
Characterization of Polylepis tarapacana Life Forms in the Highest-Elevation Altiplano in South America: Influence of the Topography, Climate and Human Uses

 , ,
, ,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Results
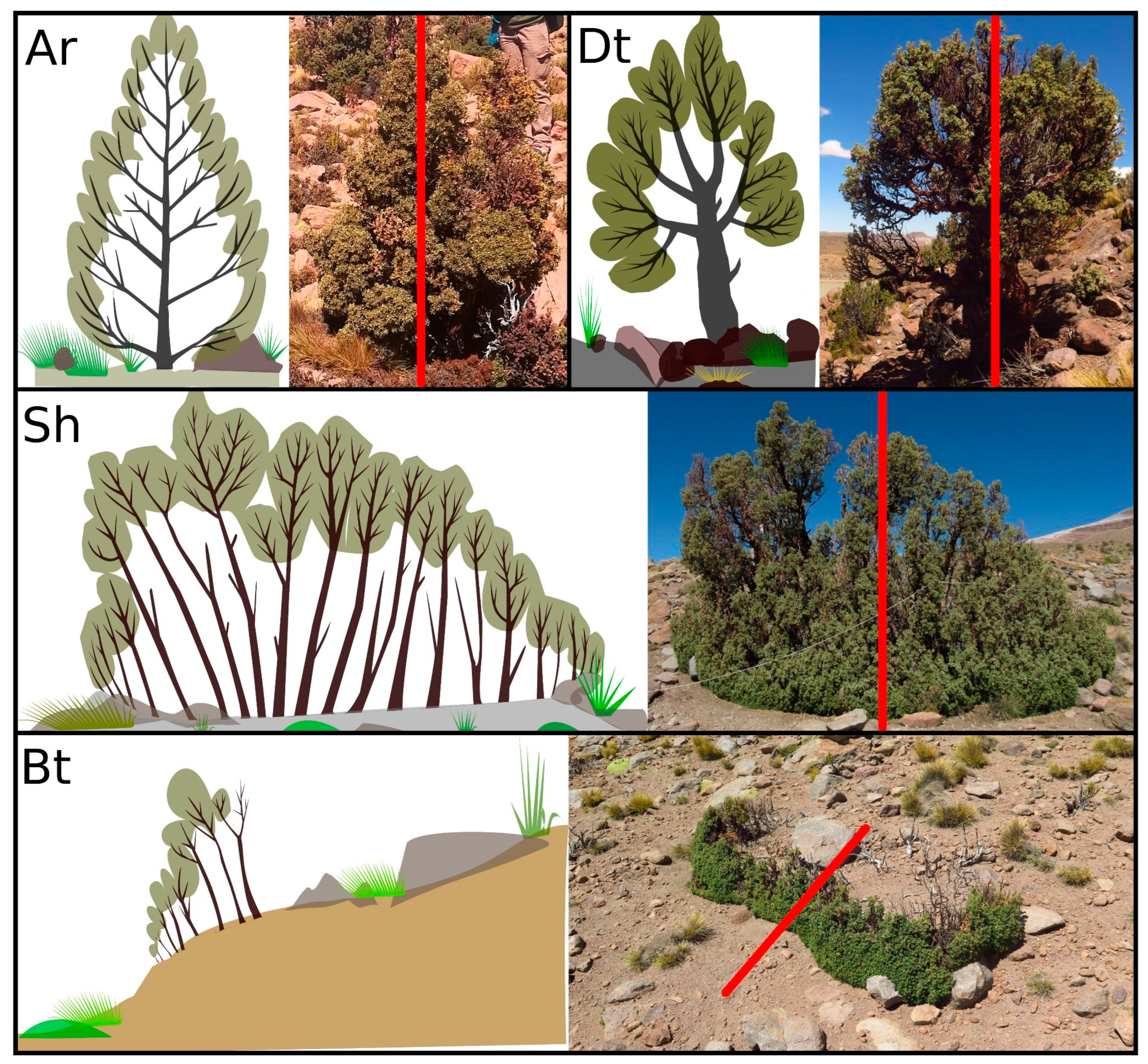
2.1. Life Forms of P. tarapacana
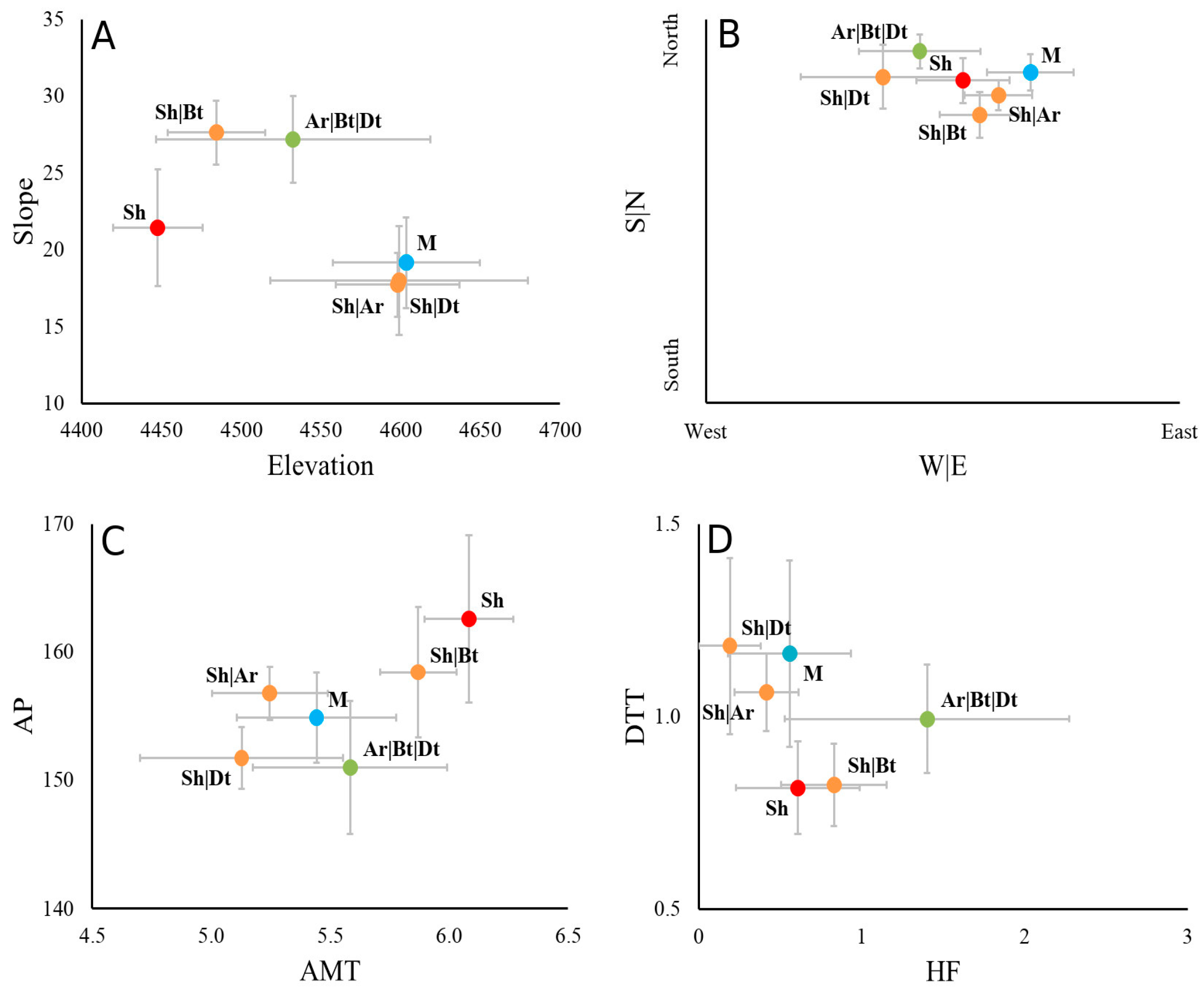
2.2. Changes in the Frequency of Life Forms According to Topographic, Climatic, Geographic and Human Use Factors
3. Discussion
3.1. Frequency of P. tarapacana Life Forms in the Argentinean Altiplano
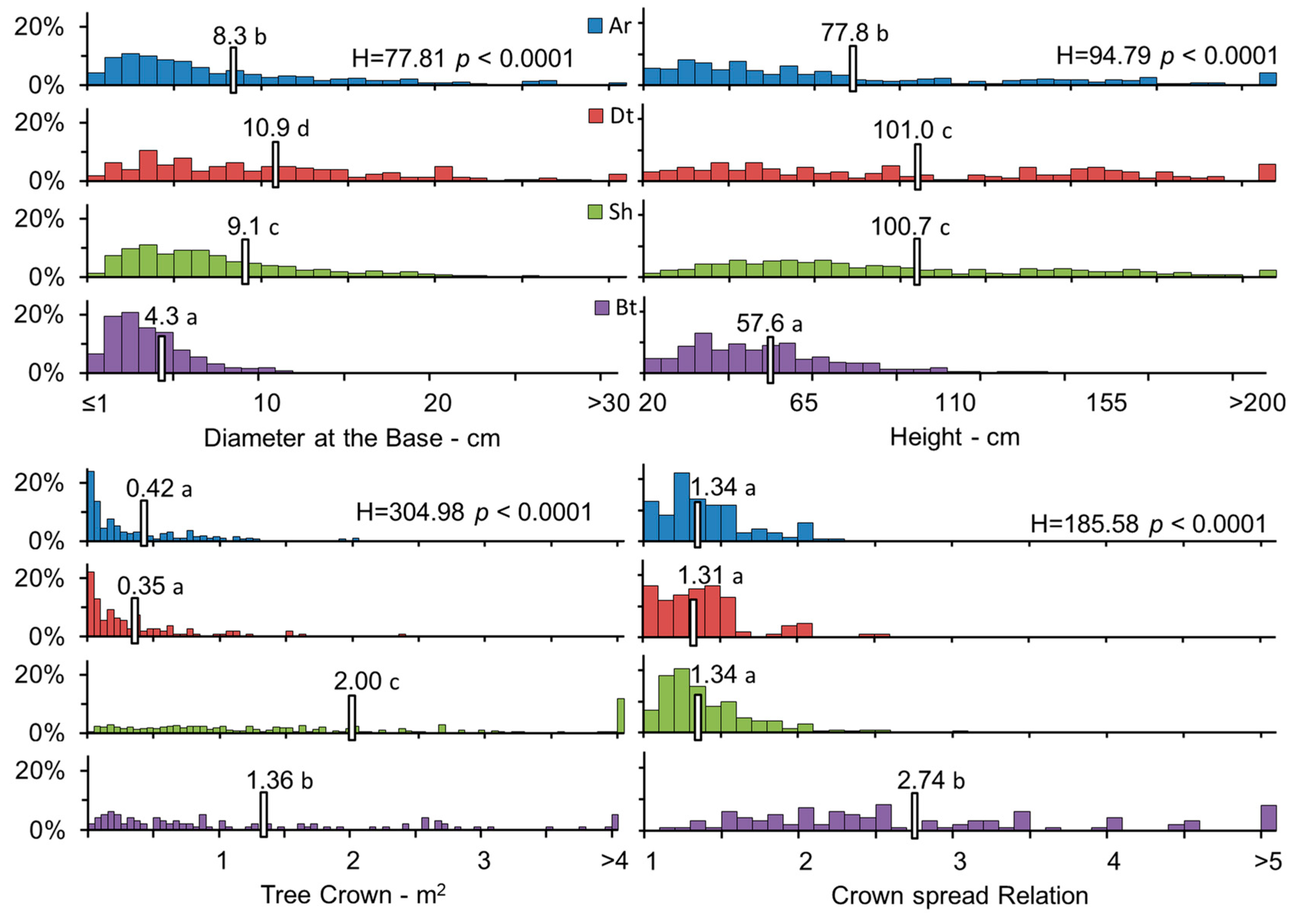
3.2. Biometric Characteristics of Different P. tarapacana Life Forms
3.3. Influence of Topographic, Climatic, Life Zones and Human Use Factors in the Frequency of Life Forms
4. Materials and Methods
4.1. Study Species
4.2. Study Area
4.3. Classification of Life Forms of P. tarapacana
4.4. Data Obtained
4.5. Environmental Characterization
4.6. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| DAB | Diameter at the base of the tree |
| H | Height |
| TC | Tree crown |
| CsR | Crown spread ratio |
| GPS | Global geopositioning device |
| TAMT | Tropical alpine moist tundra |
| TAWT | Tropical alpine wet tundra |
| TSDS | Tropical subalpine dry scrub |
| TSMF | Tropical subalpine moist forest |
| AMT | Annual mean temperature |
| AP | Annual precipitation |
| HF | Human footprint |
| DTT | Distance to town |
| M | Multiple category |
| Ar | Arborescents |
| Dt | Dwarf tree |
| Sh | Shrubs |
| Bt | Brousse tigrée |
| NA | North aspect |
| EA | East aspect |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diameter at Base (cm) | Height (cm) | Tree Crown (m2) | Crown Spread Ratio | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variable | AR | Dt | Sh | Bt | AR | Dt | Sh | Bt | AR | Dt | Sh | Bt | AR | Dt | Sh | Bt | |
| <4400 | 6.3 ± 3.5 | 5.4 ± 3.3 | 6.3 ± 4.0 | 4.1 ± 2.5 | 56.2 ± 20.1 | 59.3 ± 24.7 | 71.9 ± 29.4 | 59.4 ± 23.5 | 0.3 ± 0.4 | 0.1 ± 0.2 | 1.8 ± 2.3 | 1.3 ± 1.4 | 1.4 ± 0.3 | 1.2 ± 0.2 | 1.3 ± 0.3 | 2.5 ± 0.9 | |
| 4400–4500 | 6.7 ± 4.0 | 6.8 ± 6.7 | 5.9 ± 3.8 | 4.1 ± 2.7 | 63.9 ± 26.6 | 57.4 ± 30.5 | 76.8 ± 36.7 | 55.9 ± 18.8 | 0.3 ± 0.3 | 0.1 ± 0.1 | 1.2 ± 2.3 | 1.0 ± 0.5 | 1.3 ± 0.2 | 1.2 ± 0.1 | 1.3 ± 0.3 | 2.2 ± 1.6 | |
| Elevation | 4500–4600 | 8.6 ± 7.3 | 11.1 ± 7.6 | 8.1 ± 5.7 | 3.7 ± 2.2 | 87.4 ± 61.0 | 108.5 ± 61.1 | 99.1 ± 46.5 | 52.9 ± 17.4 | 0.5 ± 0.5 | 0.4 ± 0.4 | 1.3 ± 1.5 | 1.1 ± 0.7 | 1.5 ± 0.4 | 1.3 ± 0.2 | 1.4 ± 0.4 | 2.9 ± 1.3 |
| 4600–4700 | 8.0 ± 6.8 | 11.9 ± 8.1 | 9.7 ± 7.8 | 4.1 ± 2.6 | 72.6 ± 54.9 | 108.2 ± 65.2 | 100.2 ± 62.1 | 53.9 ± 22.0 | 0.5 ± 0.7 | 0.4 ± 0.5 | 1.3 ± 1.3 | 0.9 ± 0.8 | 1.2 ± 0.2 | 1.4 ± 0.4 | 1.3 ± 0.4 | 2.9 ± 0.9 | |
| >4700 | 9.7 ± 8.3 | 12.6 ± 7.6 | 10.8 ± 6.7 | 3.7 ± 3.0 | 92.5 ± 59.0 | 118.6 ± 49.1 | 124.3 ± 54.5 | 53.6 ± 30.0 | 0.5 ± 0.6 | 0.4 ± 0.4 | 1.6 ± 1.5 | 0.9 ± 0.9 | 1.3 ± 0.3 | 1.3 ± 0.3 | 1.3 ± 0.3 | 3.4 ± 1.4 | |
| <15 | 9.1 ± 7.8 | 12.5 ± 8.1 | 10.3 ± 6.7 | 5.0 ± 3.3 | 85.5 ± 59.6 | 118.5 ± 58.4 | 113.1 ± 50.6 | 62.4 ± 23.5 | 0.5 ± 0.6 | 0.4 ± 0.5 | 1.5 ± 1.3 | 1.2 ± 1.4 | 1.4 ± 0.4 | 1.3 ± 0.2 | 1.4 ± 0.4 | 3.3 ± 1.2 | |
| Slope | 15–25 | 8.9 ± 7.3 | 11.5 ± 7.7 | 8.4 ± 5.6 | 4.2 ± 2.7 | 86.4 ± 56.3 | 105.8 ± 56.5 | 101.7 ± 50.7 | 57.0 ± 25.6 | 0.5 ± 0.5 | 0.4 ± 0.4 | 1.7 ± 2.3 | 1.0 ± 0.9 | 1.3 ± 0.3 | 1.3 ± 0.3 | 1.3 ± 0.3 | 2.8 ± 1.5 |
| >25 | 6.4 ± 4.5 | 5.8 ± 3.8 | 5.2 ± 3.3 | 3.5 ± 2.1 | 64.5 ± 37.2 | 60.8 ± 30.3 | 67.0 ± 28.8 | 52.7 ± 18.9 | 0.3 ± 0.3 | 0.1 ± 0.2 | 1.1 ± 1.6 | 1.0 ± 0.7 | 1.4 ± 0.3 | 1.3 ± 0.3 | 1.3 ± 0.3 | 2.5 ± 1.3 | |
| N | N | 9.0 ± 7.3 | 10.4 ± 7.5 | 8.2 ± 6.0 | 4.2 ± 2.7 | 84.4 ± 56.4 | 98.8 ± 54.1 | 93.8 ± 47.9 | 58.8 ± 22.4 | 0.5 ± 0.6 | 0.3 ± 0.4 | 1.5 ± 2.1 | 1.1 ± 0.8 | 1.3 ± 0.3 | 1.3 ± 0.3 | 1.4 ± 0.3 | 2.5 ± 1.4 |
| aspect | Rest | 7.4 ± 6.5 | 11.3 ± 7.9 | 7.5 ± 5.4 | 3.7 ± 2.5 | 75.9 ± 51.2 | 106.3 ± 62.5 | 94.6 ± 50.0 | 51.2 ± 21.4 | 0.4 ± 0.4 | 0.4 ± 0.4 | 1.4 ± 1.4 | 1.0 ± 1.1 | 1.3 ± 0.3 | 1.3 ± 0.3 | 1.3 ± 0.3 | 3.0 ± 1.3 |
| E | W | 9.0 ± 7.0 | 9.9 ± 7.6 | 8.8 ± 5.1 | 4.0 ± 2.2 | 88.6 ± 51.9 | 91.5 ± 55.6 | 103.2 ± 46.8 | 54.0 ± 21.3 | 0.5 ± 0.5 | 0.3 ± 0.3 | 1.5 ± 1.5 | 1.0 ± 0.8 | 1.3 ± 0.3 | 1.1 ± 0.1 | 1.3 ± 0.3 | 1.9 ± 0.4 |
| aspect | Rest | 8.9 ± 7.3 | 10.2 ± 7.4 | 7.5 ± 5.7 | 4.2 ± 2.8 | 82.5 ± 58.6 | 97.7 ± 54.0 | 88.6 ± 45.3 | 58.8 ± 23.2 | 0.5 ± 0.6 | 0.3 ± 0.4 | 1.4 ± 2.2 | 1.1 ± 0.9 | 1.4 ± 0.3 | 1.3 ± 0.3 | 1.4 ± 0.3 | 2.9 ± 1.3 |
| E | 7.4 ± 6.8 | 11.9 ± 8.0 | 8.1 ± 6.2 | 3.6 ± 2.5 | 74.4 ± 50.6 | 112.1 ± 60.3 | 96.6 ± 53.6 | 51.5 ± 20.6 | 0.4 ± 0.4 | 0.4 ± 0.5 | 1.4 ± 1.5 | 0.9 ± 1.1 | 1.3 ± 0.3 | 1.4 ± 0.3 | 1.3 ± 0.3 | 2.7 ± 1.5 | |
| <5 | 9.5 ± 8.2 | 12.6 ± 8.4 | 11.1 ± 7.2 | 4.1 ± 3.2 | 89.2 ± 60.7 | 116.4 ± 57.5 | 120.9 ± 55.4 | 58.9 ± 30.5 | 0.5 ± 0.7 | 0.5 ± 0.4 | 1.6 ± 1.5 | 1.2 ± 1.2 | 1.3 ± 0.3 | 1.3 ± 0.3 | 1.4 ± 0.3 | 3.2 ± 1.3 | |
| AMT | 5–6 | 8.2 ± 6.9 | 9.3 ± 6.8 | 7.3 ± 5.3 | 3.6 ± 2.2 | 81.3 ± 54.5 | 91.3 ± 52.9 | 92.0 ± 46.6 | 51.3 ± 18.2 | 0.4 ± 0.5 | 0.3 ± 0.3 | 1.3 ± 1.9 | 0.9 ± 0.6 | 1.4 ± 0.4 | 1.3 ± 0.3 | 1.3 ± 0.4 | 2.4 ± 1.5 |
| >6 | 7.2 ± 4.8 | 9.8 ± 7.0 | 6.1 ± 4.3 | 3.0 ± 2.8 | 69.5 ± 41.8 | 89.5 ± 57.3 | 78.8 ± 38.5 | 59.9 ± 21.5 | 0.4 ± 0.4 | 0.3 ± 0.3 | 1.5 ± 1.9 | 1.2 ± 1.2 | 1.4 ± 0.3 | 1.2 ± 0.2 | 1.3 ± 0.3 | 2.5 ± 0.9 | |
| AP | <150 | 7.3 ± 6.1 | 11.3 ± 7.5 | 8.6 ± 5.8 | 5.0 ± 3.0 | 73.0 ± 47.5 | 96.6 ± 52.9 | 96.5 ± 44.4 | 64.5 ± 25.4 | 0.4 ± 0.5 | 0.3 ± 0.4 | 1.8 ± 2.3 | 1.3 ± 1.2 | 1.3 ± 0.3 | 1.3 ± 0.3 | 1.4 ± 0.3 | 2.9 ± 1.5 |
| >150 | 9.0 ± 7.5 | 10.4 ± 7.7 | 7.6 ± 5.7 | 3.3 ± 2.1 | 85.8 ± 57.7 | 103.7 ± 59.0 | 92.8 ± 50.9 | 50.0 ± 18.0 | 0.5 ± 0.5 | 0.3 ± 0.4 | 1.2 ± 1.5 | 0.8 ± 0.6 | 1.4 ± 0.4 | 1.3 ± 0.3 | 1.3 ± 0.3 | 2.6 ± 1.1 | |
| TAMT | 10.3 ± 8.7 | 12.7 ± 8.2 | 11.3 ± 7.1 | 5.5 ± 4.0 | 95.7 ± 62.3 | 119.2 ± 51.6 | 128.2 ± 53.4 | 71.0 ± 32.4 | 0.5 ± 0.7 | 0.5 ± 0.4 | 1.8 ± 1.7 | 1.2 ± 1.1 | 1.3 ± 0.3 | 1.3 ± 0.3 | 1.3 ± 0.3 | 3.3 ± 1.1 | |
| Life Zone | TAWT | 9.5 ± 7.8 | 11.8 ± 4.5 | 13.2 ± 5.0 | 2.9 ± 1.2 | 97.8 ± 56.1 | 123.8 ± 42.4 | 157.7 ± 47.4 | 42.5 ± 7.2 | 0.5 ± 0.4 | 0.4 ± 0.2 | 2.3 ± 1.2 | 0.6 ± 0.1 | 1.3 ± 0.3 | 1.3 ± 0.3 | 1.3 ± 0.3 | 3.3 ± 1.1 |
| TSDS | 7.1 ± 5.9 | 9.5 ± 7.6 | 7.0 ± 5.3 | 3.7 ± 2.4 | 69.3 ± 46.9 | 88.0 ± 58.2 | 85.7 ± 44.6 | 53.3 ± 20.6 | 0.4 ± 0.5 | 0.3 ± 0.4 | 1.3 ± 1.9 | 1.0 ± 0.8 | 1.3 ± 0.3 | 1.3 ± 0.3 | 1.3 ± 0.3 | 2.6 ± 1.5 | |
| TSMF | 8.5 ± 6.5 | 10.6 ± 7.6 | 7.9 ± 5.3 | 5.0 ± 2.3 | 82.5 ± 53.7 | 101.7 ± 56.4 | 90.1 ± 43.2 | 64.4 ± 22.7 | 0.4 ± 0.5 | 0.4 ± 0.4 | 1.5 ± 1.8 | 1.4 ± 1.6 | 1.4 ± 0.4 | 1.3 ± 0.2 | 1.3 ± 0.3 | 2.9 ± 0.9 | |
| HF | 0 | 8.4 ± 6.8 | 11.3 ± 7.4 | 8.5 ± 6.0 | 4.2 ± 2.8 | 80.3 ± 52.0 | 104.5 ± 54.9 | 98.6 ± 49.5 | 57.0 ± 23.6 | 0.4 ± 0.5 | 0.4 ± 0.4 | 1.6 ± 2.0 | 1.1 ± 1.1 | 1.3 ± 0.3 | 1.3 ± 0.3 | 1.3 ± 0.3 | 2.9 ± 1.4 |
| >0 | 8.4 ± 8.1 | 8.4 ± 8.3 | 6.1 ± 4.5 | 3.4 ± 2.0 | 85.8 ± 64.9 | 88.3 ± 62.9 | 79.5 ± 43.3 | 52.3 ± 18.7 | 0.5 ± 0.6 | 0.3 ± 0.4 | 0.8 ± 0.7 | 0.9 ± 0.4 | 1.3 ± 0.2 | 1.1 ± 0.0 | 1.3 ± 0.3 | 1.8 ± 0.3 | |
| <5 | 5.4 ± 3.7 | 4.8 ± 3.4 | 5.6 ± 3.9 | 3.4 ± 2.5 | 51.1 ± 20.8 | 49.4 ± 26.2 | 68.0 ± 30.9 | 50.5 ± 22.3 | 0.3 ± 0.4 | 0.1 ± 0.2 | 1.9 ± 2.2 | 1.3 ± 1.7 | 1.4 ± 0.3 | 1.1 ± 0.2 | 1.4 ± 0.3 | 2.5 ± 1.1 | |
| DTT | 5–10 | 9.0 ± 7.4 | 10.8 ± 6.8 | 7.6 ± 5.4 | 3.6 ± 2.1 | 87.3 ± 57.9 | 107.4 ± 59.0 | 96.1 ± 48.3 | 52.3 ± 18.6 | 0.5 ± 0.6 | 0.3 ± 0.4 | 1.4 ± 2.0 | 0.9 ± 0.6 | 1.3 ± 0.3 | 1.3 ± 0.3 | 1.3 ± 0.4 | 2.8 ± 1.0 |
| >10 | 8.3 ± 7.1 | 11.6 ± 8.5 | 9.6 ± 6.4 | 5.2 ± 3.0 | 80.2 ± 53.6 | 103.3 ± 54.3 | 103.5 ± 51.9 | 65.6 ± 25.1 | 0.4 ± 0.5 | 0.4 ± 0.4 | 1.3 ± 1.3 | 1.1 ± 0.9 | 1.3 ± 0.3 | 1.3 ± 0.3 | 1.3 ± 0.3 | 3.0 ± 1.7 | |
References
- Körner, C. Scaling from species to vegetation: The usefulness of functional groups. In Biodiversity and Ecosystem Function; Schulze, E.D., Mooney, H.A., Eds.; Springer: Berlin, Germany, 1994; pp. 117–140. [Google Scholar]
- Odland, A. Interpretation of altitudinal gradients in South Central Norway based on vascular plants as environmental indicators. Ecol. Indic. 2009, 9, 409–421. [Google Scholar] [CrossRef]
- Hadar, L.; Noy-Meir, I.; Perevolotsky, A. The effect of shrub clearing and grazing on the composition of a Mediterranean plant community: Functional group versus species. J. Veg. Sci. 1999, 10, 673–682. [Google Scholar] [CrossRef]
- Pavón, N.P.; Hernández-Trejo, H.; Rico-Gray, V. Distribution of plant life forms along an altitudinal gradient in the semi arid valley of Zapotitlán, México. J. Veg. Sci. 2000, 11, 39–42. [Google Scholar] [CrossRef]
- Mueller-Dombois, D.; Ellenberg, H. Aims and Methods of Vegetation Ecology; John Wiley & Sons Inc.: New York, NY, USA, 1974; p. 547. [Google Scholar]
- Solbrig, O.T. Plant traits and adaptive strategies: Their role in ecosystem function. In Biodiversity and Ecosystem Function; Schulze, E.D., Mooney, H.A., Eds.; Springer: Berlin/Heidelberg, Germany, 1993; pp. 97–116. [Google Scholar]
- Klimes, L. Life-forms and clonality of vascular plants along an altitudinal gradient in E Ladakh (NW Himalayas). Basic Appl. Ecol. 2003, 4, 317–328. [Google Scholar] [CrossRef]
- Körner, C. Climatic treelines: Conventions, global patterns, causes (Klimatische Baumgrenzen: Konventionen, globale Muster, Ursachen). Erdkunde 2007, 61, 316–324. [Google Scholar] [CrossRef]
- Arroyo, M.T.K.; Cavieres, L.A. High-Elevation Andean Ecosystems. In Encyclopedia of Biodiversity, 2nd ed.; Levin, S.A., Ed.; Academic Press: Cambridge, MA, USA, 2013; pp. 96–110. [Google Scholar] [CrossRef]
- Jaramillo Aguilar, D.A. Fotosíntesis en los Bosques a Mayor Elevación en el Planeta: Polylepis tarapacana en un Gradiente de Elevación en los Andes de Arica y Parinacota, Chile. Ph.D. Thesis, Universidad Austral de Chile, Valdivia, Chile, 2015. [Google Scholar]
- Cowling, R.M.; Esler, K.J.; Midgley, G.F.; Honing, M.A. Plant functional diversity, species diversity and climate in arid and semi-arid southern Africa. J. Arid Environ. 1994, 27, 141–158. [Google Scholar] [CrossRef]
- Huston, M.A. Biological Diversity: The Coexistence of Species; Cambridge University Press: Cambridge, UK, 1994; p. 681. [Google Scholar]
- Ramsay, P.M.; Oxley, E.R.B. The Growth Form Composition of Plant Communities in the Ecuadorian Páramos. Plant Ecol. 1997, 131, 173–192. [Google Scholar] [CrossRef]
- Diaz, S.; Cabido, M. Plant functional types and ecosystem function in relation to global change. J. Veg. Sci. 1997, 8, 463–473. [Google Scholar] [CrossRef]
- Mahdavi, P.; Akhani, H.; Van der Maarel, E. Species Diversity and Life-Form Patterns in Steppe Vegetation along a 3000 m Altitudinal Gradient in the Alborz Mountains, Iran. Folia Geobot. 2013, 48, 7–22. [Google Scholar] [CrossRef]
- Leuschner, C. Are high elevations in tropical mountains arid environments for plants? Ecology 2000, 81, 1425–1436. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life: Functional Ecology of High Mountain Ecosystems, 2nd ed.; Springer: Berlin, Germany, 2003; p. 349. [Google Scholar]
- Kessler, M.; Schmidt-Lebuhn, A.N. Taxonomical and distributional notes on Polylepis (Rosaceae). Org. Divers. Evol. 2006, 6, 67–70. [Google Scholar] [CrossRef]
- Saavedra, J. Caracterización de la Estructura Poblacional de Polylepis tarapacana en Sector Cerro Chiguana, Cuenca del Salar de Surire, Para Establecer Los Lineamientos de un Programa de Restauración Ecológica. Ph.D. Thesis, Universidad de Tarapacá, Arica, Chile, 2013. [Google Scholar]
- López, V.L.; Cellini, J.M. Plantas medicinales asociadas a bosques de Polylepis tarapacana del altiplano jujeño: Riqueza y cobertura en gradientes geográficos, topográficos y de estructura forestal. Ecol. Austral. 2022, 32, 894–907. [Google Scholar] [CrossRef]
- Hoch, G.; Körner, C. Growth, demography and carbon relations of Polylepis trees at the world\u2019s highest treeline. Funct. Ecol. 2005, 19, 941–951. [Google Scholar] [CrossRef]
- Donoso, C. Bosques Templados de Chile y Argentina: Variación, Estructura y Dinámica; Editorial Universitaria: Santiago, Chile, 1993; p. 483. [Google Scholar]
- López, V.L.; Huertas Herrera, A.; Rosas, J.M.; Cellini, J.M. Optimal environmental drivers of high-mountains forest: Polylepis tarapacana cover evaluation in their southernmost distribution range of the Andes. Trees For. People 2022, 9, 100321. [Google Scholar] [CrossRef]
- Ríos, S. Estudio de la Distribución y Caracterización Ecológica de las Poblaciones Locales de Queñoa de Altura (Polylepis tarapacana Phil.) en el Sector de la Provincia de Iquique. Ph.D. Thesis, Universidad de Chile, Tarapacá, Chile, 1998. [Google Scholar]
- De Laubenfels, D.J. Mapping the World’s Vegetation. Regionalization of Formations and Flora, 1st ed.; Syracuse University Press: New York, NY, USA, 1975; p. 246. [Google Scholar]
- Box, E.O. Macroclimate and Plant Forms: An Introduction to Predictive Modeling in Phytogeography; Tasks for Vegetation Science; Antiquariaat Junk: The Hague, The Netherlands, 1981; Volume 1, p. 58. [Google Scholar]
- Rundel, P.W. Shrub life forms. In Response of Plants to Multiple Stress; Mooney, H.A., Winner, W.E., Pell, E.J., Eds.; Academic Press: San Diego, CA, USA, 1991; pp. 345–371. [Google Scholar]
- Wilson, B.F. Shrub stems: Form and function. In Plant Stems: Physiology and Functional Morphology; Gartner, B.L., Ed.; Academic Press: New York, NY, USA, 1995; pp. 91–102. [Google Scholar]
- Valentin, C.; d’Herbès, J.M. Niger tiger bush as a natural water harvesting system. Catena 1999, 37, 231–256. [Google Scholar] [CrossRef]
- Rietkerk, M.; Boerlijst, M.C.; van Langevelde, F.; Hille-Ris-Lambers, R.; van de Koppel, J.; Kumar, L.; Prins, H.H.T.; de Roos, A.M. Self-organization of vegetation in arid ecosystems. Am. Nat. 2002, 160, 524–530. [Google Scholar] [CrossRef]
- Ludwig, J.A.; Wilcox, B.P.; Breshears, D.D.; Tongway, D.J.; Imeson, A.C. Vegetation patches and runoff–erosion as interacting ecohydrological processes in semiarid landscapes. Ecology 2005, 86, 288–297. [Google Scholar] [CrossRef]
- Lefever, R.; Barbier, N.; Couteron, P.; Lejeune, O. Deeply gapped vegetation patterns: On crown/root allometry, criticality and desertification. J. Theor. Biol. 2009, 261, 194–209. [Google Scholar] [CrossRef]
- Renison, D.; Morales, L.; Cuyckens, G.A.E.; Sevillano, C.S.; Cabrera Amaya, D.M. Ecología y conservación de los bosques y arbustales de Polylepis: ¿qué sabemos y qué ignoramos? Ecol. Austral 2018, 28, 163–174. [Google Scholar] [CrossRef]
- IUCN. Red List of Threatened Species. Version 2020-1. 2020. Available online: https://www.iucnredlist.org (accessed on 5 March 2023).
- Renison, D.; Cuyckens, G.A.E.; Pacheco, S.; Guzmán, G.F.; Grau, H.R.; Marcora, P.; Robledo, G.; Cingolani, A.M.; Dominguez, J.; Landi, M.; et al. Distribución y estado de conservación de las poblaciones de árboles y arbustos del género Polylepis (Rosaceae) en las montañas de Argentina. Ecol. Austral. 2013, 23, 27–36. [Google Scholar] [CrossRef]
- Montana, C.; Valientebanuet, A. Floristic and life-form diversity along an altitudinal gradient in an intertropical semiarid Mexican region. Southwest Nat. 1998, 43, 25–39. [Google Scholar]
- Mooney, H.A. Plant forms in relation to environment. In Handbook of Vegetation Science; Strain, B.R., Billings, W.D., Eds.; Vegetation and Environment: The Hague, The Netherlands, 1974; Volume 6, pp. 111–122. [Google Scholar]
- Givnish, T.J. Plant stems: Biomechanical adaptation for energy capture and influence on species distributions. In Plant Stems: Physiology and Functional Morphology; Gartner, B.L., Ed.; Academic Press: San Diego, CA, USA, 1995; pp. 3–49. [Google Scholar]
- Sheffer, M.; Vergnon, R.; Cornelissen, H.C.; Hantson, S.; Holmgren, M.; van Nes, E.H.; Xu, C. Why trees and shrubs but rarely trubs? Trends Ecol. Evol. 2014, 29, 433–434. [Google Scholar] [CrossRef] [PubMed]
- Raunkiær, C. Types biologiques pour la géographie botanique. Overs. Kongel. Dan. Vidensk. Selsk. Forh. Medl. Arb. 1905, 5, 347–437. [Google Scholar]
- Körner, C. A re-assessment of high-elevation treeline positions and their explanation. Oecologia 1998, 115, 445–459. [Google Scholar] [CrossRef]
- Wilson, C.; Grace, J.; Allen, S.; Slack, F. Temperature and stature: A study of temperatures in montane vegetation. Funct. Ecol. 1987, 1, 405–413. [Google Scholar] [CrossRef]
- Pereg, D.; Payette, S. Development of black spruce growth forms at treeline. Plant Ecol. 1998, 138, 137–147. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Kempf, J.S. Branching patterns in forest shrubs and understory trees in relation to habitat. New Phytol. 1980, 86, 219–228. [Google Scholar] [CrossRef]
- Givnish, T.J. Leaf and canopy adaptations in tropical forests. In Physiological Ecology of Plants in the Wet Tropics; Tasks for Vegetation Science; Antiquariaat Junk: The Hague, The Netherlands, 1984; pp. 51–84. [Google Scholar]
- Küppers, M. Ecological significance of above-ground architectural patterns in woody plants: A question of cost-benefit relationships. Trends Ecol. Evol. 1989, 4, 375–379. [Google Scholar] [CrossRef]
- Sun, F.; Kuang, Y.; Wen, D.; Xu, Z.; Li, J.; Zuo, W.; Hou, E. Long-term tree growth rate, water use efficiency, and tree ring nitrogen isotope composition of Pinus massoniana L. in response to global climate change and local nitrogen deposition in Southern China. J. Soils Sediments 2010, 10, 1453–1465. [Google Scholar] [CrossRef]
- Grace, J. Plant response to wind. Agric. Ecosyst. Environ. 1988, 22–23, 71–88. [Google Scholar] [CrossRef]
- Hallinger, M.; Manthey, M.; Wilmking, M. Establishing a missing link: Warm summers and winter snow cover promote shrub expansion into alpine tundra in Scandinavia. New Phytol. 2010, 186, 890–899. [Google Scholar] [CrossRef]
- Neuner, G. Frost resistance in alpine woody plants. Front. Plant Sci. 2014, 5, 654. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.G.; Yoder, B.J. Hydraulic Limits to Tree Height and Tree Growth. BioScience 1997, 47, 235–242. [Google Scholar] [CrossRef]
- Götmark, F.; Götmark, E.; Jensen, A.M. Why Be a Shrub? A Basic Model and Hypotheses for the Adaptive Values of a Common Growth Form. Front. Plant Sci. 2016, 7, 1095. [Google Scholar] [CrossRef]
- Tyree, M.; Sperry, J. Vulnerability of xylem to cavitation and embolism. Ann. Rev. Plant Biol. 1989, 40, 19–36. [Google Scholar] [CrossRef]
- LaMarche, V.C. Environment in Relation to Age of Bristlecone Pines. Ecology 1969, 50, 53–59. [Google Scholar] [CrossRef]
- Schauer, A.J.; Schoettle, A.W.; Boyce, R.L. Partial cambial mortality in high-elevation Pinus aristata (Pinaceae). J. Bot. 2001, 88, 646–652. [Google Scholar] [CrossRef]
- Beuselinck, L.; Steegen, A.; Govers, G.; Nachtergaele, J.; Takken, I.; Poesen, J. Characteristics of sediment deposits formed by intense rainfall events in small catchments in the Belgian Loam Belt. Geomorphology 2000, 32, 69–82. [Google Scholar] [CrossRef]
- Pan, C.; Ma, L.; Shangguan, Z.; Ding, A. Determining the sediment trapping capacity of grass filter strips. J. Hydrol. 2011, 405, 209–216. [Google Scholar] [CrossRef]
- Gallart, F.; Puigdefa’bregas, J.; del Barrio, G. Computer simulation of high mountain terracettes as interaction between vegetation growth and sediment movement. Catena 1993, 20, 529–542. [Google Scholar] [CrossRef]
- Germino, M.J. Plants in Alpine Environments. In Ecology and the Environment; Monson, R., Ed.; Springer: New York, NY, USA, 2014; pp. 1–30. [Google Scholar] [CrossRef]
- Baruch, Z. Patterns of energy content in plants from the Venezuelan pa\u0301ramos. Oecologia 1982, 55, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Stevens, G.C. The elevational gradient in altitudinal range: An extension of Rapoport’s latitudinal rule to altitude. Am. Nat. 1992, 140, 893–911. [Google Scholar] [CrossRef]
- Heitz, P.; Heitz-Seifert, U. Intra and interspecific relations within an epiphyte community in a Mexican humid montane forest. Selbyana 1995, 16, 135–140. [Google Scholar]
- Austin, M.P.; Cunningham, R.B.; Fleming, P.M. New approaches to direct gradient analysis using environmental scalars and statistical curve-fitting procedures. Vegetatio 1984, 55, 11–27. [Google Scholar] [CrossRef]
- Holten, J.J. Vascular plant species richness in relation to altitudinal and slope gradients in the mountain landscape of central Norway. In The Impacts of Climate Variability on Forests; Beniston, M., Innes, J.L., Eds.; Springer: Berlin, Germany, 1998; pp. 231–239. [Google Scholar]
- Macek, P.; Macková, J.; de Bello, F. Morphological and ecophysiological traits shaping altitudinal distribution of three Polylepis treeline species in the dry tropical Andes. Acta Oecol. 2009, 35, 778–785. [Google Scholar] [CrossRef]
- García-Plazaola, J.I.; Rojas, R.; Christie, D.A.; Coopman, R.E. Photosynthetic responses of trees in high-elevation forests: Comparing evergreen species along an elevation gradient in the Central Andes. AoB Plants 2015, 7, plv058. [Google Scholar] [CrossRef] [PubMed]
- Derguy, M.R.; Frangi, J.L.; Drozd, A.A.; Arturi, M.F.; Martinuzzi, S. Holdridge Life Zone Map Republic of Argentina. Gen. Tech. Rep. Dep. Agric. 2019, 51, 1–48. [Google Scholar]
- Luebert, F.; Gajardo, R. Vegetacion alto andina de Parinacota (norte de Chile) y una sinopsis de la vegetacion de la Puna meridional. Phytocoenologia 2005, 35, 79–128. [Google Scholar] [CrossRef]
- López, V.L.; Cellini, J.M.; Cuyckens, G.A.E. Influencia del micrositio y el ambiente en la instalación de Polylepis tarapacana en los Altos Andes. Neotrop. Biodivers. 2021, 7, 135–145. [Google Scholar] [CrossRef]
- Lizárraga, L.; Monguillot, J. Huella Humana en Argentina. Dirección Regional Noroeste, Administración de Parques Nacionales. Salta, Argentina. 2018. Available online: http://geoportal.idesa.gob.ar/layers/geonode%3Ah_humana_v01 (accessed on 15 March 2023).
- Peng, Y.; Lachmuth, S.; Gallegos, S.C.; Kessler, M.; Ramsay, P.M.; Renison, D.; Suarez, R.; Hensen, I. Pleistocene climatic oscillations rather than recent human disturbance influence genetic diversity in one of the world’s highest treeline species. Am. J. Bot. 2015, 102, 1676–1684. [Google Scholar] [CrossRef]
- Cuyckens, G.; Christie, D.; Domic, A.; Malizia, L.; Renison, D. Climate change and the distribution and conservation of the world’s highest elevation woodlands in the South American Altiplano. Glob. Planet. Chang. 2015, 137, 79–87. [Google Scholar] [CrossRef]
- Boza Espinoza, T.E.; Kessler, M. A monograph of the genus Polylepis (Rosaceae). PhytoKeys 2022, 203, 1–274. [Google Scholar] [CrossRef]
- Rojas, R.; Flexas, J.; Coopman, R.E. Particularities of the highest elevation treeline in the world: Polylepis tarapacana Phil. as a model to study ecophysiological adaptations to extreme environments. Flora 2022, 292, 152076. [Google Scholar] [CrossRef]
- Araya-Presa, J.; Squeo, F.A.; Barrientos, L.; Belmonte, E.; Mamani, M.; Arancio, G. Manual de Plantas y Canciones Aymará. Proyecto ExploraCONYCIT ED7/027085: Etnobotánica y Etnomusicología Aymará: Divulgación de La Sabiduría Ancestral Sobre Las Plantas Nativas Del Altiplano de Arica. Universidad de La Serena. Chile. 2003. Available online: http://www.biouls.cl/aymara (accessed on 3 March 2023).
- Garreaud, R.; Vuille, M.; Clement, A.C. The climate of the Altiplano: Observed current conditions and mechanisms of past changes. Palaeogeogr. Palaeoclim. Palaeoecol. 2003, 194, 5–22. [Google Scholar] [CrossRef]
- Morello, J.; Matteucci, S.D.; Rodríguez, A.F.; Silva, M.; De Haron., J.C. Ecorregiones y Complejos Ecosistémicos Argentinos, 1st ed.; Orientación Gráfica Editora: Buenos Aires, Argentina, 2012; p. 773. [Google Scholar]
- Cabrera, A.L. Ecología vegetal de la Puna. Colloq. Geogr. 1968, 9, 91–116. [Google Scholar]
- Oyarzabal, M.; Clavijo, J.; Oakley, L.; Biganzoli, F.; Tognetti, P.; Barberis, I.; Maturo, H.M.; Aragón, R.; Campanello, P.I.; Prado, D.; et al. Unidades de vegetación de la Argentina. Ecol. Austral 2018, 28, 40–63. [Google Scholar] [CrossRef]
- Jenness, J.S. Calculating landscape surface area from digital elevation models. Wildl. Soc. Bull. 2004, 32, 829–839. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1 km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]





| Variable | Range | n | Ar | Dt | Sh | Bt |
|---|---|---|---|---|---|---|
| Elevation | <4400 | 14 | 17 a | 5 a | 59 b | 19 b |
| 4400–4500 | 17 | 17 a | 8 ab | 51 ab | 24 b | |
| 4500–4600 | 19 | 26 ab | 10 abc | 53 ab | 11 a | |
| 4600–4700 | 6 | 30 b | 16 c | 39 a | 14 ab | |
| >4700 | 14 | 35 b | 13 bc | 40 a | 12 a | |
| p | 0.0082 | 0.0349 | 0.0123 | 0.0017 | ||
| HTest | 13.7 | 10.2 | 12.7 | 17.1 | ||
| Slope | <15 | 18 | 30 b | 11 | 55 | 8 a |
| 15–25 | 25 | 27 ab | 11 | 48 | 14 a | |
| >25 | 27 | 18 a | 7 | 50 | 24 b | |
| p | 0.0268 | 0.2320 | 0.5501 | 0.0002 | ||
| HTest | 7.2 | 2.8 | 1.2 | 17.2 | ||
| NA | N | 27 | 22 | 8 | 52 | 18 |
| Rest | 43 | 25 | 10 | 49 | 15 | |
| p | 0.3250 | 0.3049 | 0.5542 | 0.2000 | ||
| HTest | 0.9 | 1.0 | 0.3 | 2.7 | ||
| EA | W | 13 | 28 | 10 | 50 | 12 |
| Rest | 34 | 22 | 10 | 51 | 17 | |
| E | 23 | 24 | 9 | 50 | 17 | |
| p | 0.6889 | 0.9677 | 0.9456 | 0.5978 | ||
| HTest | 0.7 | 0.1 | 1.1 | 1.0 |
| Variable | Range | n | Ar | Dt | Sh | Bt |
|---|---|---|---|---|---|---|
| Life Zone | TAMT | 8 | 42 b | 17 b | 36 a | 5 a |
| TAWT | 2 | 39 ab | 18 ab | 39 ab | 5 ab | |
| TSDS | 43 | 18 a | 8 a | 52 ab | 22 b | |
| TSMF | 17 | 29 b | 8 a | 55 b | 8 a | |
| p | 0.0006 | 0.0446 | 0.0426 | 0.0001 | ||
| HTest | 17.3 | 7.9 | 8.2 | 21.8 | ||
| AMT | <5 | 15 | 34 b | 15 b | 39 a | 11 |
| 5–6 | 33 | 22 a | 9 a | 52 b | 17 | |
| >6 | 22 | 20 a | 7 a | 55 b | 18 | |
| p | 0.0340 | 0.0232 | 0.0199 | 0.0629 | ||
| HTest | 6.8 | 7.5 | 7.8 | 5.5 | ||
| AP | <150 | 26 | 24 | 9 | 50 | 17 |
| >150 | 44 | 24 | 10 | 51 | 16 | |
| p | 0.9399 | 0.6171 | 0.9890 | 0.3618 | ||
| HTest | <0.01 | 0.3 | <0.01 | 0.9 | ||
| HF | 0 | 53 | 25 | 10 | 50 | 15 |
| >0 | 17 | 20 | 8 | 50 | 21 | |
| p | 0.2758 | 0.5673 | 0.9891 | 0.1700 | ||
| HTest | 1.2 | 0.3 | <0.01 | 1.9 | ||
| DTT | <5 | 13 | 14 a | 6 | 58 | 23 b |
| 5–10 | 32 | 23 a | 9 | 51 | 17 ab | |
| >10 | 25 | 31 b | 11 | 45 | 12 a | |
| p | 0.0039 | 0.2661 | 0.1225 | 0.0436 | ||
| HTest | 11.0 | 2.6 | 4.2 | 6.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López, V.L.; Bottan, L.; Martínez Pastur, G.; Lencinas, M.V.; Cuyckens, G.A.E.; Cellini, J.M. Characterization of Polylepis tarapacana Life Forms in the Highest-Elevation Altiplano in South America: Influence of the Topography, Climate and Human Uses. Plants 2023, 12, 1806. https://doi.org/10.3390/plants12091806
López VL, Bottan L, Martínez Pastur G, Lencinas MV, Cuyckens GAE, Cellini JM. Characterization of Polylepis tarapacana Life Forms in the Highest-Elevation Altiplano in South America: Influence of the Topography, Climate and Human Uses. Plants. 2023; 12(9):1806. https://doi.org/10.3390/plants12091806
Chicago/Turabian StyleLópez, Victoria Lien, Lucia Bottan, Guillermo Martínez Pastur, María Vanessa Lencinas, Griet An Erica Cuyckens, and Juan Manuel Cellini. 2023. "Characterization of Polylepis tarapacana Life Forms in the Highest-Elevation Altiplano in South America: Influence of the Topography, Climate and Human Uses" Plants 12, no. 9: 1806. https://doi.org/10.3390/plants12091806