
Optimizing Insecticide Application Timing for Broad Bean Weevil Control and Minimizing Crop Damage in Broad Bean (Vicia faba Linn.)


Abstract
:1. Introduction
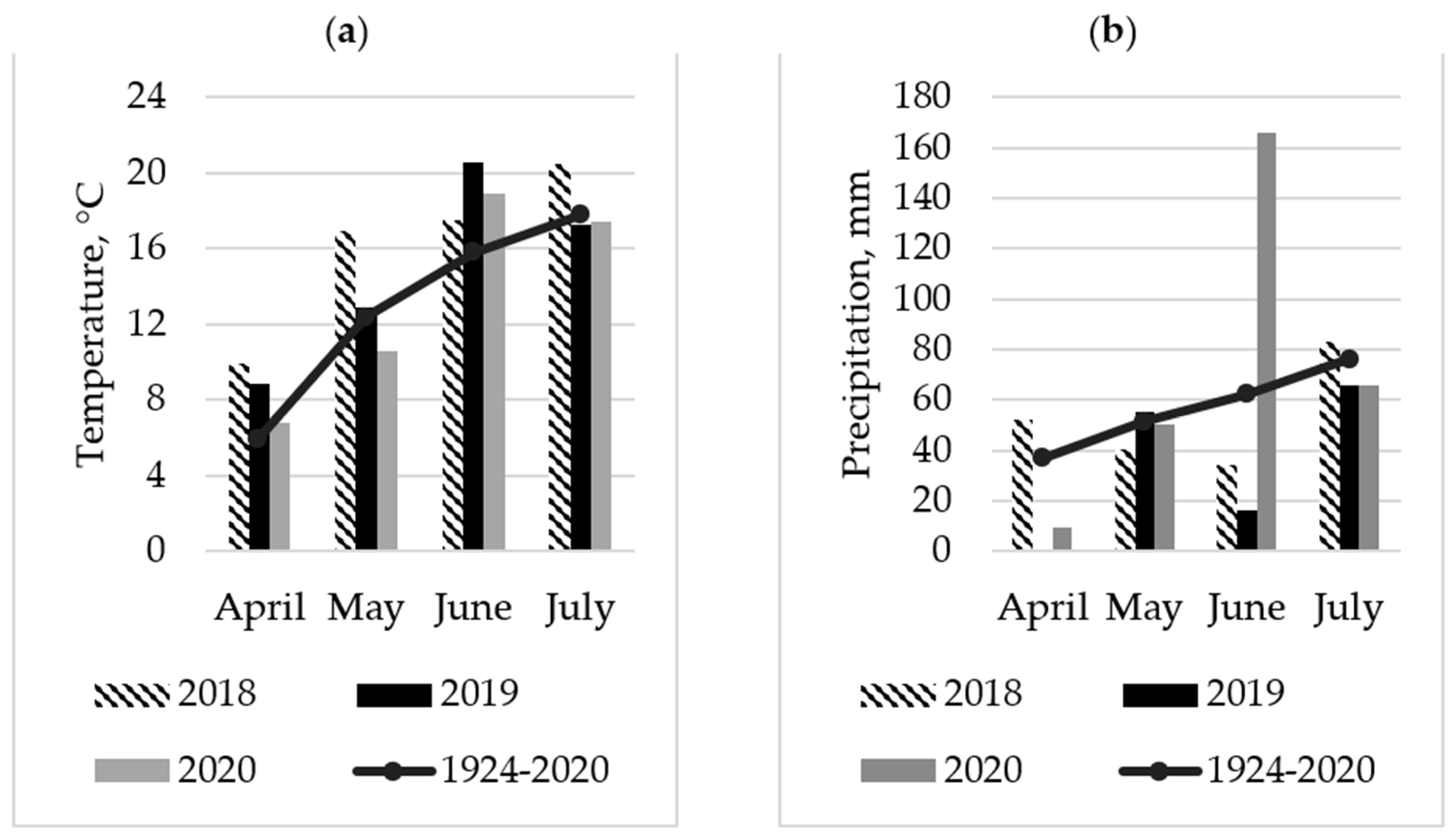
2. Results
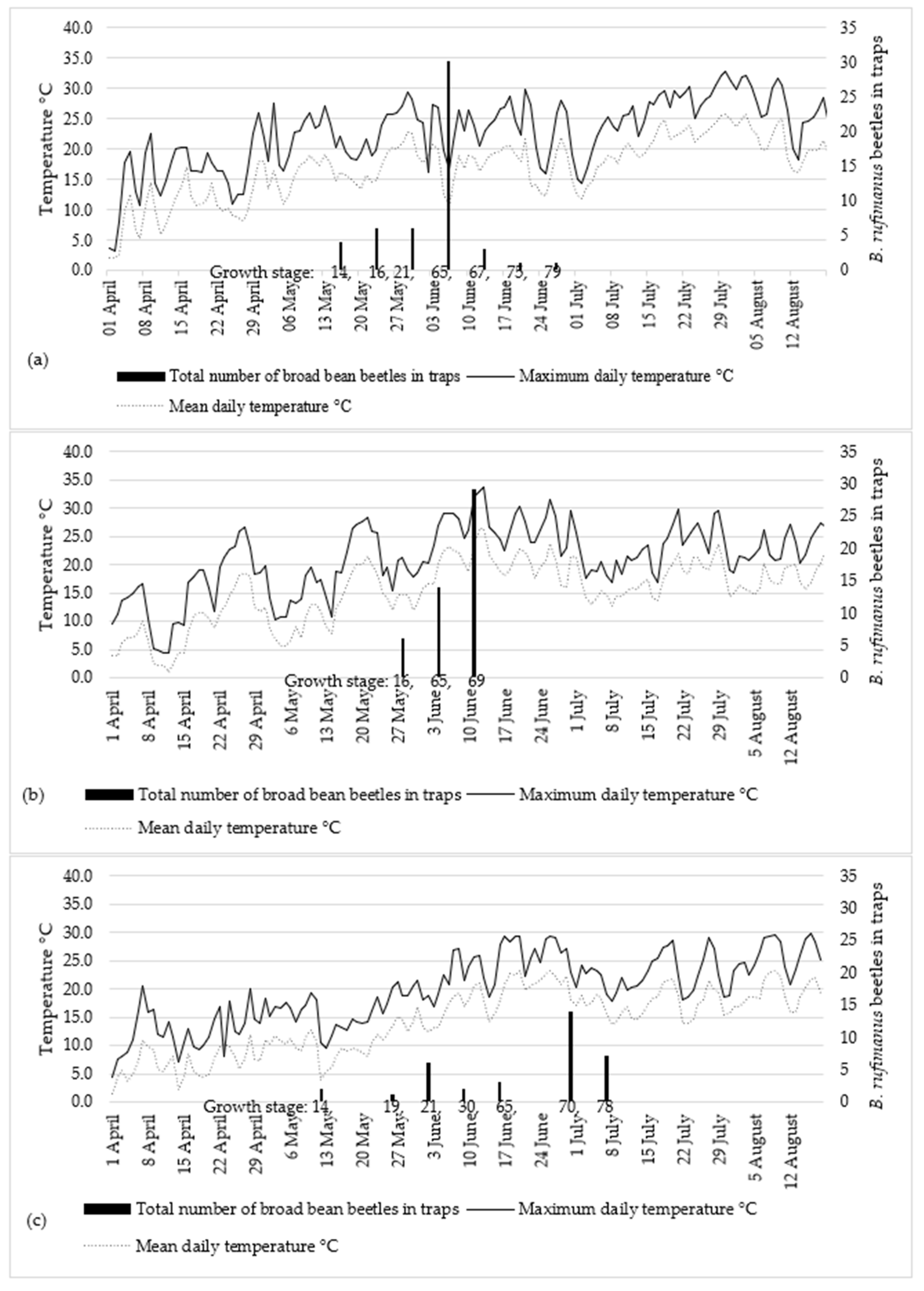
2.1. Seasonal Abundance of Bruchus rufimanus Adults
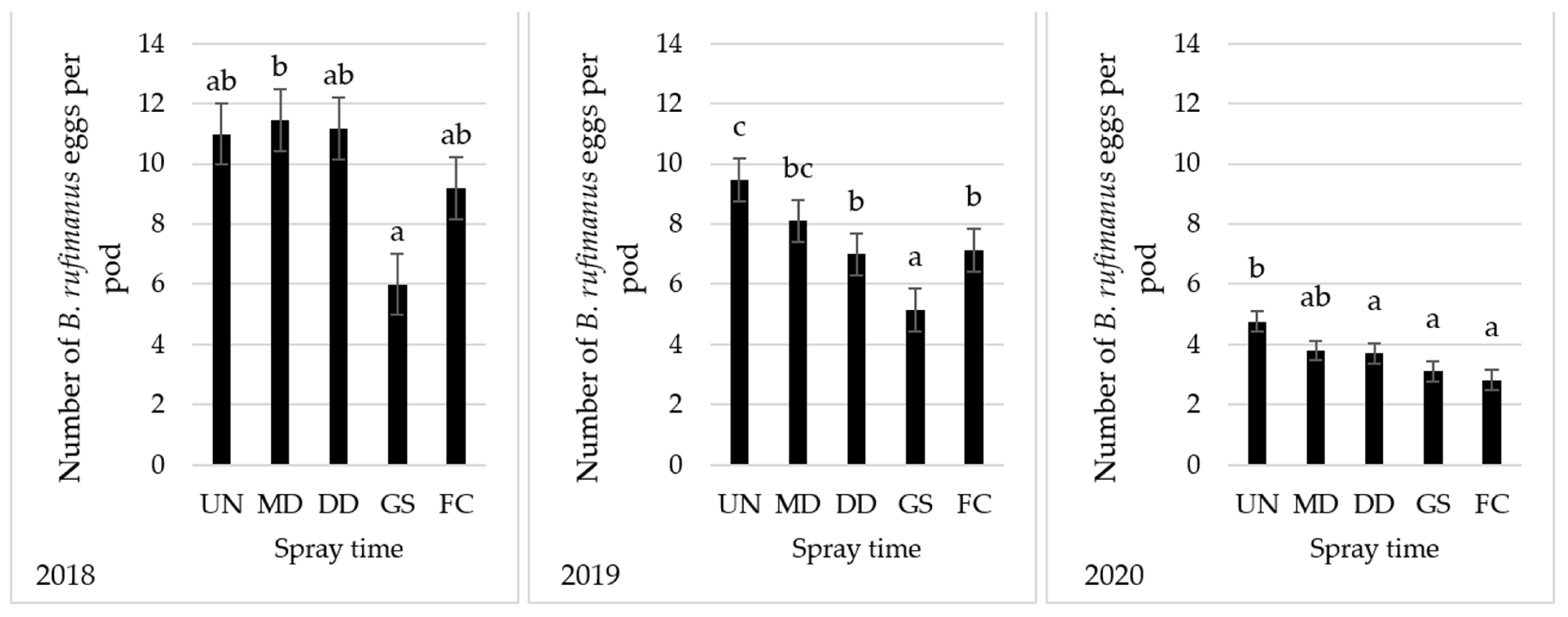
2.2. Bruchus rufimanus Oviposition
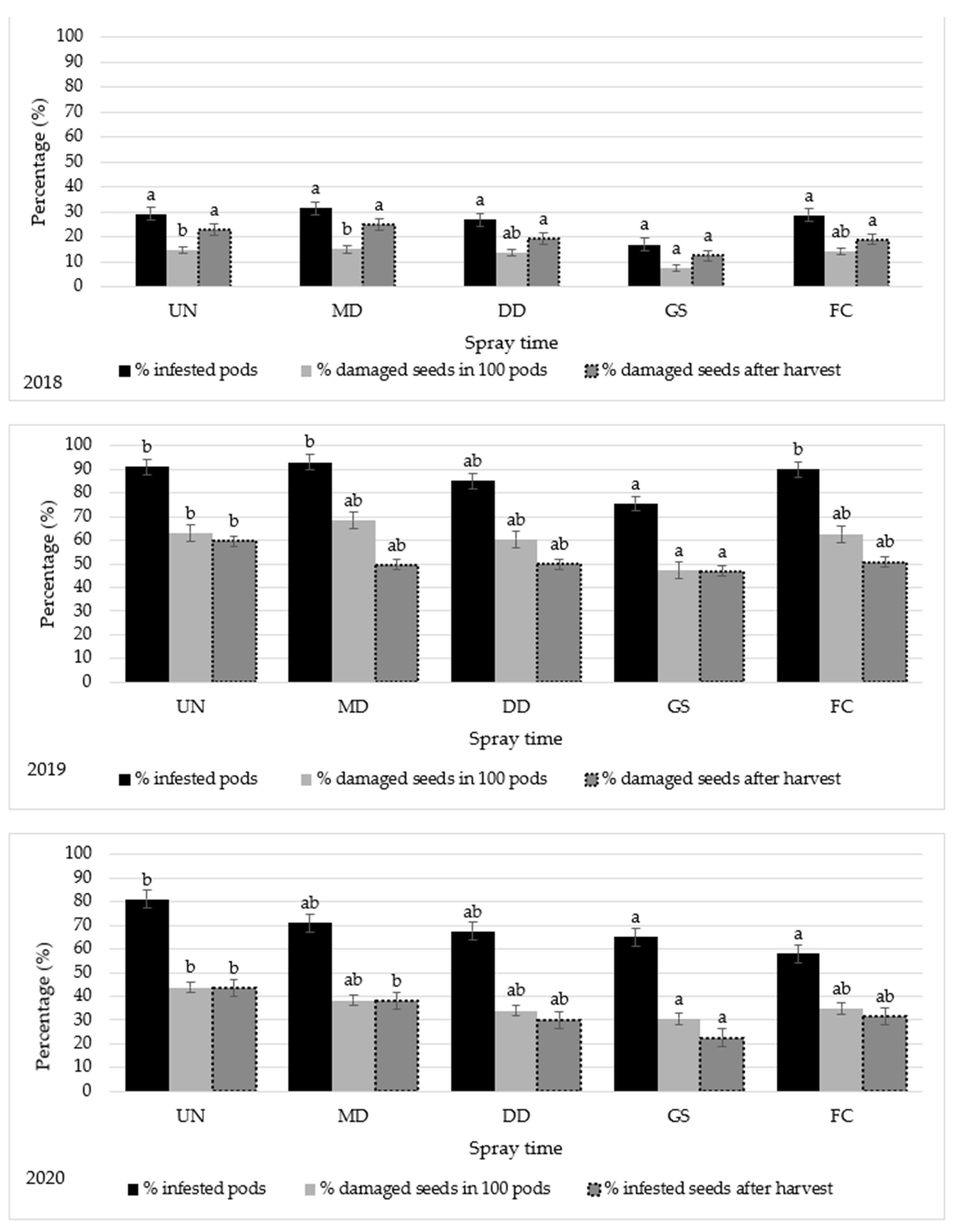
2.3. Bruchus rufimanus Control
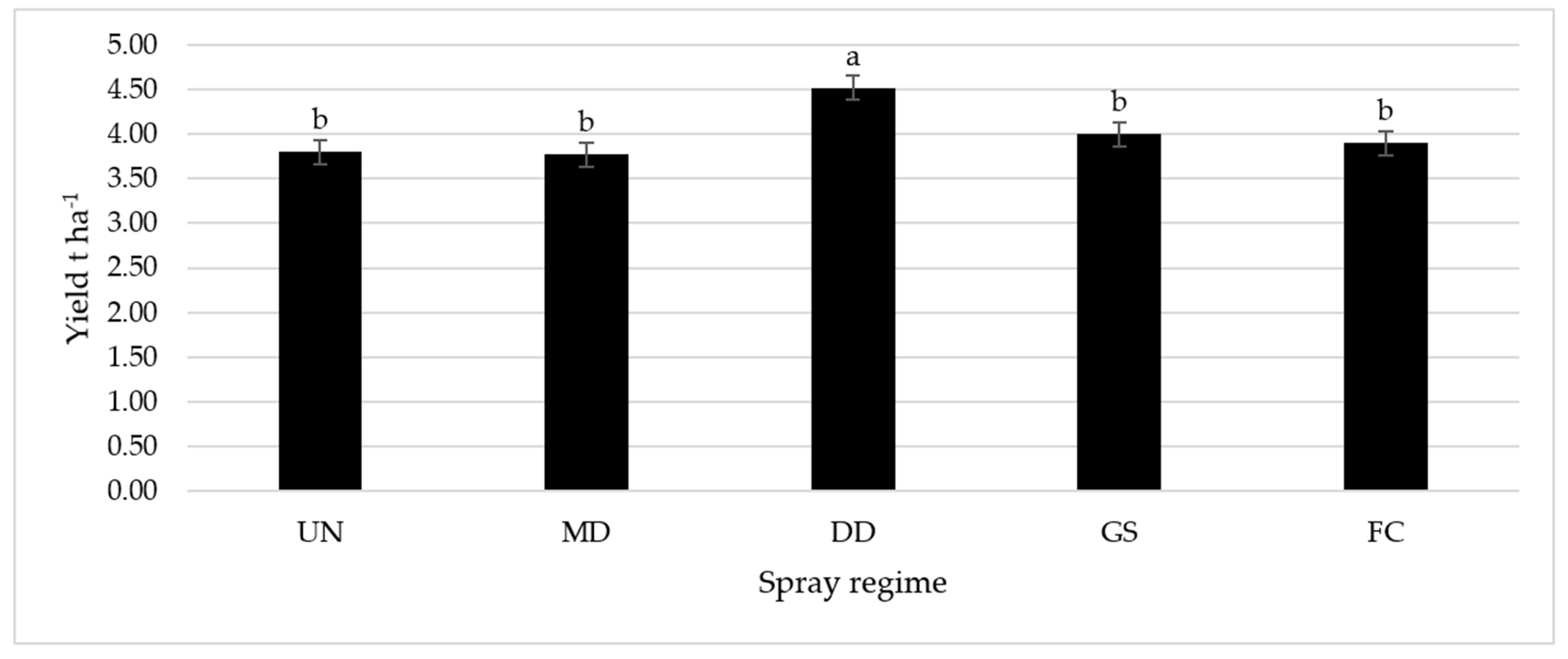
2.4. Yield
3. Discussion
4. Materials and Methods
4.1. Details of the Field Experiment
4.2. Bruchus rufimanus Monitoring, Collection and Identification
4.3. Bruchus rufimanus Control
4.4. Number of Eggs Laid by Bruchus rufimanus on Pods
4.5. Damaged Seed by Bruchus rufimanus Larvae
4.6. Yield
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pratap, A.; Gupta, S. The Beans and the Peas: From Orphan to Mainstream Crops; Woodhead Publishing: Cambridge, UK, 2020; ISBN 0-12-821450-3. [Google Scholar]
- Kusvuran, A.; Parlak, E.L.; Sağlamtiïmur, T. Biomass Yield of Faba Bean (Vicia faba L.) and Its Mixture with Some Grasses (Poaceae). Turk. J. Agric. Nat. Sci. 2015, 2, 178–184. [Google Scholar]
- Hoffmann, D.; Jiang, Q.; Men, A.; Kinkema, M.; Gresshoff, P.M. Nodulation Deficiency Caused by Fast Neutron Mutagenesis of the Model Legume Lotus japonicus. J. Plant Physiol. 2007, 164, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Merga, B.; Egigu, M.C.; Wakgari, M. Reconsidering the Economic and Nutritional Importance of Faba Bean in Ethiopian Context. Cogent Food Agric. 2019, 5, 1683938. [Google Scholar] [CrossRef]
- OSP. Official Statistics Portal. Sown Area of Agricultural Crops. 2021. Available online: http://www.Osp.Stat.Gov.Lt/En. (accessed on 5 January 2022).
- Wang, L.; Lin, K.; Chen, C.; Fu, S.; Xue, F. Diapause Induction and Termination in the Small Brown Planthopper, Laodelphax Striatellus (Hemiptera: Delphacidae). PLoS ONE 2014, 9, e107030. [Google Scholar] [CrossRef] [PubMed]
- Sgolastra, F.; Bosch, J.; Molowny-Horas, R.; Maini, S.; Kemp, W.P. Effect of Temperature Regime on Diapause Intensity in an Adult-Wintering Hymenopteran with Obligate Diapause. J. Insect Physiol. 2010, 56, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Schebeck, M.; Hansen, E.M.; Schopf, A.; Ragland, G.J.; Stauffer, C.; Bentz, B.J. Diapause and Overwintering of Two Spruce Bark Beetle Species. Physiol. Entomol. 2017, 42, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Knell, R.J.; Thackeray, S.J. Voltinism and Resilience to Climate-Induced Phenological Mismatch. Clim. Chang. 2016, 137, 525–539. [Google Scholar] [CrossRef]
- Ozolinèius, R. Possible Effects of Climate Change on Forest Biodiversity, Tree Growth and Condition: Review of Research in Lithuania. Balt. For. 2012, 18, 156–167. [Google Scholar]
- Ward, R.L.; Smart, L. The Effect of Temperature on the Effectiveness of Spray Applications to Control Bean Seed Beetle (Bruchus Rufimanus) in Field Beans (Vicia faba). Asp. Appl. Biol. 2011, 106, 247–254. [Google Scholar]
- Ward, R. The Biology and Ecology of Bruchus Rufimanus (Bean Seed Beetle). Ph.D. Dissertation, Newcastle University, Newcastle, UK, 2018. [Google Scholar]
- Seidenglanz, M.; Huňady, I. Effects of Faba Bean (Vicia faba) Varieties on the Development of Bruchus Rufimanus. Czech J. Genet. Plant Breed. 2016, 52, 22–29. [Google Scholar] [CrossRef]
- Gailis, J.; Astašova, N.; Jākobsone, E.; Ozoliņa-Pole, L. Biology of Broadbean Seed Beetle (Bruchus rufimanus; Coleoptera: Chrysomelidae) in Latvia. Acta Agric. Scand. Sect. Soil Plant Sci. 2021, 72, 4–16. [Google Scholar] [CrossRef]
- Pölitz, B.; Reike, H.-P. Untersuchungen zu Biologie und Befallsdynamik des Ackerbohnenkäfers (Coleoptera, Bruchidae: Bruchus rufimanus) in Sachsen. Gesunde Pflanz. 2019, 71, 79–85. [Google Scholar] [CrossRef]
- Epperlein, K. Investigation of the damage of broad bean weevil Bruchus rufimanus Bohem. (Col., Bruchidae) on broad bean seed (Vicia faba L.). Anz. Fuer. Schaedlingskunde Pflanzenschutz Umweltschutz 1992, 65, 147–150. [Google Scholar] [CrossRef]
- Kaniuczak, Z. Seed Damage of Field Bean (Vicia Faba L. Var. Minor Harz.) Caused by Bean Weevils (Bruchus rufimanus Boh.) (Coleoptera: Bruchidae). J. Plant Prot. Res. 2004, 44, 125–129. [Google Scholar]
- Roubinet, E. Management of the Broad Bean Weevil (Bruchus rufimanus Boh.) in Faba Bean (Vicia faba L.); Swedish University of Agricultural Sciences: Uppsala, Sweden, 2016; p. 23. [Google Scholar]
- Tran, B.; Darquenne, J.; Huignard, J. Changes in Responsiveness to Factors Inducing Diapause Termination in Bruchus rufimanus (Boh.) (Coleoptera: Bruchidae). J. Insect Physiol. 1993, 39, 769–774. [Google Scholar] [CrossRef]
- Tamutis, V.; Tamutė, B.; Ferenca, R. A Catalogue of Lithuanian Beetles (Insecta, Coleoptera). Zookeys 2011, 121, 1–494. [Google Scholar] [CrossRef] [PubMed]
- Gahukar, R.T.; Reddy, G.V.P. Management of Insect Pests in the Production and Storage of Minor Pulses. Ann. Entomol. Soc. Am. 2018, 111, 172–183. [Google Scholar] [CrossRef]
- Mansoor, M.M.; Afzal, M.; Raza, A.B.M.; Akram, Z.; Waqar, A.; Afzal, M.B.S. Post-Exposure Temperature Influence on the Toxicity of Conventional and New Chemistry Insecticides to Green Lacewing Chrysoperla Carnea (Stephens) (Neuroptera: Chrysopidae). Saudi J. Biol. Sci. 2015, 22, 317–321. [Google Scholar] [CrossRef]
- Rusch, A.; Valantin-Morison, M.; Roger-Estrade, J.; Sarthou, J.-P. Local and Landscape Determinants of Pollen Beetle Abundance in Overwintering Habitats. Agric. For. Entomol. 2012, 14, 37–47. [Google Scholar] [CrossRef]
- Hamani-Aoudjit, S.; Medjdoub-Bensaad, F. Monitoring the Activity and Biology of Pest Bruchus Rufimanus (Coleoptera: Chrysomelidae) in Beans Crops. IJARe 2019, 53, 632–635. [Google Scholar] [CrossRef]
- Segers, A.; Caparros Megido, R.; Lognay, G.; Francis, F. Overview of Bruchus Rufimanus Boheman 1833 (Coleoptera: Chrysomelidae): Biology, Chemical Ecology and Semiochemical Opportunities in Integrated Pest Management Programs. Crop Prot. 2021, 140, 105411. [Google Scholar] [CrossRef]
- Medjdoub-Bensaad, F.; Khelil, M.; Huignard, J. Bioecology of Broad Bean Bruchid Bruchus rufimanus Boh. (Coleoptera: Bruchidae) in a Region of Kabylia in Algeria. Afr. J. Agric. Res. 2007, 2, 412–417. [Google Scholar]
- Titouhi, F.; Amri, M.; Jemâa, J.M.B. Status of Coleopteran Insects Infesting Faba Bean in Tunisia with Emphasis on Population Dynamics and Damage of Bruchus rufimanus (Chrysomylidae). Basic Res. J. Agric. Sci. Rev. 2015, 4, 225–233. [Google Scholar]
- Titouhi, F.; Amri, M.; Mediouni-Ben Jemâa, J. Susceptibility of Some Tunisian Broad Bean Varieties to Injuries Caused by the Broad Bean Weevil Bruchus rufimanus in North Tunisia. Tunis. J. Plant Prot. 2015, 10, 55–61. [Google Scholar]
- Gebremedhin, H.; Tadesse, A.; Belay, T. Relating Climatic Factors to Foraging Behavior of Honeybees (Apis mellifera) during Blooming Period of Guizotia abyssinica (L.F.). Livest. Res. Rural. Dev. 2014, 26, 2–7. [Google Scholar]
- Hamidi, R.; Taupin, P.; Frérot, B. Physiological Synchrony of the Broad Bean Weevil, Bruchus rufimanus Boh., to the Host Plant Phenology, Vicia faba L. Front. Insect Sci. 2021, 1, 707323. [Google Scholar] [CrossRef]
- Bruce, T.J.; Martin, J.L.; Smart, L.E.; Pickett, J.A. Development of Semiochemical Attractants for Monitoring Bean Seed Beetle, Bruchus rufimanus. Pest Manag. Sci. 2011, 67, 1303–1308. [Google Scholar] [CrossRef]
- Samir, M.; Karima, K.-G.; Ferroudja, M.-B. Effect of Broad Bean Varieties and Faba Bean upon Populations Dynamic of Bruchus rufimanus (Coleoptera: Chrysomelidae: Bruchinae) in Kabylia Region (Algeria). Int. J. Agric. Sci. Res. 2015, 5, 79–88. [Google Scholar]
- Franssen, C.J.H. De Levenswijze en de Bestrijding van de Tuinboonkever. Land Bouwke Onderz 1956, 62, 1–75. [Google Scholar]
- Hoffmann, A.; Labeyrie, V.; Balachowsky, A.S. Famille des Bruchidae. In Entomologie Appliquée à L’agriculture; Masson et Cie: Paris, France, 1962; pp. 185–188. [Google Scholar]
- Leppik, E.; Pinier, C.; Frerot, B. Chemical Landscape of Agro-Biocoenosis: Case Study of Broad Faba Bean and Its Specialized Pest Bruchus rufimanus. In Proceedings of the 10th International Conference on Agricultual Pests, Montpellier, France, 22–23 October 2014. [Google Scholar]
- Kaniuczak, Z. Bruchus Rufimanus on Beans. Ochr. Roślin 2006, 51, 15–17. [Google Scholar]
- Carrillo-Perdomo, E.; Raffiot, B.; Ollivier, D.; Deulvot, C.; Magnin-Robert, J.-B.; Tayeh, N.; Marget, P. Identification of Novel Sources of Resistance to Seed weevils (Bruchus Spp.) in a Faba Bean Germplasm Collection. Front. Plant Sci. 2019, 9, 1914. [Google Scholar] [CrossRef]
- Aznar-Fernández, T.; Carrillo-Perdomo, E.; Flores, F.; Rubiales, D. Identification and Multi-Environment Validation of Resistance to Pea Weevil (Bruchus pisorum) in Pisum Germplasm. J. Pest Sci. 2018, 91, 505–514. [Google Scholar] [CrossRef]
- Catarino, R.; Ceddia, G.; Areal, F.J.; Park, J. The Impact of Secondary Pests on Bacillus Thuringiensis (Bt) Crops. Plant Biotechnol. J. 2015, 13, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Douglas, M.R.; Tooker, J.F. Meta-Analysis Reveals That Seed-Applied Neonicotinoids and Pyrethroids Have Similar Negative Effects on Abundance of Arthropod Natural Enemies. PeerJ 2016, 4, e2776. [Google Scholar] [CrossRef] [PubMed]
- Hagstrum, D. Atlas of Stored-Product Insects and Mites; Elsevier: Amsterdam, The Netherlands, 2017; Volume 588. [Google Scholar]
- Titouhi, F. Protective Effects of Three Artemisia Essential Oils against Callosobruchus maculatus and Bruchus rufimanus (Coleoptera: Chrysomelidae) and the Extended Side-Effects on Their Natural Enemies. J. Stored Prod. Res. 2017, 72, 11–20. [Google Scholar] [CrossRef]
- Saeidi, K.; Mirfakhraie, S. Trials on the Timing of Chemical Control of Lentil Weevil, Bruchus Lentis Frölich (Coleoptera: Chrysomelidae: Bruchinae) in Lentil Field in Gachsaran Region (Iran). J. Entomol. Acarol. Res. 2017, 49, 3. [Google Scholar] [CrossRef]
- Teferra, Y.K.; Dubale, B.F. Evaluation of the Exact Spraying Time of Chemicals for Management of Bruchus Pisorum Under Field Condition’s. J. Agric. Crops 2019, 5, 109–111. [Google Scholar] [CrossRef]
- Alexoaei, A.; Robu, R.; Cojanu, V.; Miron, D.; Holobiuc, A.-M. Good Practices in Reforming the Common Agricultural Policy to Support the European Green Deal—A Perspective on the Consumption of Pesticides and Fertilizers. Amfiteatru Econ. 2022, 24, 525–545. [Google Scholar] [CrossRef]
- Weber, E.; Bieiholder, H. Erläuterungen Zu Den BBCH-Dezimal-Codes Für Die Entwicklungsstadien von Mais, Raps, Faba-Bohne, Sonnenblume Und Erbse-Mit Abbildungen. Gesunde Pflanz. 1990, 42, 308–321. [Google Scholar]
- Feller, C.; Bleiholder, H.; Buhr, L.; Hack, H.; Heß, M.; Klose, R.; Meier, U.; Stauß, R.; Boom, T.; Weber, E. Phänologische Entwicklungsstadien von GemüsepflanzenII. Fruchtgemüse und Hülsenfrüchte: Codierung und Beschreibung nach der erweiterten BBCH-Skala—Mit Abbildungen. Heft 1995, 47, 217–232. [Google Scholar]
- Kingsolver, J.M. Handbook of the Bruchidae of the United States and Canada (Insecta, Coleoptera); Agricultural Research Service: Washington, DC, USA, 2004; Volume 2, p. 636. [Google Scholar]
- Herms, D. Using Degree-Days and Plant Phenology to Predict Pest Activity. In IPM (Integrated Pest Management) of Midwest Landscapes; Minnesota Agricultural Experiment Station Publication: Saint Paul, MN, USA, 2004; Volume 58, pp. 49–59. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mean Daily Temperatures Observed (°C) | Spearman’s Correlation Coefficients | p Value |
|---|---|---|
| 2 days prior to monitoring | 0.15378 | 0.742 |
| 7 days prior to monitoring | 0.43842 | 0.3251 |
| 14 days prior to monitoring | 0.64908 | 0.1147 |
| 28 days prior to monitoring | 0.78357 | 0.0071 |
| Spray Regime | Year | ||
|---|---|---|---|
| 2018 | 2019 | 2020 | |
| UN | – | – | – |
| MD | BBCH 14—16 May | BBCH 15—23 May | BBCH 16—26 May |
| DD | BBCH 51—28 May | BBCH 65—4 June | BBCH 16—28 May |
| GS | BBCH 67—14 June | BBCH 69—14 June | BBCH 69—21 June |
| FC | BBCH 11—7 May | BBCH 13—17 May | BBCH 10—3 May |
| BBCH 51—28 May | BBCH 15—23 May | BBCH 16—28 May | |
| BBCH 71—21 June | BBCH 70—12 June | BBCH 69—11 June | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almogdad, M.; Semaškienė, R.; Tamošiūnas, K. Optimizing Insecticide Application Timing for Broad Bean Weevil Control and Minimizing Crop Damage in Broad Bean (Vicia faba Linn.). Plants 2023, 12, 1839. https://doi.org/10.3390/plants12091839
Almogdad M, Semaškienė R, Tamošiūnas K. Optimizing Insecticide Application Timing for Broad Bean Weevil Control and Minimizing Crop Damage in Broad Bean (Vicia faba Linn.). Plants. 2023; 12(9):1839. https://doi.org/10.3390/plants12091839
Chicago/Turabian StyleAlmogdad, Mohammad, Roma Semaškienė, and Kęstutis Tamošiūnas. 2023. "Optimizing Insecticide Application Timing for Broad Bean Weevil Control and Minimizing Crop Damage in Broad Bean (Vicia faba Linn.)" Plants 12, no. 9: 1839. https://doi.org/10.3390/plants12091839