Light-Emitting Diodes Impact Growth, Morphology, and Nutrient Content of Brassica Microgreens. HortScience 2016, 51, 497–503. [Google Scholar] [CrossRef]
- Li, Y.; Wu, L.; Jiang, H.; He, R.; Song, S.; Su, W.; Liu, H. Supplementary Far-Red and Blue Lights Influence the Biomass and Phytochemical Profiles of Two Lettuce Cultivars in Plant Factory. Molecules 2021, 26, 7405. [Google Scholar] [CrossRef] [PubMed]
- Long, S.P.; Zhu, X.; Naidu, S.L.; Ort, D.R. Can Improvement in Photosynthesis Increase Crop Yields? Plant Cell Environ. 2006, 29, 315–330. [Google Scholar] [CrossRef] [PubMed]
- Ort, D.R.; Merchant, S.S.; Alric, J.; Barkan, A.; Blankenship, R.E.; Bock, R.; Croce, R.; Hanson, M.R.; Hibberd, J.M.; Long, S.P.; et al. Redesigning Photosynthesis to Sustainably Meet Global Food and Bioenergy Demand. Proc. Natl. Acad. Sci. USA 2015, 112, 8529–8536. [Google Scholar] [CrossRef]
- Ying, Q.; Kong, Y.; Zheng, Y. Overnight Supplemental Blue, Rather than Far-Red, Light Improves Microgreen Yield and Appearance Quality without Compromising Nutritional Quality during Winter Greenhouse Production. HortScience 2020, 55, 1468–1474. [Google Scholar] [CrossRef]
- Lobiuc, A.; Vasilache, V.; Pintilie, O.; Stoleru, T.; Burducea, M.; Oroian, M.; Zamfirache, M.M. Blue and Red LED Illumination Improves Growth and Bioactive Compounds Contents in Acyanic and Cyanic Ocimum basilicum L. Microgreens. Molecules 2017, 22, 2111. [Google Scholar] [CrossRef]
- Bantis, F. Light Spectrum Differentially Affects the Yield and Phytochemical Content of Microgreen Vegetables in a Plant Factory. Plants 2021, 10, 2182. [Google Scholar] [CrossRef]
- Brazaitytė, A.; Miliauskienė, J.; Vaštakaitė-Kairienė, V.; Sutulienė, R.; Laužikė, K.; Duchovskis, P.; Małek, S. Effect of Different Ratios of Blue and Red Led Light on Brassicaceae Microgreens under a Controlled Environment. Plants 2021, 10, 801. [Google Scholar] [CrossRef]
- Clavijo-Herrera, J.; Van Santen, E.; Gómez, C. Growth, Water-Use Efficiency, Stomatal Conductance, and Nitrogen Uptake of Two Lettuce Cultivars Grown under Different Percentages of Blue and Red Light. Horticulturae 2018, 4, 16. [Google Scholar] [CrossRef]
- Pennisi, G.; Pistillo, A.; Orsini, F.; Gianquinto, G.; Fernandez, J.A.; Crepaldi, A.; Nicola, S. Improved Red and Blue Ratio in LED Lighting for Indoor Cultivation of Basil. In Proceedings of the Acta Horticulturae, International Society for Horticultural Science, Istanbul, Turkey, 20 March 2020; Volume 1271, pp. 115–118. [Google Scholar]
- Pistillo, A.; Pennisi, G.; Crepaldi, A.; Giorgioni, M.E.; Minelli, A.; Orsini, F.; Gianquinto, G. Influence of Red:Blue Ratio in LED Lighting for Indoor Cultivation of Edible Marigold Flowers. In Proceedings of the Acta Horticulturae, International Society for Horticultural Science, Malmö, Sweden, 1 April 2022; Volume 1337, pp. 249–254. [Google Scholar]
- Pennisi, G.; Orsini, F.; Blasioli, S.; Cellini, A.; Crepaldi, A.; Braschi, I.; Spinelli, F.; Nicola, S.; Fernandez, J.A.; Stanghellini, C.; et al. Resource Use Efficiency of Indoor Lettuce (Lactuca sativa L.) Cultivation as Affected by Red:Blue Ratio Provided by LED Lighting. Sci. Rep. 2019, 9, 14127. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, G.; Blasioli, S.; Cellini, A.; Maia, L.; Crepaldi, A.; Braschi, I.; Spinelli, F.; Nicola, S.; Fernandez, J.A.; Stanghellini, C.; et al. Unraveling the Role of Red:Blue LED Lights on Resource Use Efficiency and Nutritional Properties of Indoor Grown Sweet Basil. Front. Plant Sci. 2019, 10, 305. [Google Scholar] [CrossRef] [PubMed]
- Loconsole, D.; Cocetta, G.; Santoro, P.; Ferrante, A. Optimization of LED Lighting and Quality Evaluation of Romaine Lettuce Grown in an Innovative Indoor Cultivation System. Sustainability 2019, 11, 841. [Google Scholar] [CrossRef]
- Kono, M.; Kawaguchi, H.; Mizusawa, N.; Yamori, W.; Suzuki, Y.; Terashima, I. Far-Red Light Accelerates Photosynthesis in the Low-Light Phases of Fluctuating Light. Plant Cell Physiol. 2020, 61, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Matsuda, R.; Fujiwara, K. A Mathematical Model of Photosynthetic Electron Transport in Response to the Light Spectrum Based on Excitation Energy Distributed to Photosystems. Plant Cell Physiol. 2018, 59, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- Battle, M.W.; Jones, M.A. Cryptochromes Integrate Green Light Signals into the Circadian System. Plant Cell Environ. 2020, 43, 16–27. [Google Scholar] [CrossRef]
- Smith, H.L.; Mcausland, L.; Murchie, E.H. Don’t Ignore the Green Light: Exploring Diverse Roles in Plant Processes. J. Exp. Bot. 2017, 68, 2099–2110. [Google Scholar] [CrossRef]
- Bian, Z.; Cheng, R.; Wang, Y.; Yang, Q.; Lu, C. Effect of Green Light on Nitrate Reduction and Edible Quality of Hydroponically Grown Lettuce (Lactuca sativa L.) under Short-Term Continuous Light from Red and Blue Light-Emitting Diodes. Environ. Exp. Bot. 2018, 153, 63–71. [Google Scholar] [CrossRef]
- Sellaro, R.; Crepy, M.; Trupkin, S.A.; Karayekov, E.; Buchovsky, A.S.; Rossi, C.; Casal, J.J. Cryptochrome as a Sensor of the Blue/Green Ratio of Natural Radiation in Arabidopsis. Plant Physiol. 2010, 154, 401–409. [Google Scholar] [CrossRef]
- Kong, Y.; Zheng, Y. Growth and Morphology Responses to Narrow-Band Blue Light and Its Co-Action with Low-Level UVB or Green Light: A Comparison with Red Light in Four Microgreen Species. Environ. Exp. Bot. 2020, 178, 104189. [Google Scholar] [CrossRef]
- Johkan, M.; Shoji, K.; Goto, F.; Hahida, S.; Yoshihara, T. Effect of Green Light Wavelength and Intensity on Photomorphogenesis and Photosynthesis in Lactuca Sativa. Environ. Exp. Bot. 2012, 75, 128–133. [Google Scholar] [CrossRef]
- El-Nakhel, C.; Pannico, A.; Graziani, G.; Kyriacou, M.C.; Gaspari, A.; Ritieni, A.; de Pascale, S.; Rouphael, Y. Nutrient Supplementation Configures the Bioactive Profile and Production Characteristics of Three Brassica L. Microgreens Species Grown in Peat-Based Media. Agronomy 2021, 11, 346. [Google Scholar] [CrossRef]
- Barbi, S.; Barbieri, F.; Bertacchini, A.; Barbieri, L.; Montorsi, M. Effects of Different LED Light Recipes and NPK Fertilizers on Basil Cultivation for Automated and Integrated Horticulture Methods. Appl. Sci. 2021, 11, 2497. [Google Scholar] [CrossRef]
- Wieth, A.R.; Pinheiro, W.D.; Duarte, T.D.S. Purple Cabbage Microgreens Grown in Different Substrates and Nutritive Solution Concentrations. Rev. Caatinga 2019, 32, 976–985. [Google Scholar] [CrossRef]
- Mi, R.; Taylor, A.G.; Smart, L.B.; Mattson, N.S. Developing Production Guidelines for Baby Leaf Hemp (Cannabis sativa L.) as an Edible Salad Green: Cultivar, Sowing Density and Seed Size. Agriculture 2020, 10, 617. [Google Scholar] [CrossRef]
- Wang, Q.; Kniel, K.E. Survival and Transfer of Murine Norovirus within a Hydroponic System during Kale and Mustard Microgreen Harvesting. Appl. Environ. Microbiol. 2016, 82, 705–713. [Google Scholar] [CrossRef]
- Xiao, Z.; Bauchan, G.; Nichols-Russell, L.; Luo, Y.; Wang, Q.; Nou, X. Proliferation of Escherichia Coli O157:H7 in Soil-Substitute and Hydroponic Microgreen Production Systems. J. Food Prot. 2015, 78, 1785–1790. [Google Scholar] [CrossRef]
- Priti; Sangwan, S.; Kukreja, B.; Mishra, G.P.; Dikshit, H.K.; Singh, A.; Aski, M.; Kumar, A.; Taak, Y.; Stobdan, T.; et al. Yield Optimization, Microbial Load Analysis, and Sensory Evaluation of Mungbean (Vigna radiata L.), Lentil (Lens culinaris Subsp. Culinaris), and Indian Mustard (Brassica juncea L.) Microgreens Grown under Greenhouse Conditions. PLoS ONE 2022, 17, e0268085. [Google Scholar] [CrossRef]
- Pennisi, G.; Sanyé-Mengual, E.; Orsini, F.; Crepaldi, A.; Nicola, S.; Ochoa, J.; Fernandez, J.A.; Gianquinto, G. Modelling Environmental Burdens of Indoor-Grown Vegetables and Herbs as Affected by Red and Blue LED Lighting. Sustainability 2019, 11, 4063. [Google Scholar] [CrossRef]
- Rahman, M.M.; Vasiliev, M.; Alameh, K. Led Illumination Spectrum Manipulation for Increasing the Yield of Sweet Basil (Ocimum basilicum L.). Plants 2021, 10, 344. [Google Scholar] [CrossRef] [PubMed]
- Tavan, M.; Wee, B.; Brodie, G.; Fuentes, S.; Pang, A.; Gupta, D. Optimizing Sensor-Based Irrigation Management in a Soilless Vertical Farm for Growing Microgreens. Front. Sustain. Food Syst. 2021, 4, 622720. [Google Scholar] [CrossRef]
- Weber, C.F. Broccoli Microgreens: A Mineral-Rich Crop That Can Diversify Food Systems. Front. Nutr. 2017, 4, 7. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Valoya Reference | High Red | No Green | High Blue Green | High Far-Red | 24VHELED | |
|---|---|---|---|---|---|---|
| Blue µmol/m2/s (400–500 nm) | x | 28.64 | 95.38 | 70.04 | 53.43 | 7.91 |
| Green µmol/m2/s (500–600 nm) | x | 50.84 | 0.44 | 72.81 | 60.30 | 21.41 |
| Red µmol/m2/s (600–700 nm) | x | 165.50 | 156.60 | 106.75 | 93.30 | 15.68 |
| Far-Red µmol/m2/s (700–800 nm) | x | 21.93 | 22.06 | 21.88 | 61.96 | 1.10 |
| PPFD µmol/m2/s (400–700 nm) | x | 244.98 | 252.42 | 249.60 | 207.03 | 45.00 |
| PFD µmol/m2/s (350–800 nm) | x | 266.92 | 274.49 | 271.48 | 268.99 | 46.09 |
| YPFD µmol/m2/s (350–800 nm) | x | 219.90 | 222.34 | 212.36 | 186.07 | 39.75 |
| DLI (mol/m2) | x | 15.37 | 15.81 | 15.64 | 15.49 | 2.65 |
| % Blue (400–500 nm) | 10% | 10.73% | 34.75% | 25.80% | 19.86% | 17.16% |
| % Green (500–600 nm) | 19% | 19.05% | 0.16% | 26.82% | 22.42% | 46.45% |
| % Red (600–700 nm) | 63% | 62.01% | 57.05% | 39.32% | 34.69% | 34.02% |
| % Far-Red (700–800 nm) | 8% | 8.22% | 8.04% | 8.06% | 23.03% | 2.38% |
| B:R | 0.16 | 0.17 | 0.61 | 0.66 | 0.57 | 0.50 |
| B:G | 0.53 | 0.56 | 216.77 | 0.96 | 0.89 | 0.37 |
| G:R | 0.30 | 0.31 | 0.00 | 0.68 | 0.65 | 1.37 |
| R:FR | 7.88 | 7.55 | 7.10 | 4.88 | 1.51 | 14.25 |
| FR:PAR | 0.09 | 0.09 | 0.09 | 0.09 | 0.30 | 0.02 |
| PPE | x | 0.79 | 0.84 | 0.83 | 0.75 | 0.86 |
| CIE | 60.00 | 22.01 | 0.00 | 60.78 | 60.04 | 81.29 |
| Power Consumption (Watts) | x | 171 | 141 | 192 | 189 | 48 |
| Period | Low Zone Temp. (°C) | Low Zone RH (%) | Low Zone VPD (kPa) | High Zone Temp. (°C) | High Zone RH (%) | High Zone VPD (kPa) | CC Temp (°C) |
|---|---|---|---|---|---|---|---|
| One | 21.6 ± 0.2 | 69.6 ± 4.5 | 0.79 ± 0.12 | 21.8 ± 0.2 | 68.8 ± 3.6 | 0.81 ± 0.10 | 21.1 ± 0.1 |
| Two | 21.5 ± 0.2 | ND | ND | 21.9 ± 0.6 | 66.1 ± 5.1 | 0.89 ± 0.15 | 21.0 ± 0.1 |
| Measurement | Period One Unfert. | Period Two Unfert. | Period One Fert. | Period Two Fert. |
|---|---|---|---|---|
| pH | 6.57 ± 0.55 | 6.56 ± 0.28 | 6.20 ± 0.20 | 6.18 ± 0.19 |
| EC | 0.81 ± 0.04 | 0.86 ± 0.06 | 1.86 ± 0.09 | 1.81 ± 0.08 |
| Source of Variation | df | FW (η2) | FW (p) | DW (η2) | DW (p) |
|---|---|---|---|---|---|
| Species | 2 | 65.28% | <0.001 | 77.24% | <0.001 |
| Light Recipe | 4 | 2.09% | <0.001 | 2.32% | <0.001 |
| Fertilizer | 1 | 15.65% | <0.001 | 3.78% | <0.001 |
| Seeding Density | 1 | 6.81% | <0.001 | 10.28% | <0.001 |
| Light Recipe:Fertilizer | 4 | 0.02% | 0.98 | 0.09% | 0.42 |
| Fertilizer:Seeding Density | 1 | 0.21% | 0.02 | 0.04% | 0.17 |
| Light Recipe:Seeding Density | 4 | 0.06% | 0.83 | 0.10% | 0.39 |
| Light Recipe:Fertilizer:Seeding Density | 4 | 0.01% | 1.00 | 0.03% | 0.83 |
| Error | 9.88% | 6.11% |
| Kohlrabi (Unfert.) | Kohlrabi (Fert.) | Mustard (Unfert.) | Mustard (Fert.) | Radish (Unfert.) | Radish (Fert.) | |
|---|---|---|---|---|---|---|
| 24VHELED | 511.61 ± 10.66 | 814.45 ± 25.5 | 336.96 ± 50.04 | 601.42 ± 68.57 | 440.39 ± 131.75 | 512.53 ± 23.53 |
| Low Far-Red OSRAM | 400.25 ± 34.19 | 583.03 ± 21.00 | 309.73 ± 17.64 | 408.33 ± 84.12 | 282.98 ± 8.13 | 398.82 ± 15.83 |
| High Far-Red OSRAM | 557.73 ± 20.47 | 559.67 ± 14.57 | 415.7 ± 23.56 | 473.77 ± 7.96 | 352.33 ± 13.07 | 465.73 ± 9.97 |
| Species | Fertilizer | 24VHELED | High Blue and Green | High Far-Red | High Red | No Green |
|---|---|---|---|---|---|---|
| Kohlrabi | No | 5.25% | 6.09% | 6.15% | 7.14% | 7.10% |
| Kohlrabi | Yes | 4.84% | 5.29% | 5.57% | 5.64% | 6.15% |
| Mustard | No | 6.48% | 7.21% | 6.86% | 7.71% | 7.94% |
| Mustard | Yes | 5.26% | 5.46% | 5.06% | 5.95% | 5.67% |
| Radish | No | 6.52% | 7.14% | 6.66% | 7.85% | 7.71% |
| Radish | Yes | 5.17% | 5.42% | 4.57% | 5.29% | 5.45% |
| Light Recipe | Kohlrabi (FW) | Kohlrabi (DW) | Mustard (FW) | Mustard (DW) | Radish (FW) | Radish (DW) |
|---|---|---|---|---|---|---|
| 24VHELED | 1.51 | 1.35 | 1.67 | 1.32 | 1.64 | 1.22 |
| High Blue and Green | 1.57 | 1.36 | 1.75 | 1.31 | 1.53 | 1.16 |
| High Far-Red | 1.48 | 1.36 | 1.49 | 1.13 | 1.50 | 1.04 |
| High Red | 1.58 | 1.26 | 1.61 | 1.25 | 1.59 | 1.05 |
| No Green | 1.74 | 1.51 | 1.76 | 1.26 | 1.55 | 1.09 |
| Total Average | 1.58 ± 0.05 | 1.36 ± 0.04 | 1.66 ± 0.05 | 1.25 ± 0.03 | 1.56 ± 0.03 | 1.11 ± 0.02 |
| Treatment | Kohlrabi | Mustard | Radish |
|---|---|---|---|
| Unfertilized FW | 1.33 ± 0.06 | 1.45 ± 0.01 | 1.38 ± 0.04 |
| Fertilized FW | 1.42 ± 0.04 | 1.31 ± 0.04 | 1.35 ± 0.02 |
| Average | 1.37 ± 0.05 | 1.38 ± 0.02 | 1.36 ± 0.03 |
| Fert. | Light Recipe | LUE (g FW/mol PFD) | Light EUE (g FW/kWh) | Total EUE (g FW/kWh) | WUE (g FW/L H2O) | SUE (g FW/m2) | CUE (g FW/Dollar) |
|---|---|---|---|---|---|---|---|
| No | 24VHELED | 45.75 ± 3.83 c | 1116.04 ± 93.44 c | 2.27 ± 0.19 a | 57.14 ± 4.79 b,c | 263.59 ± 22.07 a | 70.84 ± 5.36 c |
| No | HBG | 8.55 ± 1.10 a | 189.00 ± 24.23 a | 2.48 ± 0.32 b | 55.25 ± 7.08 a,b | 290.30 ± 37.21 b | 54.33 ± 6.49 a |
| No | HFR | 10.67 ± 1.28 b | 237.25 ± 28.52 b | 3.06 ± 0.37 c | 63.13 ± 7.59 c | 358.73 ± 43.11 c | 67.60 ± 7.58 b |
| No | HR | 7.74 ± 0.80 a | 188.85 ± 19.62 a,b | 2.21 ± 0.23 b | 46.84 ± 4.87 a | 258.35 ± 26.84 b | 50.77 ± 4.96 a |
| No | NG | 8.01 ± 1.16 a | 243.78 ± 35.14 b | 2.35 ± 0.34 a,b | 50.57 ± 7.29 a | 274.98 ± 39.64 a,b | 56.65 ± 7.57 a |
| Yes | 24VHELED | 74.51 ± 4.59 b | 1817.46 ± 111.98 c | 3.70 ± 0.23 a | 93.05 ± 5.73 b,c | 429.26 ± 26.45 a | 114.67 ± 6.48 c |
| Yes | HBG | 15.77 ± 1.87 a | 301.93 ± 26.66 a | 3.96 ± 0.35 b | 88.26 ± 7.79 a,b | 463.77 ± 40.94 b | 86.56 ± 7.22 a |
| Yes | HFR | 15.95 ± 1.94 a | 354.62 ± 43.12 b | 4.58 ± 0.56 c | 94.36 ± 11.47 c | 536.19 ± 65.19 c | 100.47 ± 11.42 b |
| Yes | HR | 14.03 ± 1.84 a | 342.30 ± 44.87 a,b | 4.00 ± 0.53 b | 84.90 ± 11.13 a | 468.26 ± 61.38 b | 90.72 ± 11.06 a |
| Yes | NG | 14.95 ± 1.45 a | 454.71 ± 44.13 b | 4.39 ± 0.43 a,b | 94.33 ± 9.15 a | 512.91 ± 49.78 a,b | 104.46 ± 9.42 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cowden, R.J.; Markussen, B.; Ghaley, B.B.; Henriksen, C.B. The Effects of Light Spectrum and Intensity, Seeding Density, and Fertilization on Biomass, Morphology, and Resource Use Efficiency in Three Species of Brassicaceae Microgreens. Plants 2024, 13, 124. https://doi.org/10.3390/plants13010124
Cowden RJ, Markussen B, Ghaley BB, Henriksen CB. The Effects of Light Spectrum and Intensity, Seeding Density, and Fertilization on Biomass, Morphology, and Resource Use Efficiency in Three Species of Brassicaceae Microgreens. Plants. 2024; 13(1):124. https://doi.org/10.3390/plants13010124
Chicago/Turabian StyleCowden, Reed John, Bo Markussen, Bhim Bahadur Ghaley, and Christian Bugge Henriksen. 2024. "The Effects of Light Spectrum and Intensity, Seeding Density, and Fertilization on Biomass, Morphology, and Resource Use Efficiency in Three Species of Brassicaceae Microgreens" Plants 13, no. 1: 124. https://doi.org/10.3390/plants13010124