
Genomics and Transcriptomics Reveal Genetic Contribution to Population Diversity and Specific Traits in Coconut


Abstract
:1. Introduction
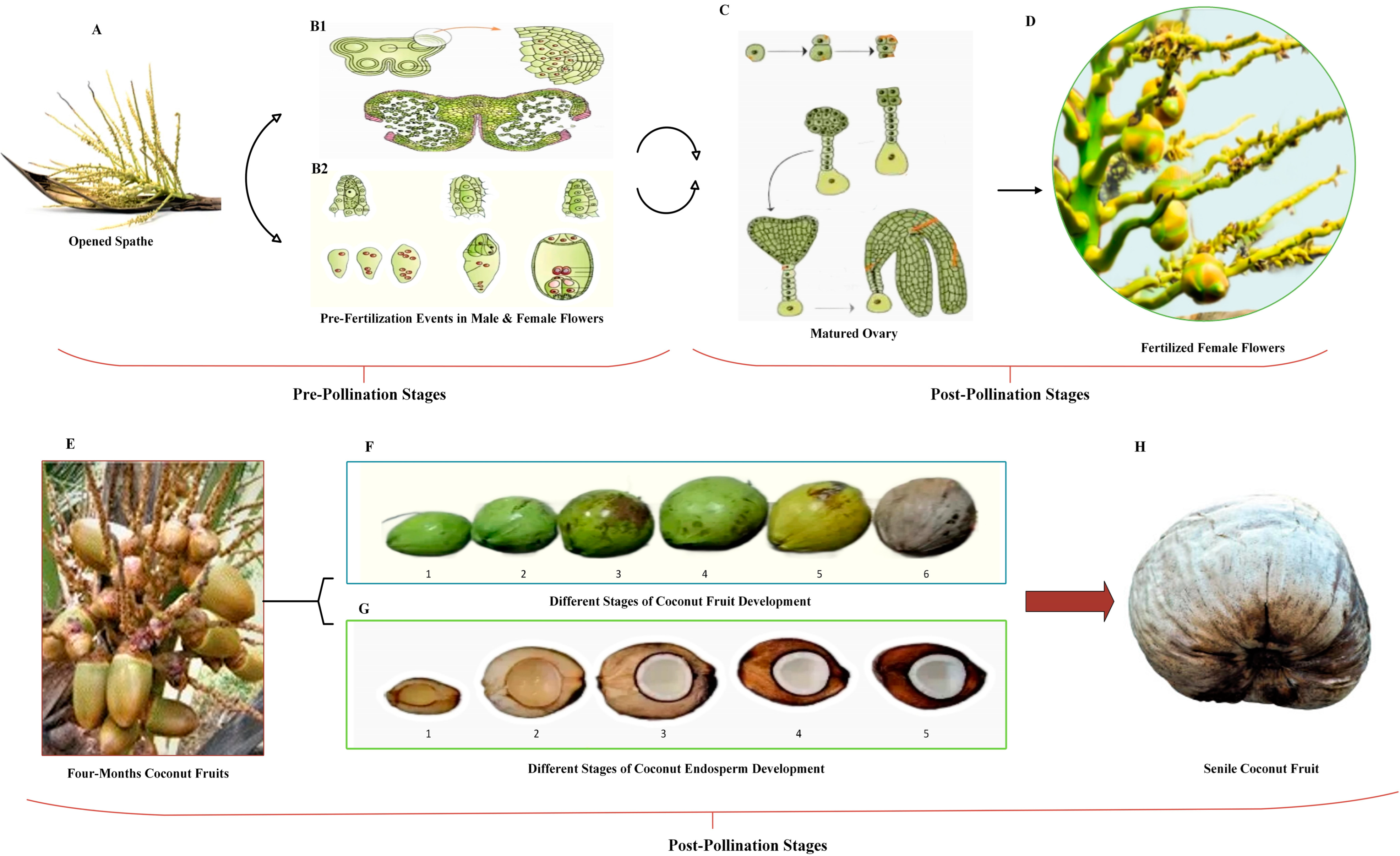
2. Fruit Development in Coconut
3. Genetic Diversity Studies Using Molecular Markers
Linkage Mapping, QTL Identification, and Association Studies
4. Whole Genome Sequencing Efforts
4.1. Discovery of Genes Involved in Oil Biosynthesis from the Whole Genome Sequence
4.2. Sequencing of Mitochondrial and Chloroplast Genomes
5. Transcriptomics Initiatives
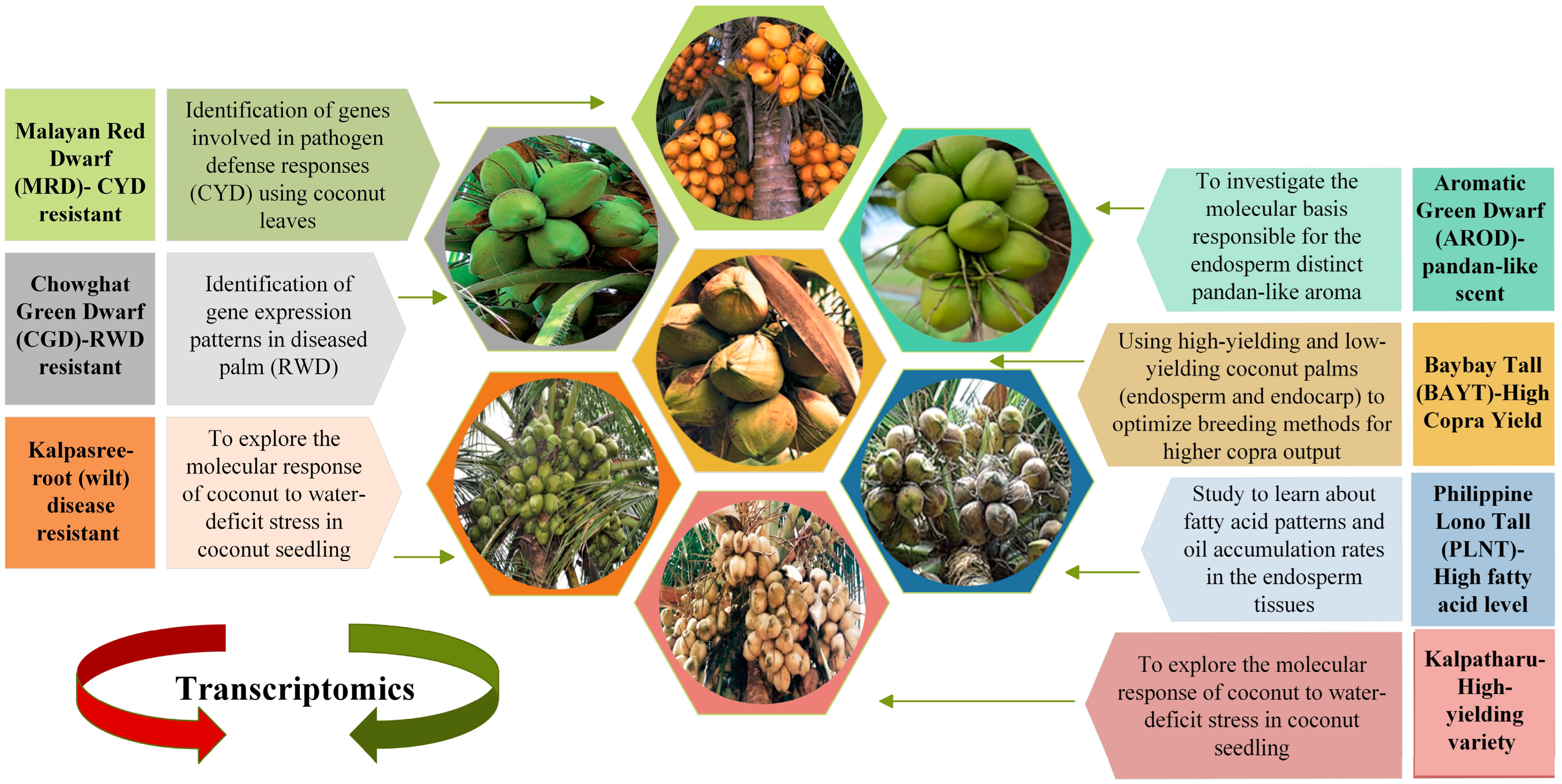
5.1. Comparative Transcriptomics between Different Genotypes

5.2. Transcriptomic to Reveal Molecular Mechanisms Underlying Biochemical and Developmental Processes in Specific Tissues
5.3. Transcriptomics for Fresh Insights on Defense Responses against Pathogen Attacks
6. Conclusions and Future Perspective
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Yang, Y.; Iqbal, A.; Qadri, R. Breeding of Coconut (Cocos nucifera L.): The Tree of Life: Volume 3. In Advances in Plant Breeding Strategies: Fruits; Springer: Cham, Switzerland, 2018; pp. 673–725. [Google Scholar] [CrossRef]
- Rajamohan, T.; Archana, U. Nutrition and Health Aspects of Coconut. In The Coconut Palm (Cocos nucifera L.)—Research and Development Perspectives; Krishnakumar, V., Thampan, P.K., Nair, M.A., Eds.; Springer: Singapore, 2018; pp. 757–777. [Google Scholar] [CrossRef]
- UN Data F-F and, AO. Available online: http://data.un.org/Data.aspx?d=FAO&f=itemCode%3A249 (accessed on 19 December 2022).
- Mat, K.; Abdul Kari, Z.; Rusli, N.D.; Che Harun, H.; Wei, L.S.; Rahman, M.M.; Khalid, H.N.M.; Hanafiah, M.H.M.A.; Sukri, S.A.M.; Khalif, R.I.A.R.; et al. Coconut Palm: Food, Feed, and Nutraceutical Properties. Animals 2022, 12, 2107. [Google Scholar] [CrossRef] [PubMed]
- Fernando, W.; Martins, I.J.; Goozee, K.G.; Brennan, C.S.; Jayasena, V.; Martins, R.N. The role of dietary coconut for the prevention and treatment of Alzheimer’s disease: Potential mechanisms of action. Br. J. Nutr. 2015, 114, 1–14. [Google Scholar] [CrossRef]
- Lerchenmüller, C.; Lerchenmueller, M.J.; Sorenson, O. Long-term analysis of sex differences in prestigious authorships in cardiovascular research supported by the National Institutes of Health. Circulation 2018, 137, 880–882. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, R.; Swarnamali, H.; Lanerolle, P.; Ranasinghe, P. Effect of coconut oil on cardio-metabolic risk: A systematic review and meta-analysis of interventional studies. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 14, 2007–2020. [Google Scholar] [CrossRef]
- Jayawardena, R.; Swarnamali, H.; Ranasinghe, P.; Misra, A. Health effects of coconut oil: Summary of evidence from systematic reviews and meta-analysis of interventional studies. Diabetes Metab. Syndr. Clin. Res. Rev. 2021, 15, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Xu, P.; Fan, H.; Baudouin, L.; Xia, W.; Bocs, S.; Xu, J.; Li, Q.; Guo, A.; Zhou, L.; et al. The genome draft of coconut (Cocos nucifera). Gigascience 2017, 6, gix095. [Google Scholar] [CrossRef] [PubMed]
- Harries, H.C. Coconut varieties in America. Oleagineaux 1971, 26, 235–242. [Google Scholar]
- Chan, E. Progress in coconut breeding in United Plantations Bhd, Malaysia. Oleagineaux 1983, 38, 371–376. [Google Scholar]
- Man, N.; Shah, J.A. Acceptance of new coconut seed Matag among coconut growers in Bagan Datoh, Perak and Bachok, Kelantan. Int. J. Acad. Res. Bus. Soc. Sci. 2020, 10, 1–26. [Google Scholar] [CrossRef]
- Kuhn, D.N.; Bally, I.S.E.; Dillon, N.L.; Innes, D.; Groh, A.M.; Rahaman, J.; Ophir, R.; Cohen, Y.; Sherman, A. Genetic Map of Mango: A Tool for Mango Breeding. Front. Plant Sci. 2017, 8, 577. [Google Scholar] [CrossRef]
- Zheng, S.; Hao, Y.; Fan, S.; Cai, J.; Chen, W.; Li, X.; Zhu, X. Metabolomic and transcriptomic profiling provide novel insights into fruit ripening and ripening disorder caused by 1-mcp treatments in papaya. Int. J. Mol. Sci. 2021, 22, 916. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, G.R. Genetic markers for coconut palms. In Lethal Yellowing: Research and Practical Aspects; Oropeza, C., Howard, F.W., Ashburner, G.R., Eds.; Springer: Dordrecht, The Netherlands, 1995; pp. 173–186. [Google Scholar] [CrossRef]
- Al-Khayri, J.M.; Jain, S.M.; Johnson, D.V. Advances in Plant Breeding Strategies: Fruits; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Vollmann, J.; Rajcan, I. Oil Crop Breeding and Genetics; Springer: New York, NY, USA, 2010. [Google Scholar]
- Perera, C.; Bandupriya, H.D.D.; Thomas, R.J.; Bourdeix, R. Diversity Studies Using Molecular Markers. In Coconut Biotechnology: Towards the Sustainability of the ‘Tree of Life’; Adkins, S., Foale, M., Bourdeix, R., Nguyen, Q., Biddle, J., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 101–122. [Google Scholar] [CrossRef]
- Jangra, S.; Mishra, A.; Kamboj, D.; Yadav, N.R.; Yadav, R.C. Engineering Abiotic Stress Tolerance Traits for Mitigating Climate Change. In Plant Biotechnology: Recent Advancements and Developments; Gahlawat, S.K., Salar, R.K., Siwach, P., Duhan, J.S., Kumar, S., Kaur, P., Eds.; Springer: Singapore, 2017; pp. 59–73. [Google Scholar] [CrossRef]
- Rajesh, M.K.; Ramesh, S.V.; Karun, A.; Chowdappa, P. Genome Sequencing, Transcriptomics, Proteomics and Metabolomics. In The Coconut Genome; Rajesh, M.K., Ramesh, S.V., Perera, L., Kole, C., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 119–132. [Google Scholar] [CrossRef]
- Li, D.; Fan, Y. Construction and Characterization of a cDNA Library from the Pulp of Coconut (Cocos nucifera L.). Agric. Sci. China 2008, 7, 1071–1076. [Google Scholar] [CrossRef]
- Liang, Y.; Yuan, Y.; Liu, T.; Mao, W.; Zheng, Y.; Li, D. Identification and computational annotation of genes differentially expressed in pulp development of Cocos nucifera L. by suppression subtractive hybridization. BMC Plant Biol. 2014, 14, 205. [Google Scholar] [CrossRef]
- Cutter, V.M.; Wilson, K.S.; Dubé, G.R. The isolation of living nuclei from the endosperm of Cocos nucifera. Science 1952, 115, 58–59. [Google Scholar] [CrossRef] [PubMed]
- Azam, K.; Mir, H.; Deo Prasad, B.; Ahmad, F.; Hidayatullah Mir, C. Identification of microsatellite markers associated with the horticultural traits in elite mango cultivars. J. Pharmacogn. Phytochem. 2018, 7, 2830–2834. [Google Scholar]
- Padmakar, B.; Dinesh, M.R.; Ravishankar, K.V. Marker-Trait association for fruit characters in mango (Mangifera indica L.) cultivars. J. Hortic. Sci. 2016, 11, 170–178. Available online: https://jhs.iihr.res.in/index.php/jhs/index (accessed on 9 April 2023).
- Rahman, S.; Gangaraj, K.P.; Muralikrishna, K.S.; Rajesh, M.K. Isolation and characterization of full-length coding sequences of gibberellic acid biosynthetic genes in coconut (Cocos nucifera L.) and their expression analysis. S. Afr. J. Bot. 2023, 153, 297–307. [Google Scholar] [CrossRef]
- Gao, L.; Wang, Y.; Guo, Q.; Li, D. Identification and functional analysis of transcription factors related to coconut (Cocos nucifera L.) endosperm development based on ATAC-seq. Trop. Plants 2022, 1, 8. [Google Scholar] [CrossRef]
- Sun, R.H.; Gao, L.; Mi, Z.; Zheng, Y.; Li, D. CnMADS1, a MADS transcription factor, positively modulates cell proliferation and lipid metabolism in the endosperm of coconut (Cocos nucifera L.). Planta 2020, 252, 83. [Google Scholar] [CrossRef]
- Li, D.; Zheng, Y.; Wan, L.; Zhu, X.; Wang, Z. Differentially expressed microRNAs during solid endosperm development in coconut (Cocos nucifera L.). Sci. Hortic. 2009, 122, 666–669. [Google Scholar] [CrossRef]
- Thomas, R.J.; Josephrajkumar, A. Flowering and pollination biology in coconut. J. Plant. Crops 2013, 41, 109–117. [Google Scholar]
- Gillaspy, G.; Ben-David, H.; Gruissem, W. Fruits: A Developmental Perspective. Plant Cell 1993, 5, 1439–1451. [Google Scholar] [CrossRef] [PubMed]
- Peiris, T.S.G.; Thattil, R.O.; Mahindapala, R. An Analysis of the Effect of Climate and Weather on Coconut (Cocos nucifera). Exp. Agric. 1995, 31, 451–460. [Google Scholar] [CrossRef]
- Ranasinghe, C.S.; Silva, L.R.S.; Premasiri, R.D.N. Major determinants of fruit set and yield fluctuation in coconut (Cocos nucifera L.). J. Natl. Sci. Found. 2015, 43, 253–264. [Google Scholar] [CrossRef]
- Thomas, R.J.; Nair, R.V.; Mathews, C.; Ajithkumar, R.; Sasikala, M.; Nampoothiri, C.K. Studies on fruit set in coconut upon artificial pollination in various cross combinations. Indian J. Hortic. 2012, 69, 7–12. [Google Scholar]
- Legros, S.; Mialet-Serra, I.; Caliman, J.-P.; Siregar, F.A.; Clement-Vidal, A.; Fabre, D.; Dingkuhn, M. Phenology, growth and physiological adjustments of oil palm (Elaeis guineensis) to sink limitation induced by fruit pruning. Ann. Bot. 2009, 104, 1183–1194. [Google Scholar] [CrossRef]
- Wubs, A.M.; Heuvelink, E.; Marcelis, L.F.M. Abortion of reproductive organs in sweet pepper (Capsicum annuum L.): A review. J. Hortic. Sci. Biotechnol. 2009, 84, 467–475. [Google Scholar] [CrossRef]
- Juliano, J.B. Origin, Development, and Nature of the Stony Layer of the Coconut (Cocos nucifera Linnaeus); Bureau of Printing: Manila, Philippines, 1926. [Google Scholar]
- Tammes, P.L.M. Review of coconut selection in Indonesia. Euphytica 1955, 4, 17–24. [Google Scholar] [CrossRef]
- Child, R. Coconuts, 2nd ed.; Logman: London, UK, 1974. [Google Scholar]
- Jayasuriya, V.U.D.S.; Perera, R. Growth, development and dry matter accumulation in the fruit of Cocos nucifera L. var nana form pumila. Cocos 1985, 3, 16–21. [Google Scholar] [CrossRef]
- Niral, V.; Nair, R.V.; Jerard, B.A.; Samsudeen, K.; Ratnambal, M.J. Evaluation of coconut germplasm for fruit component traits and oil yield. J. Oilseeds Res. 2009, 26, 668–670. [Google Scholar]
- Prades, A.; Dornier, M.; Diop, N.; Pain, J.-P. Coconut water preservation and processing: A review. Fruits 2012, 67, 157–171. [Google Scholar] [CrossRef]
- Javel, I.M.; Bandala, A.A.; Salvador, R.C.; Bedruz, R.A.R.; Dadios, E.P.; Vicerra, R.R.P. Coconut Fruit Maturity Classification using Fuzzy Logic. In Proceedings of the 2018 IEEE 10th International Conference on Humanoid, Nanotechnology, Information Technology, Communication and Control, Environment and Management (HNICEM), Baguio City, Philippines, 29 November–2 December 2018; pp. 1–6. [Google Scholar] [CrossRef]
- Shutterstock. Available online: https://www.https://www.shutterstock.com/image-photo/flower-coconut-tree-isolated-on-white-273312014 (accessed on 23 December 2022).
- Biology—The World of Life Home Page. Available online: https://velasserilblog.wordpress.com/chapter-2-sexual-reproduction-in-flowering-plants/ (accessed on 23 December 2022).
- Dreamstime Home Page. Available online: https://www.dreamstime.com/flowers-coconut-flower-both-male-female-same-flower-coconuts-fragrant-flowers-can-image120449119 (accessed on 24 December 2022).
- Waidyarathne, K.P.; Chandrathilake, T.H.; Wickramarachchi, W.S. Application of artificial neural network to predict copra conversion factor. Neural Comput. Applic. 2022, 34, 7909–7918. [Google Scholar] [CrossRef]
- Mohd Lazim, M.I.; Badruzaman, N.A. Quantification of Cytokinins in Coconut Water from Different Maturation Stages of Malaysia’s Coconut (Cocos nucifera L.) Varieties. J. Food Process. Technol. 2015, 6, 11. [Google Scholar] [CrossRef]
- Prades, A.; Dornier, M.; Diop, N.; Pain, J.-P. Coconut water uses, composition and properties: A review. Fruits 2012, 67, 87–107. [Google Scholar] [CrossRef]
- Alamy Home Page. Available online: https://www.alamy.com/stock-photo/coconut-water-cut-out.html?sortBy=relevant (accessed on 25 December 2022).
- Sudha, R.; Rajesh, M.; Niral, V.; Samsudeen, K.; Shil, S.; Ramchander, S.; Diwakar, Y. Analysis of genetic diversity and population structure in worldwide coconut germplasm (Cocos nucifera L.) using microsatellite markers. Sci. Hortic. 2023, 309, 111681. [Google Scholar] [CrossRef]
- Preethi, P.; Rahman, S.; Naganeeswaran, S.; Sabana, A.A.; Gangaraj, K.P.; Jerard, B.A.; Niral, V.; Rajesh, M.K. Development of EST-SSR markers for genetic diversity analysis in coconut (Cocos nucifera L.). Mol. Biol. Rep. 2020, 47, 9385–9397. [Google Scholar] [CrossRef]
- Kamaral, L.C.J.; Perera, S.; Perera, K.; Dassanayaka, P.N. Characterisation of Sri Lanka yellow dwarf coconut (Cocos nucifera L.) by DNA fingerprinting with SSR markers. J. Natl. Sci. Found. Sri Lanka 2017, 45, 405. [Google Scholar] [CrossRef]
- Azevedo, A.O.N.; Azevedo, C.D.D.O.; Santos, P.H.A.D.; Ramos, H.C.C.; Boechat, M.S.B.; Arêdes, F.A.S.; Ramos, S.R.R.; Mirizola, L.; Perera, L.; Aragão, W.M.; et al. Selection of legitimate dwarf coconut hybrid seedlings using DNA fingerprinting. Crop Breed. Appl. Biotechnol. 2018, 18, 409–416. [Google Scholar] [CrossRef]
- Rajesh, M.K.; Gangurde, S.S.; Pandey, M.K.; Niral, V.; Sudha, R.; Jerard, B.A.; Kadke, G.N.; Sabana, A.A.; Muralikrishna, K.S.; Samsudeen, K.; et al. Insights on genetic diversity, population structure, and linkage disequilibrium in globally diverse coconut accessions using genotyping-by-sequencing. OMICS 2021, 25, 796–809. [Google Scholar] [CrossRef]
- Muñoz-Pérez, J.M.; Cañas, G.P.; López, L.; Arias, T. Genome-wide diversity analysis to infer population structure and linkage disequilibrium among Colombian coconut germplasm. Sci. Rep. 2022, 12, 2958. [Google Scholar] [CrossRef]
- Novarianto, H.; Tulalo, M.A. Performance of Coconut Hybrids Dwarf x Bido Tall Penampilan Kelapa Hibrida Genjah x Dalam Bido. Bul. Palma Vol. 2021, 22, 1–10. [Google Scholar] [CrossRef]
- Sudha, R.; Samsudeen, K.; Rajesh, M.K.; Niral, V. Molecular marker assisted confirmation of hybrids in coconut (Cocos nucifera L.). Indian J. Genet. Plant Breed. 2022, 82, 369–372. [Google Scholar] [CrossRef]
- Upadhyay, A.; Jayadev, K.; Manimekalai, R.; Parthasarathy, V.A. Genetic relationship and diversity in Indian coconut accessions based on RAPD markers. Sci. Hortic. 2004, 99, 353–362. [Google Scholar] [CrossRef]
- Wadt, L.H.O.; Sakiyama, N.S.; Pereira, M.G.; Tupinamba, E.A.; Ribeiro, F.E.; Aragao, W.M. RAPD markers in the genetic diversity study of the coconut palm. Curr. Adv. Coconut Biotechnol. 1999, 35, 89–97. [Google Scholar] [CrossRef]
- Paul, R.; George, J.; Rajesh, M.K.; Jerard, B.A.; Niral, V. Genetic diversity in yellow dwarf populations of coconut assessed using RAPD markers. Indian J. Hortic. 2008, 65, 60–64. [Google Scholar]
- Masumbuko, L.I.; Sinje, S.; Kullaya, A. Genetic diversity and structure of East African tall coconuts in Tanzania using RAPD markers. Open J. Genet. 2014, 4, 175–181. [Google Scholar] [CrossRef]
- Manimekalai, R. Informative Inter Simple Sequence Repeat (ISSR) primers for genetic analysis of coconut (Cocos nucifera L.) germplasm. CORD 2005, 21, 34. [Google Scholar] [CrossRef]
- Manimekalai, R. Inter Simple Sequence Repeat (ISSR) markers for identification of germplasm and diversity analysis in coconut (Cocos nucifera L.). Int. J. Innov. Hortic. 2012, 1, 63–67. [Google Scholar]
- Mahayu, W.M. Taryono Coconut (Cocos nucifera L.) diversity in Indonesia based on SSR molecular marker. AIP Conf. Proc. AIP Publ. LLC 2019, 2099, 020013. [Google Scholar] [CrossRef]
- Rahayu, M.S.; Setiawan, A.; Maskromo, I.; Purwito, A.; Sudarsono, S. Genetic diversity analysis of Puan Kalianda kopyor coconuts (Cocos nucifera) from South Lampung, Indonesia based on SSR markers. Biodiversitas 2022, 23, 1. [Google Scholar] [CrossRef]
- Oyoo, M.E.; Muhammed, N.; Cyrus, K.N.; Githiri, S.M. Assessment of the genetic diversity of Kenyan coconut germplasm using simple sequence repeat (SSR) markers. Afr. J. Biotechnol. 2016, 15, 2215–2223. [Google Scholar] [CrossRef]
- Martinez, R.T.; Baudouin, L.; Berger, A.; Dollet, M. Characterization of the genetic diversity of the tall coconut (Cocos nucifera L.) in the Dominican Republic using microsatellite (SSR) markers. Tree Genet. Genomes 2010, 6, 73–81. [Google Scholar] [CrossRef]
- Liu, X.; Tang, H.; Li, D.; Hou, L. Genetic diversity of coconut cultivars in China by microsatellite (SSR) markers. Mol. Plant Breed. 2011, 2, 12. [Google Scholar] [CrossRef]
- Geethanjali, S.; Rukmani, J.A.; Rajakumar, D.; Kadirvel, P.; Viswanathan, P.L. Genetic diversity, population structure and association analysis in coconut (Cocos nucifera L.) germplasm using SSR markers. Plant Genet. Resour. 2018, 16, 156–168. [Google Scholar] [CrossRef]
- Kandoliya, U.K.; Joshi, A.; Mori, D.S.; Marviya, G.V.; Golakiya, B.A. Genetic Diversity Analysis of Coconut (Cocos nucifera L.) Genotypes and Hybrids Using Molecular Marker. Indian J. Agric. Biochem. 2018, 31, 25–32. [Google Scholar] [CrossRef]
- Jerard, B.A.; Rajesh, M.K.; Thomas, R.J.; Niral, V.; Samsudeen, K. Island Ecosystems Host Rich Diversity in Coconut (Cocos nucifera): Evidences from Minicoy Island, India. Agric. Res. 2017, 6, 214–226. [Google Scholar] [CrossRef]
- Rasam, D.V.; Gokhale, N.B.; Sawardekar, S.V.; Patil, D.M. Molecular characterisation of coconut (Cocos nucifera L.) varieties using ISSR and SSR markers. J. Hortic Sci. Biotechnol. 2016, 91, 347–352. [Google Scholar] [CrossRef]
- Mauro-Herrera, M.; Meerow, A.W.; Borrone, J.W.; Kuhn, D.N.; Schnell, R.J. Ten informative markers developed from WRKY sequences in coconut (Cocos nucifera). Mol. Ecol. Notes 2006, 6, 904–906. [Google Scholar] [CrossRef]
- Yates, C.M.; Sternberg, M.J.E. Proteins and Domains Vary in Their Tolerance of Non-Synonymous Single Nucleotide Polymorphisms (nsSNPs). J. Mol. Biol. 2013, 425, 1274–1286. [Google Scholar] [CrossRef]
- Santos, P.H.A.D.; Venâncio, T.M.; Ramos, H.C.C.; Arêdes, F.A.S.; Azevedo, A.O.N.; Boechat, M.S.B.; Filho, G.A.D.S.; Ramos, S.R.R.; Mirisola, L.A.; Aragão, W.M.; et al. Genotyping-by-sequencing technology reveals directions for coconut (Cocos nucifera L.) breeding strategies for water production. Euphytica 2020, 216, 45. [Google Scholar] [CrossRef]
- Muñoz-Pérez, J.M.; Cañas, G.P.; López, L.; Arias, T. Genome-wide diversity of coconut from northern South America uncovers genotypes present in Colombia and strong population structure. bioRxiv 2020, 2020, 825398. [Google Scholar] [CrossRef]
- Pesik, A.; Efendi, D.; Novarianto, H.; Dinarti, D.; Sudarsono, S. Development of SNAP markers based on nucleotide variability of WRKY genes in coconut and their validation using multiplex PCR. Biodiversitas 2017, 18, 465–475. [Google Scholar] [CrossRef]
- Rajesh, M.K.; Ramesh, S.V.; Perera, L.; Manickavelu, A. Quantitative Trait Loci (QTL) and Association Mapping for Major Agronomic Traits. In The Coconut Genome; Springer: Berlin/Heidelberg, Germany, 2021; pp. 91–101. [Google Scholar] [CrossRef]
- Rajesh, M.K.; Karun, A.; Parthasarathy, V.A. Coconut Biotechnology. In The Coconut Palm (Cocos nucifera L.)—Research and Development Perspectives; Krishnakumar, V., Thampan, P.K., Nair, M.A., Eds.; Springer: Singapore, 2018; pp. 191–226. [Google Scholar] [CrossRef]
- Jaccoud, D.; Peng, K.; Feinstein, D.; Kilian, A. Diversity Arrays: A solid-state technology for sequence information independent genotyping. Nucleic Acids Res. 2001, 29, e25. [Google Scholar] [CrossRef]
- Baudouin, L.; Lebrun, A.P.; Konan, A.J.L.; Ritter, A.E.; Berger, A.; Billotte, A.N. QTL analysis of fruit components in the progeny of a Rennell Island Tall coconut (Cocos nucifera L.) individual. Theor. Appl. Genet. 2006, 112, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Riedel, M.; Riederer, M.; Becker, D.; Herran, A.; Kullaya, A.; Arana-López, G.; Peña-Rodríguez, L.; Billotte, N.; Sniady, V.; Rohde, W.; et al. Cuticular wax composition in Cocos nucifera L.: Physicochemical analysis of wax components and mapping of their QTLs onto the coconut molecular linkage map. Tree Genet. Genomes 2009, 5, 53–69. [Google Scholar] [CrossRef]
- Zhou, L.; Yarra, R.; Cao, H. SSR based association mapping analysis for fatty acid content in coconut flesh and exploration of the elite alleles in Cocos nucifera L. Curr. Plant Biol. 2020, 21, 100141. [Google Scholar] [CrossRef]
- Gunn, B.F.; Baudouin, L.; Beulé, T.; Ilbert, P.; Duperray, C.; Crisp, M.; Issali, A.; Konan, J.-L.; Rival, A. Ploidy and domestication are associated with genome size variation in Palms. Am. J. Bot. 2015, 102, 1625–1633. [Google Scholar] [CrossRef]
- Alsaihati, B.; Liu, W.; Lin, Q.; Al-Mssallem, I.S. Coconut genome de novo sequencing. In Proceedings of the Plant and Animal Genome XXII Conference, San Diego, CA, USA, 12–17 January 2014; Available online: http://www.intpag.org/ (accessed on 30 May 2015).
- Yang, Y.; Bocs, S.; Fan, H.; Armero, A.; Baudouin, L.; Xu, P.; Xu, J.; This, D.; Hamelin, C.; Iqbal, A.; et al. Coconut genome assembly enables evolutionary analysis of palms and highlights signaling pathways involved in salt tolerance. Commun. Biol. 2021, 4, 105. [Google Scholar] [CrossRef]
- Bandupriya, H.D.D.; Perera, S.A.C.N. Coconut Genomics. In Oil Crop Genomics; Springer: Cham, Switzerland, 2021; pp. 161–188. [Google Scholar] [CrossRef]
- Singh, R.; Ong-Abdullah, M.; Low, E.-T.L.; Manaf, M.A.A.; Rosli, R.; Nookiah, R.; Ooi, L.C.-L.; Ooi, S.-E.; Chan, K.-L.; Halim, M.A.; et al. Oil palm genome sequence reveals divergence of interfertile species in old and new worlds. Nature 2013, 500, 335–339. [Google Scholar] [CrossRef]
- Al-Mssallem, I.S.; Hu, S.; Zhang, X.; Lin, Q.; Liu, W.; Tan, J.; Yu, X.; Liu, J.; Pan, L.; Zhang, T.; et al. Genome sequence of the date palm Phoenix dactylifera L. Nat. Commun 2013, 4, 2274. [Google Scholar] [CrossRef]
- Al-Dous, E.K.; George, B.; Al-Mahmoud, M.E.; Al-Jaber, M.Y.; Wang, H.; Salameh, Y.M.; Al-Azwani, E.K.; Chaluvadi, S.; Pontaroli, A.C.; DeBarry, J.; et al. De novo genome sequencing and comparative genomics of date palm (Phoenix dactylifera). Nat. Biotechnol. 2011, 29, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Lantican, D.V.; Strickler, S.R.; Canama, A.O.; Gardoce, R.R.; Mueller, L.A.; Galvez, H.F. De Novo Genome Sequence Assembly of Dwarf Coconut (Cocos nucifera L. ’Catigan Green Dwarf’) Provides Insights into Genomic Variation between Coconut Types and Related Palm Species. G3 (Bethesda) 2019, 9, 2377–2393. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.V.; Jacob, P.M.; Thomas, R.J.; Sasikala, M. Development of varieties of coconut Cocos nucifera D resistant. J. Plant. GGGG 2004, 32, 33. [Google Scholar]
- Thomas, R.J.; Rajesh, M.K.; Jacob, P.M.; Jose, M.; Nair, R.V. Studies on genetic uniformity of Chowghat Green Dwarf and Malayan Green Dwarf varieties of coconut using molecular and morphometric methods. J. Plant. Crops 2015, 43, 89–96. [Google Scholar]
- Muliyar, R.K.; Chowdappa, P.; Behera, S.K.; Kasaragod, S.; Gangaraj, K.P.; Kotimoole, C.N.; Nekrakalaya, B.; Mohanty, V.; Sampgod, R.B.; Banerjee, G.; et al. Assembly and Annotation of the Nuclear and Organellar Genomes of a Dwarf Coconut (Chowghat Green Dwarf) Possessing Enhanced Disease Resistance. OMICS 2020, 24, 726–742. [Google Scholar] [CrossRef]
- Wang, S.; Xiao, Y.; Zhou, Z.-W.; Yuan, J.; Guo, H.; Yang, Z.; Yang, J.; Sun, P.; Sun, L.; Deng, Y.; et al. High-quality reference genome sequences of two coconut cultivars provide insights into evolution of monocot chromosomes and differentiation of fiber content and plant height. Genome Biol. 2021, 22, 304. [Google Scholar] [CrossRef]
- Guerin, C.; Serret, J.; Montúfar, R.; Vaissayre, V.; Bastos-Siqueira, A.; Durand-Gasselin, T.; Tregear, J.; Morcillo, F.; Dussert, S. Palm seed and fruit lipid composition: Phylogenetic and ecological perspectives. Ann. Bot. 2020, 125, 157–172. [Google Scholar] [CrossRef]
- Xiao, Y.; Xia, W.; Mason, A.S.; Cao, Z.; Fan, H.; Zhang, B.; Zhang, J.; Ma, Z.; Peng, M.; Huang, D. Genetic control of fatty acid composition in coconut (Cocos nucifera), African oil palm (Elaeis guineensis), and date palm (Phoenix dactylifera). Planta 2019, 249, 333–350. [Google Scholar] [CrossRef]
- Manohar, A.N.; Lantican, D.V.; Dancel, M.P.; Cardona, D.E.; Ibarra, A.C.; Gulay, C.R.; Canama, A.O.; Gardoce, R.R.; Galvez, H.F. Genome-guided molecular characterization of oil genes in coconut (Cocos nucifera L.). Philipp. J. Sci. 2019, 148, 153–164. [Google Scholar]
- Huang, Y.Y.; Matzke, A.J.M.; Matzke, M. Complete Sequence and Comparative Analysis of the Chloroplast Genome of Coconut Palm (Cocos nucifera). PLoS ONE 2013, 8, e74736. [Google Scholar] [CrossRef]
- Uthaipaisanwong, P.; Chanprasert, J.; Shearman, J.; Sangsrakru, D.; Yoocha, T.; Jomchai, N.; Jantasuriyarat, C.; Tragoonrung, S.; Tangphatsornruang, S. Characterization of the chloroplast genome sequence of oil palm (Elaeis guineensis Jacq.). Gene 2012, 500, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Zhang, X.; Liu, G.; Yin, Y.; Chen, K.; Yun, Q.; Zhao, D.; Al-Mssallem, I.S.; Yu, J. The complete chloroplast genome sequence of date palm (Phoenix dactylifera L.). PLoS ONE 2010, 5, e12762. [Google Scholar] [CrossRef] [PubMed]
- Aljohi, H.A.; Liu, W.; Lin, Q.; Zhao, Y.; Zeng, J.; Alamer, A.; Alanazi, I.O.; Alawad, A.O.; Al-Sadi, A.M.; Hu, S.; et al. Complete Sequence and Analysis of Coconut Palm (Cocos nucifera) Mitochondrial Genome. PLoS ONE 2016, 11, e0163990. [Google Scholar] [CrossRef]
- Fang, Y.; Wu, H.; Zhang, T.; Yang, M.; Yin, Y.; Pan, L.; Yu, X.; Zhang, X.; Hu, S.; Al-Mssallem, I.S.; et al. A complete sequence and transcriptomic analyses of date palm (Phoenix dactylifera L.) mitochondrial genome. PLoS ONE 2012, 7, e37164. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Luo, Y.; Yang, Y.; Fan, H.; Xia, W.; Mason, A.S.; Zhao, S.; Sager, R.; Qiao, F. Development of microsatellite markers in Cocos nucifera and their application in evaluating the level of genetic diversity of Cocos nucifera. Plant Omics 2013, 6, 193–200. [Google Scholar]
- Rethinam, P.; Krishnakumar, V. Tender Coconut Varieties. In Coconut Water: A Promising Natural Health Drink-Distribution, Processing and Nutritional Benefits; Rethinam, P., Krishnakumar, V., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 37–76. [Google Scholar] [CrossRef]
- Saensuk, C.; Wanchana, S.; Choowongkomon, K.; Wongpornchai, S.; Kraithong, T.; Imsabai, W.; Chaichoompu, E.; Ruanjaichon, V.; Toojinda, T.; Vanavichit, A.; et al. De novo transcriptome assembly and identification of the gene conferring a “pandan-like” aroma in coconut (Cocos nucifera L.). Plant Sci. 2016, 252, 324–334. [Google Scholar] [CrossRef]
- Niral, V.; Augustine, B.; Samsudeen, J.K.; Thomas, R.J.; Thomas, G.V. Coconut Varieties and Hybrids; Central Plantation Crops Research Institute, Indian Council of Agricultural Research: Kasaragod, India, 2014. [Google Scholar]
- Ramesh, S.V.; Arunachalam, V.; Rajesh, M.K. Genomic Designing of Climate-Smart Coconut. In Genomic Designing of Climate-Smart Fruit Crops; Kole, C., Ed.; Springer: Cham, Switzerland, 2020; pp. 135–156. [Google Scholar] [CrossRef]
- Bourdeix, R.; Batugal, P. Standardized Catalogues of Coconut Germplasm: Catalogue of Conserved Germplasm and Farmers’ Varieties; IPGRI: Rome, Italy, 2005. [Google Scholar]
- Ratnambal, M.J.; Kumaran, P.M.; Bashkara Rao, E.V.V.; Pillai, R.V. Catalog of Conserved Coconut Germplasm-by Country of Origin-India; International Coconut Genetic Resources Network (COGENT), Bioversity International, Regional Office for Asia, the Pacific and Oceania: Serdang, Malaysia, 2010; pp. 86–91. [Google Scholar]
- Ratnambal, M.J.; Kumaran, P.M.; Bashkara Rao, E.V.V.; Pillai, R.V. Catalogue of Conserved Coconut Germplasm by Country of Origin-Philippine; International Coconut Genetic Resources Network (COGENT), Bioversity International, Regional Office for Asia, the Pacific and Oceania: Serdang, Malaysia, 2010; pp. 218–223. [Google Scholar]
- Rivera, R.; Santos, G.; Emmanuel, E.; Rivera, S. Catalog of Conserved Coconut Germplasm-by Country of Origin-Philippines Baybay Tall (BAYT) in the Philippines; International Coconut Genetic Resources Network (COGENT), Bioversity International, Regional Office for Asia, the Pacific and Oceania: Serdang, Malaysia, 2010; pp. 212–217. [Google Scholar]
- Adobe Stock. Available online: https://stock.adobe.com/sg/search?k=%22aromatic+coconut%22&asset_id=361589161 (accessed on 7 March 2023).
- Appaiah, P.; Sunil, L.; Prasanth Kumar, P.K.; Gopala Krishna, A.G. Composition of coconut testa, coconut kernel and its oil. J. Am. Oil Chem. Soc. 2014, 91, 917–924. [Google Scholar] [CrossRef]
- Benjakul, S.; Patil, U.; Prodpran, T.; Senphan, T.; Cheetangdee, N. A comparative study of the physicochemical properties and emulsion stability of coconut milk at different maturity stages. Ital. J. Food Sci. 2017, 29, 1. [Google Scholar]
- Tuyekar, S.T.; Tawade, B.S.; Singh, K.S.; Wagh, V.S.; Vidhate, P.K.; Yevale, R.P.; Gaikwad, S.; Kale, M. An Overview on Coconut Water: As a Multipurpose Nutrition. Int. J. Pharm. Sci. Rev. Res. 2021, 68, 63–70. [Google Scholar] [CrossRef]
- Boateng, L.; Ansong, R.; Owusu, W.; Steiner-Asiedu, M. Coconut oil and palm oil’s role in nutrition, health and national development: A review. Ghana Med. J. 2016, 50, 189–196. [Google Scholar] [CrossRef]
- Ghosh, P.K.; Bhattacharjee, P.; Mitra, S.; Poddar-Sarkar, M. Physicochemical and phytochemical analyses of copra and oil of Cocos nucifera L. (West Coast Tall Variety). Int. J. Food Sci. 2014, 2014, 310852. [Google Scholar] [CrossRef] [PubMed]
- McKeon, T.A.; Hayes, D.G.; Hildebrand, D.F.; Weselake, R.J. Introduction to industrial oil crops. In Industrial Oil Crops; Elsevier: Amsterdam, The Netherlands, 2016; pp. 1–13. [Google Scholar] [CrossRef]
- Fan, H.; Xiao, Y.; Yang, Y.; Xia, W.; Mason, A.S.; Xia, Z.; Qiao, F.; Zhao, S.; Tang, H. RNA-Seq analysis of Cocos nucifera: Transcriptome sequencing and de novo assembly for subsequent functional genomics approaches. PLoS ONE 2013, 8, e59997. [Google Scholar] [CrossRef] [PubMed]
- Ann Osio, C.L.; Mae Aquino, G.B.; Aala, W.F., Jr.; Lado, J.P.; Jo Tendero, B.T.; Cueto, C.A.; Diaz, M.G.Q.; Laude, R.P. Transcriptome Analysis of “Philippine Lono Tall” Coconut (Cocos nucifera L.) Endosperm Reveals Differential Expression of Genes Involved in Oil Biosynthesis. Philipp. Agric. Sci. 2019, 102, 101–118. [Google Scholar]
- Punzalan, M.a.R.; Cabria, G.L.; Bautista, M.; Emmanuel, E.; Rivera, R.; Rivera, S.; Saloma, C. Differential Expression Analysis in High-yielding and Low-yielding Philippine Coconut through Transcriptome Sequencing. Philipp. J. Sci. 2019, 148, 83–95. [Google Scholar]
- Huang, Y.Y.; Lee, C.P.; Fu, J.L.; Chang, B.C.; Matzke, A.J.; Matzke, M. De novo transcriptome sequence assembly from coconut leaves and seeds with a focus on factors involved in RNA-directed DNA methylation. G3 (Bethesda) 2014, 4, 2147–2157. [Google Scholar] [CrossRef]
- Bandupriya, H.D.D.; Dunwell, J.M. Transcriptome analysis for discovering candidate genes involve in embryogenesis in coconut (Cocos nucifera L.) through 454 pyrosequencing. J. Natl. Sci. Found. 2015, 43, 319. [Google Scholar] [CrossRef]
- Punzalan, M.a.R.; Bautista, M.; Emmanuel, E.; Rivera, R.; Rivera, S.; Saloma, C. Transcriptome of the Traditional Coconut Variety Laguna Tall. Philipp. J. Sci. 2019, 148, 153–164. [Google Scholar]
- Rajesh, M.K.; Fayas, T.P.; Naganeeswaran, S.; Rachana, K.E.; Bhavyashree, U.; Sajini, K.K.; Karun, A. De novo assembly and characterization of global transcriptome of coconut palm (Cocos nucifera L.) embryogenic calli using Illumina paired-end sequencing. Protoplasma 2016, 253, 913–928. [Google Scholar] [CrossRef]
- Sabana, A.A.; Antony, G.; Rahul, C.U.; Rajesh, M.K. In silico identification of microRNAs and their targets associated with coconut embryogenic calli. Agri Gene 2018, 7, 59–65. [Google Scholar] [CrossRef]
- Sabana, A.A.; Rajesh, M.K.; Antony, G. Dynamic changes in the expression pattern of miRNAs and associated target genes during coconut somatic embryogenesis. Planta 2020, 251, 79. [Google Scholar] [CrossRef]
- Nejat, N.; Sijam, K.; Abdullah, S.N.A.; Vadamalai, G.; Dickinson, M. Phytoplasmas associated with disease of coconut in Malaysia: Phylogenetic groups and host plant species. Plant Pathol 2009, 58, 1152–1160. [Google Scholar] [CrossRef]
- Nejat, N.; Cahill, D.M.; Vadamalai, G.; Ziemann, M.; Rookes, J.; Naderali, N. Transcriptomics-based analysis using RNA-Seq of the coconut (Cocos nucifera) leaf in response to yellow decline phytoplasma infection. Mol. Genet. Genom. 2015, 290, 1899–1910. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.K.; Jasrotia, R.S.; Iquebal, M.; Jaiswal, S.; Angadi, U.; Rai, A.; Kumar, D. Deciphering genes associated with root wilt disease of coconut and development of its transcriptomic database (CnTDB). Physiol. Mol. Plant Pathol. 2017, 100, 255–263. [Google Scholar] [CrossRef]
- Silverio-Gómez, C.; Vega-Arreguín, J.; Nic-Matos, G.; Narváez-Cab, M.; Sáenz-Carbonell, L.; Oropeza, C. Transcriptome analysis reveals key defense-related genes upon SA induction in Cocos nucifera L. Genes Genom. 2022, 44, 197–210. [Google Scholar] [CrossRef]
- Rajesh, M.K.; Rachana, K.E.; Naganeeswaran, S.A.; Shafeeq, R.; Thomas, R.J.; Shareefa, M.; Merin, B.; Anitha, K. Identification of expressed resistance gene analog sequences in coconut leaf transcriptome and their evolutionary analysis. Turk. J. Agric. For. 2015, 39, 489–502. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Trait | Marker | Locus | Genomic Position (Mb) | Phenotypic Effect Mean | Frequency | Carrier Germplasm | Reference |
|---|---|---|---|---|---|---|---|
| Fatty acid content | SSR-C-7566-3 | CnfatB3-359 | Chr11:12.36 | +2.35 | 0.53 | Aromatica Green Dwarf | [84] |
| Fatty acid content | SSR-C-7566-2 | CnFatB2-830 | Chr11:18.33 | −0.65 | 0.15 | MAWA | [84] |
| Parameter (Variety) | Sequencing Platform | Estimated Genome Size (Gb) | Assembly Size (Gb) | Sequence Count | Scaffold N50 (Kb) | Scaffold Presenting Percentage of Genome | Seq Coverage | Contig N50 (Kb) | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Hainan Tall | Ilumina HiSeq2000 | 2.42 | 2.2 | 1,11,366 | 418.07 | 90.91% | ×173 | 72.64 | [9] |
| Catigan Green Dwarf | PacBio-SMRT+Illumina Miseq | 2.15 | 2.10 | 7998 | 151.93 | 97.6% | ×175.91 | 46.36 | [92] |
| Chowghat Green Dwarf | Ilumina HiSeq2000 +PacBioRSII | 1.94 | 1.93 | 26,885 | 128.74 | 81.56% | ×70.6 | 48 | [95] |
| Cn. Tall | nanopore sequencing+Hi-C +Illumina HiSeq2000 | 2.42 | 2.4 | 34,251,876 | - | 83.55% | - | 2927 | [96] |
| Cn. dwarf | 2.44 | 2.39 | 56,639,188 | - | 84.00% | - | 14,296 |
| Variety | Organs Used in Sequencing | Whole Nucleotide | Pair-End Reads | Clean Bases | Number of Unigenes | N50 Value of Unigenes | Total Transcripts | Average Length of Unigenes | Percentage of Gene Predicted from Genome-Seq Match to Transcriptome | Reference |
|---|---|---|---|---|---|---|---|---|---|---|
| Hainan Tall | 8-month Endosperm and Leaf | 4.9 (Gb) | 54,931,406 | Not Mentioned | 57,304 | 1219 | Not Mentioned | 752 | 20,541 (71%) | [121] |
| Green Dwarf | 8-month Embryo 5-month Endosperm Spear Leaf | 10 (Gb) | 81,128,552 103,080,366 121,151,552 | 12,435,008,760 9,682,020,834 13,609,335,965 | 58,211 61,159 33,446 | 951 969 912 | 86,254 229,886 159,509 | 732 684 744 | 24,857 (42.7%) 29,731 (48.6%) 26,069 (47.9%) | [124] |
| West Coast Tall | 12-month zygotic embryo along with endosperm | 7.73 (Gb) | 50,839,994 | 5.42 (Gb) | 73,308 | 561 | 161,426 | 436 | 40,367 (54.84%) | [127] |
| PLNT & LAGT | 6–7-month Endosperm | 4 (Gb) | 395,511,022 | 226,653,479 | 416,488 | 874 | 267,827(PLNT) and 285,037 (LAGT) | 406(PLNT) and 736 (LAGT) | 436 (19%) | [122] |
| Green Dwarf | 7-month endosperm (Aromatic and Non-Aromatic) | 11 (Gb) | 54,884,736 (Aromatic) and 55,031,246 (non-aromatic) | 5,487,475,543 (Aromatic) and 5,502,110,501 (non-aromatic) | 24,572 (Aromatic) and 23,322 (non-aromatic) | 1074 (Aromatic) and 1134 (non-aromatic) | 118,221 (Aromatic) and 95,613 (non-aromatic) | 653 (Aromatic) and 662 (non-aromatic) | N | [107] |
| Baybay Tall (BAYT) | 7-month Endosperm and Endocarp | 66.9–88.9 (Mb) | 586,631,950 | 334,181,119 | 95,474 (EC) and 103,979 (ES) | 270–2287 | 234,588 (EC) and 284,696 (ES) | 80.3–285.9 | 35,650 (28.1%) | [123] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yousefi, K.; Abdullah, S.N.A.; Hatta, M.A.M.; Ling, K.L. Genomics and Transcriptomics Reveal Genetic Contribution to Population Diversity and Specific Traits in Coconut. Plants 2023, 12, 1913. https://doi.org/10.3390/plants12091913
Yousefi K, Abdullah SNA, Hatta MAM, Ling KL. Genomics and Transcriptomics Reveal Genetic Contribution to Population Diversity and Specific Traits in Coconut. Plants. 2023; 12(9):1913. https://doi.org/10.3390/plants12091913
Chicago/Turabian StyleYousefi, Kobra, Siti Nor Akmar Abdullah, Muhammad Asyraf Md Hatta, and Kong Lih Ling. 2023. "Genomics and Transcriptomics Reveal Genetic Contribution to Population Diversity and Specific Traits in Coconut" Plants 12, no. 9: 1913. https://doi.org/10.3390/plants12091913