

Effect of Abiotic Stresses from Drought, Temperature, and Density on Germination and Seedling Growth of Barley (Hordeum vulgare L.)
 , , ,
, , ,
Abstract
:1. Introduction
1.1. Specific Aims of the Study
1.2. Novelty
2. Materials and Methods
2.1. Temperature Experiment
2.2. Water Availability Experiment
2.3. Seed and Seedling Density Experiment
2.4. Antifungal Experiment
2.5. Statistical Analysis
3. Results
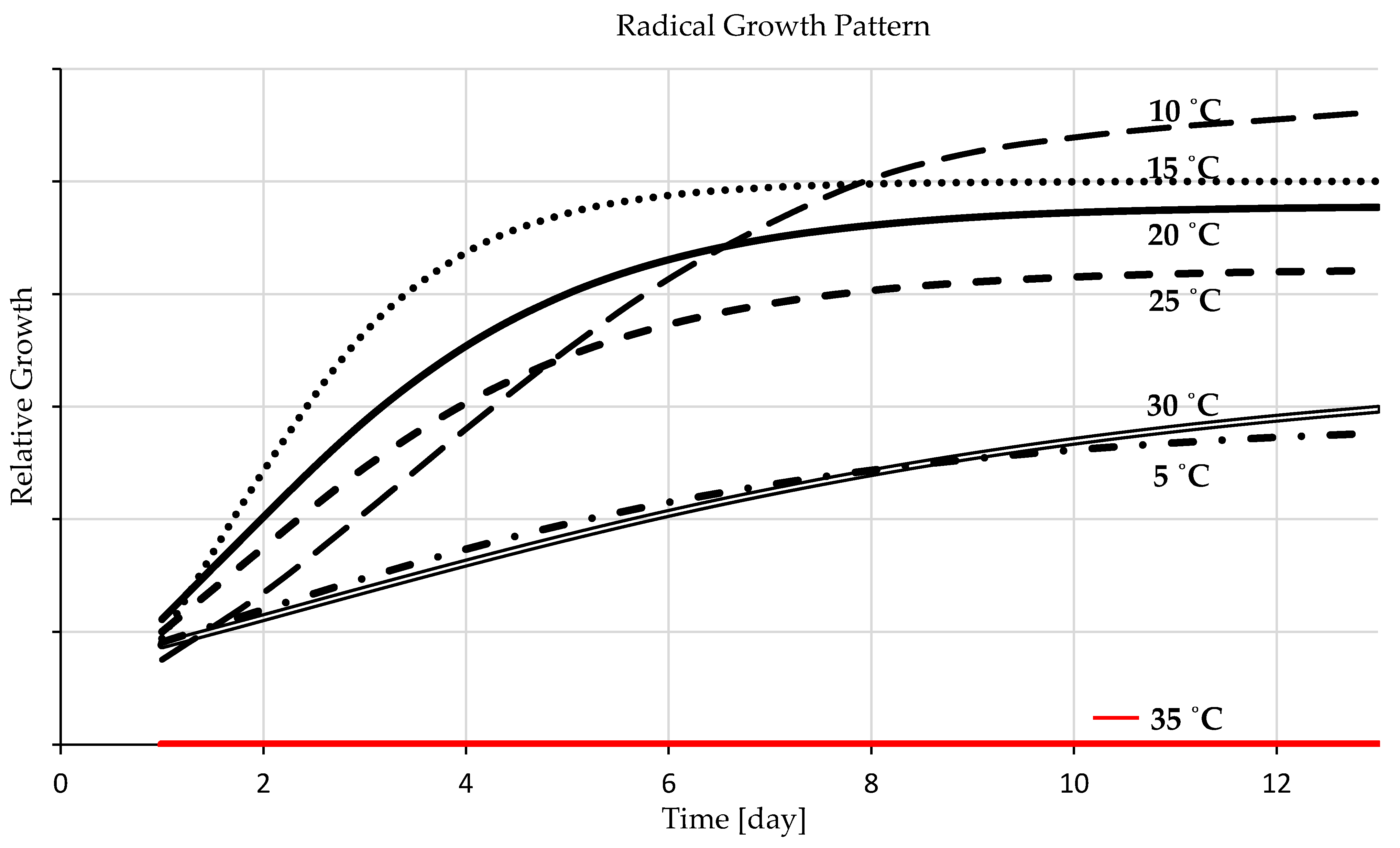
3.1. Temperature Experiment
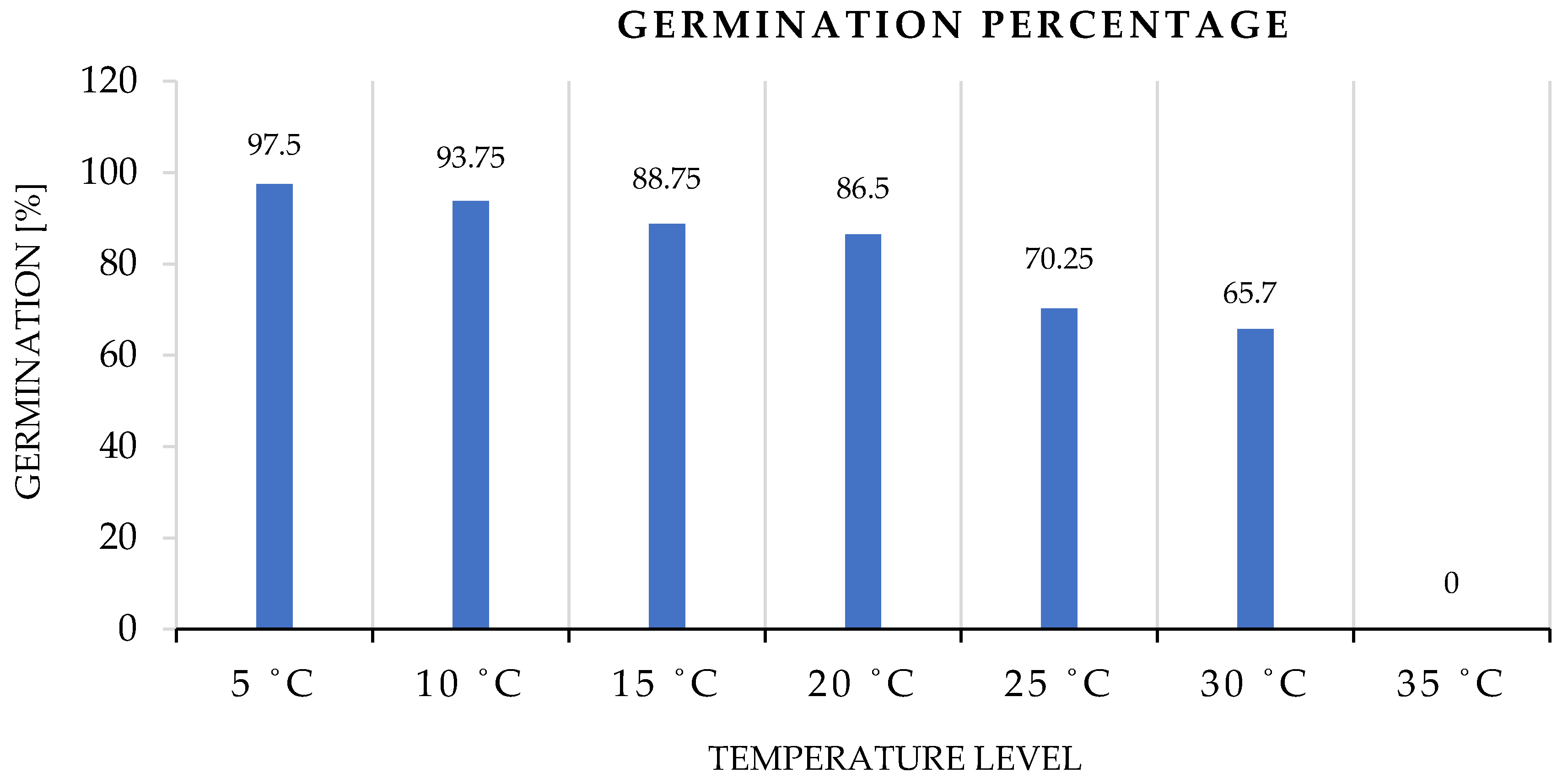
3.1.1. Germination Percentage
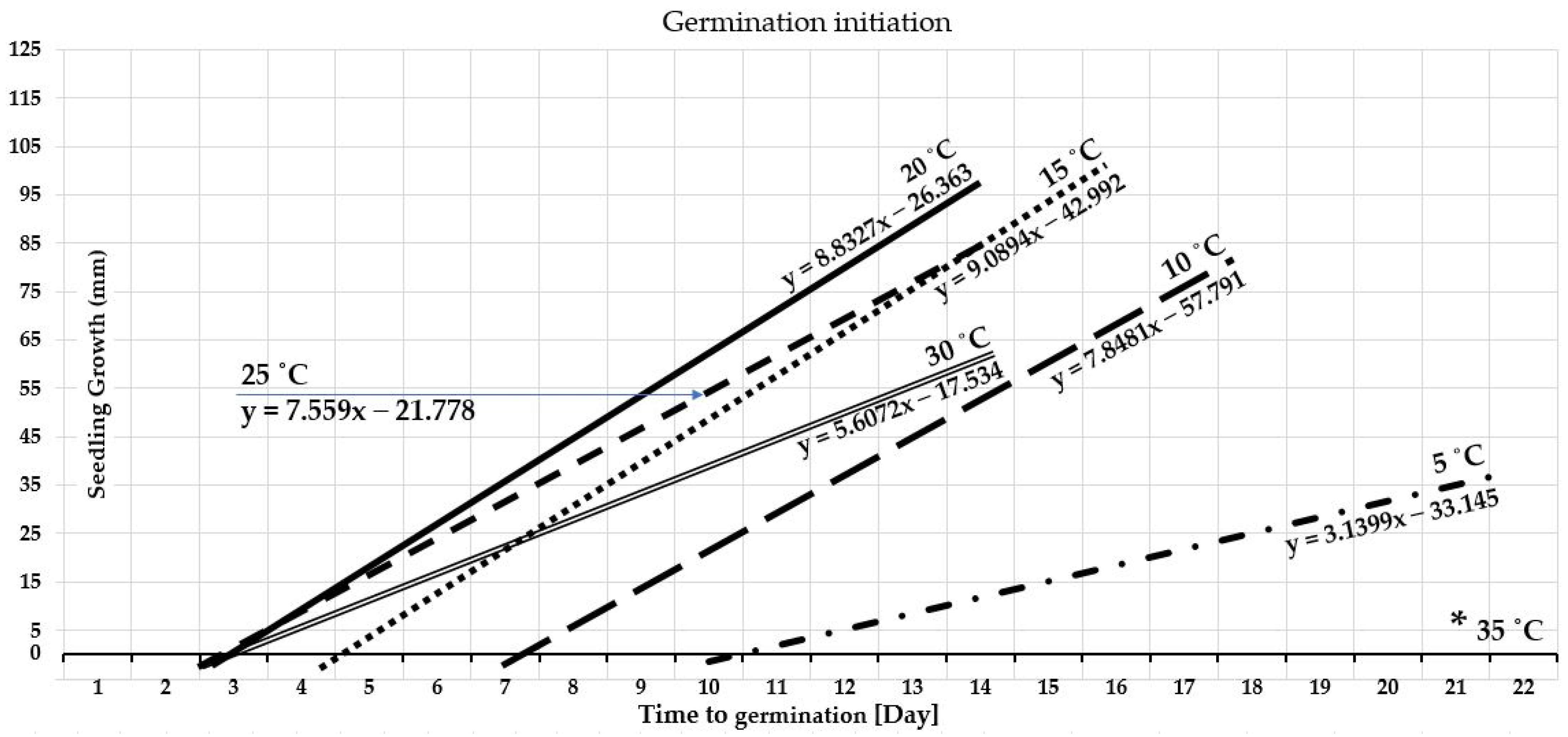
3.1.2. Germination Duration
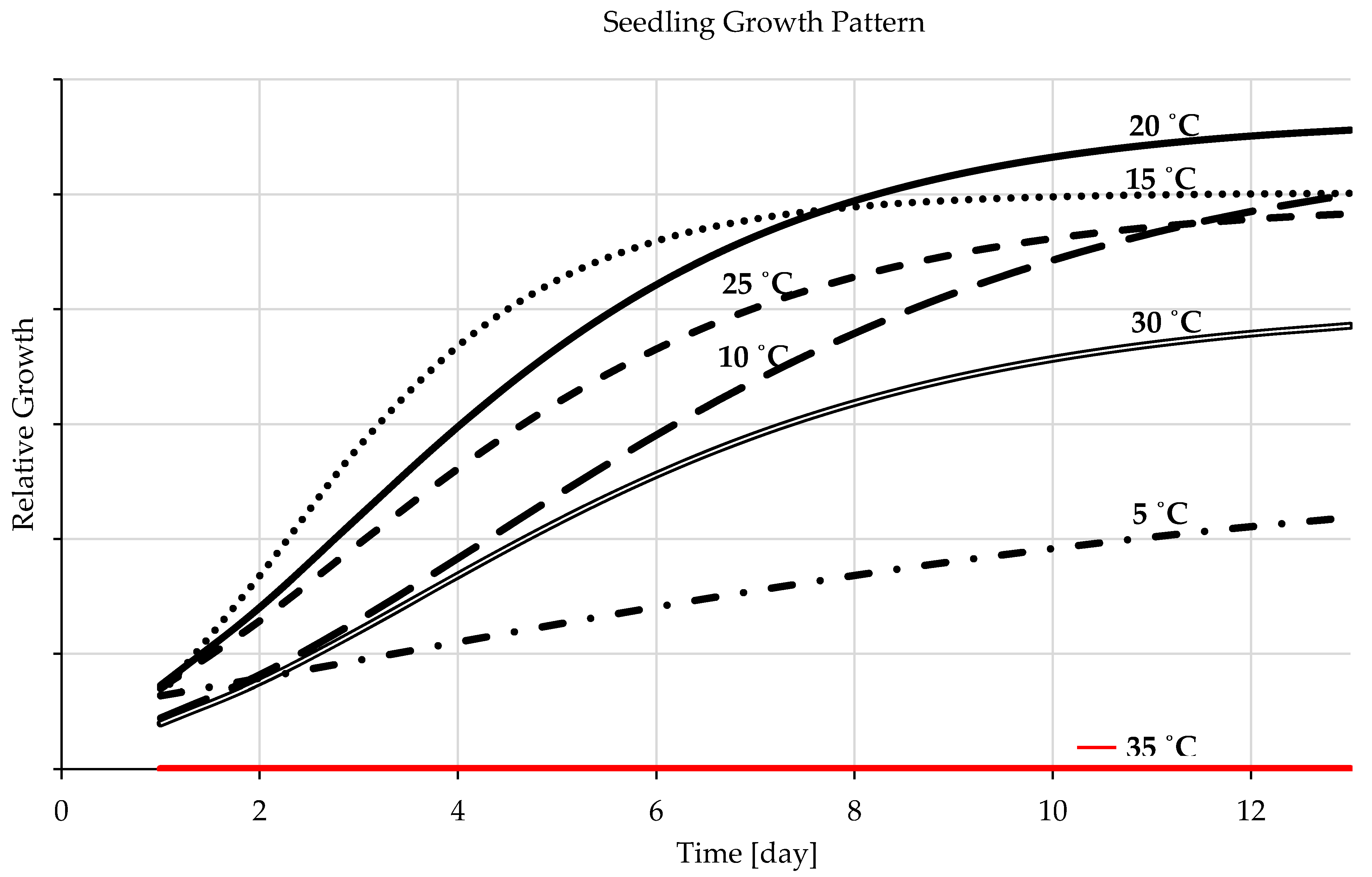
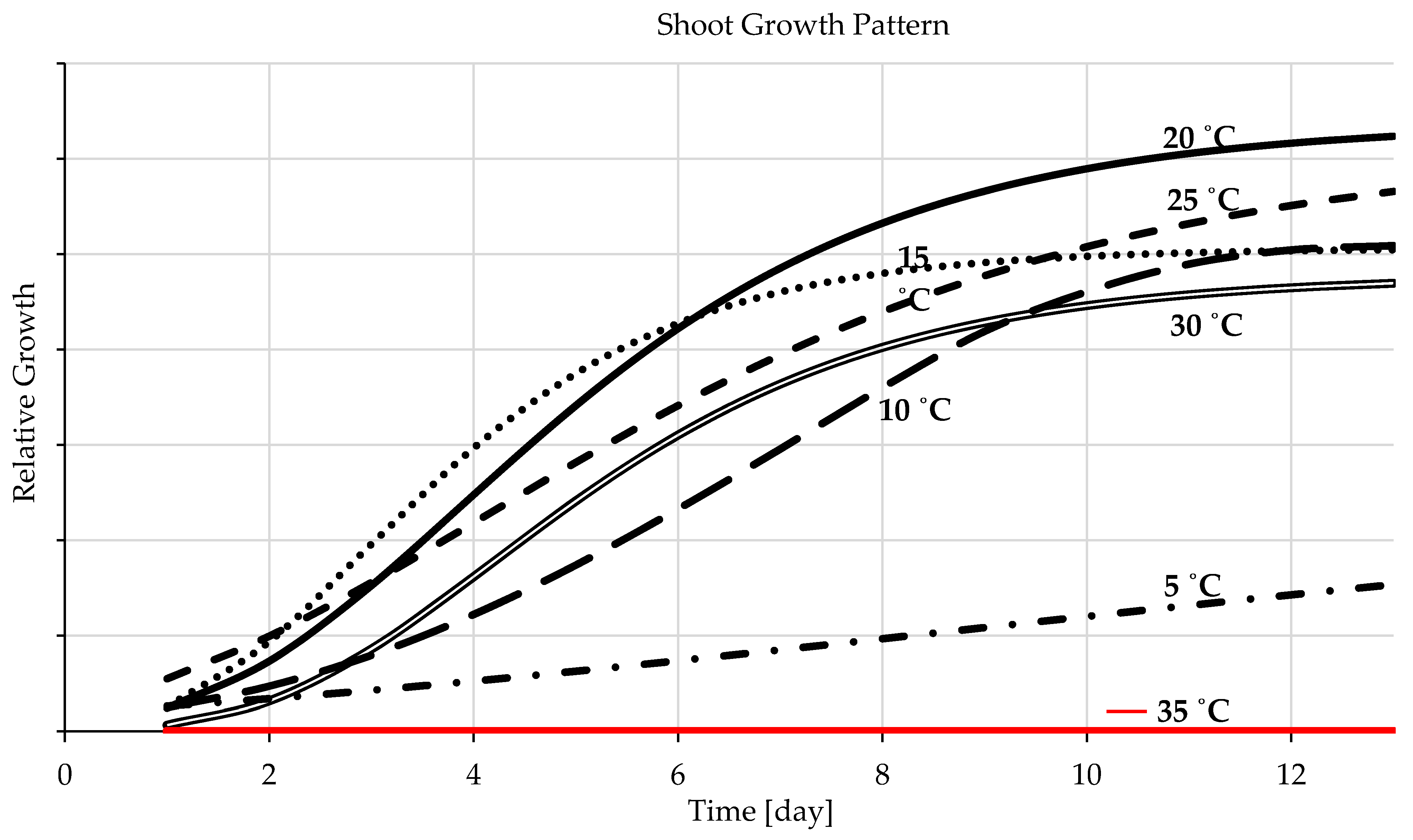
3.1.3. Seedling Development
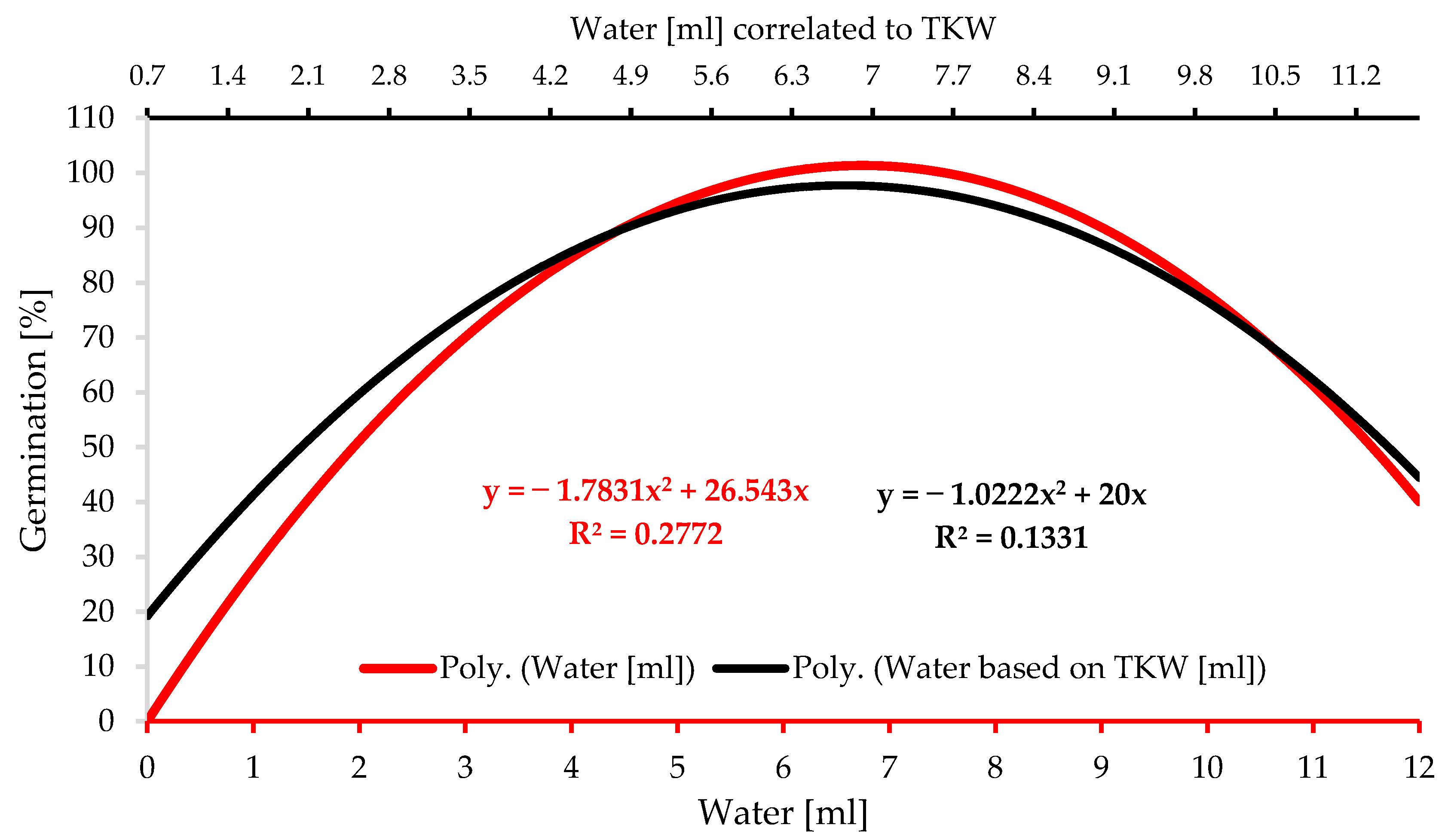
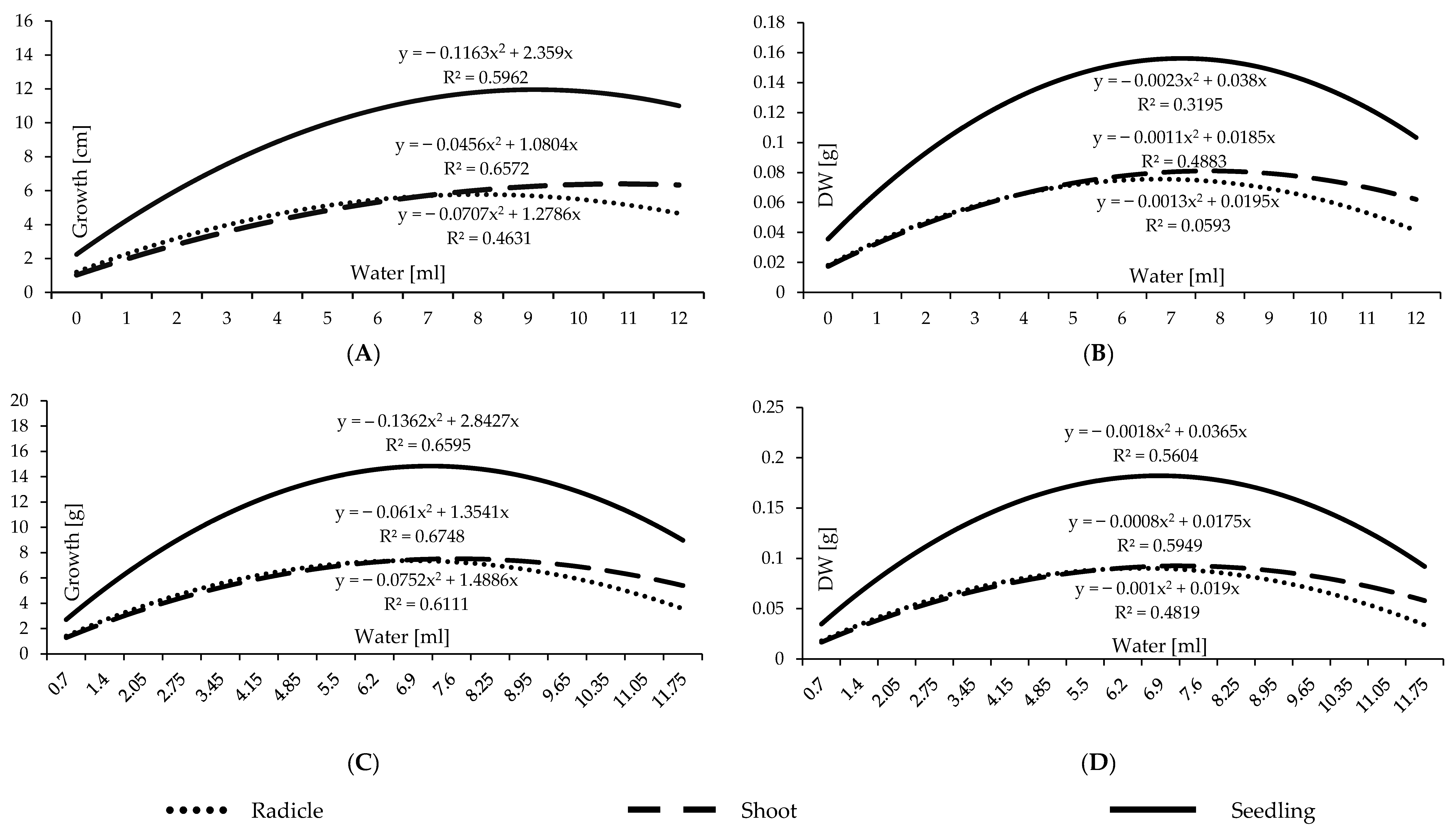
3.2. Water Availability Experiment
3.3. Seed and Seedling Density Experiment
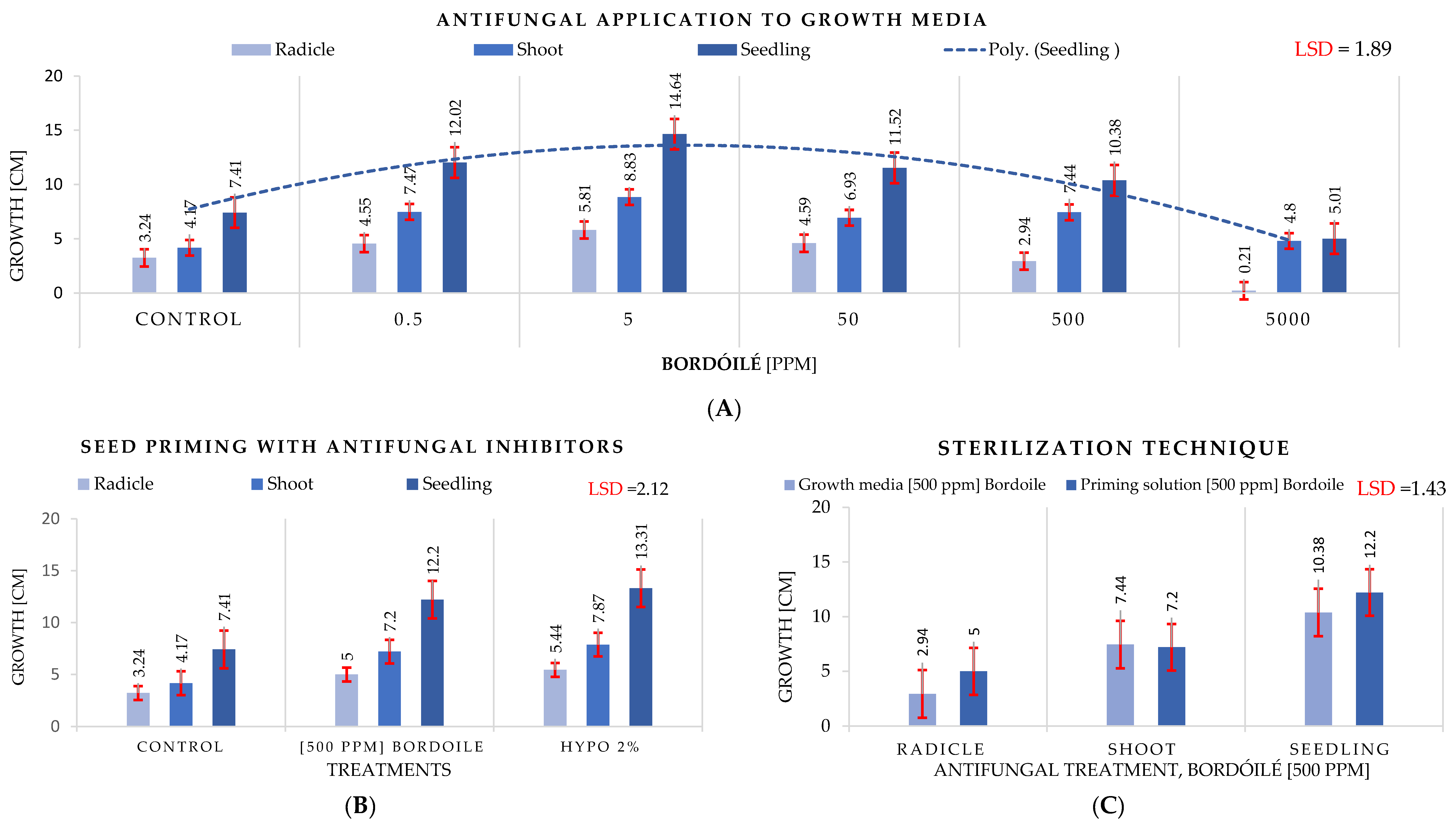
3.4. Antifungal Experiment
4. Discussion
4.1. Temperature Experiment
4.1.1. Germination Percentage
4.1.2. Germination Duration
4.1.3. Seedling Development
4.2. Water Availability Experiment
4.3. Seed and Seedling Density Experiment
4.4. Antifungal Experiment
5. Conclusions
- The sigmoid curves possess a solid fit for the experimental data representing seedling growth temperatures. The ideal temperature for barley seedling development is 20 °C followed by 15 °C and the range in between, and a more comprehensive range for germination rate is from 20 to 30 °C. A temperature lower than the optimal range decreases the germination rate (germination speed) but increases germination percentages, and a higher temperature raises fungal development.
- Seed size impacts the demanded level of water for germination. Hence, TKW administrates a more accurate base for water level application. Barley seed germination in different percentages can be initiated in a wide range of water levels starting at 1 mL, representing 130% of the barley TKW, but the optimal range for germination is 2.05 to 4.15 mL, representing 270–540% of the TKW. The optimal range for seedling growth is 5.5 mL to 8.25 mL, equivalent to 720% and 1080% of TKW.
- The dry weight measurement can be a reliable reference for seedling development since dry matter accumulation is compatible and consistent with the physical measurement of seedling development.
- There is no discernible difference in planting densities for seeds and seedlings in vitro; hence, planting densities as low as 10 barley seeds per 9 cm PD are advised for laboratory study.
- Both studied antifungal treatment techniques, seed priming and direct antifungal application at 5 ppm, effectively suppress fungal development. Seed priming is slightly superior to direct antifungal application on growth media, and both are significantly superior to the control.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
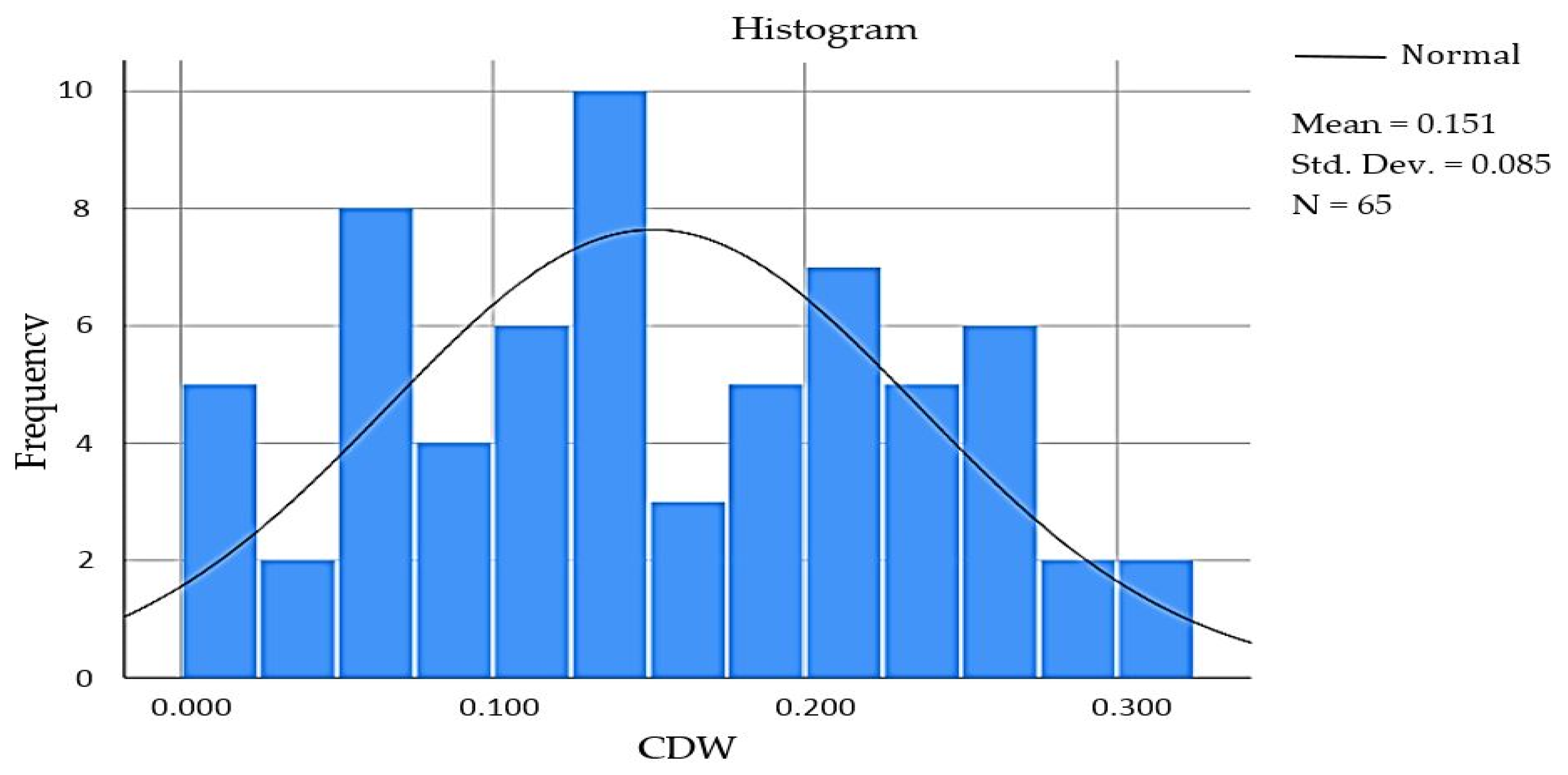
| a Kolmogorov–Smirnov | Shapiro–Wilk | |||||
|---|---|---|---|---|---|---|
| Statistic | df | Sig. | Statistic | df | Sig. | |
| CDW | 0.081 | 65 | 0.200 * | 0.967 | 65 | 0.084 |

References
- Abbasi, S.; Eslahi, N. Improved defense responses in modern barley cultivars compared to wild relatives against Rhizoctonia solani L. Rhizosphere 2022, 21, 100442. [Google Scholar] [CrossRef]
- Newman, C.W.; Newman, R.K. A Brief History of Barley Foods. Cereal Foods World 2006, 51, 4–7. [Google Scholar] [CrossRef]
- Panasiewicz, M. Economic and Academic Importance of Barley; Springer International Publishing: Cham, Switzerland, 2007; Volume 57. [Google Scholar] [CrossRef]
- Newton, A.C.; Flavell, A.J.; George, T.; Leat, P.; Mullholland, B.; Ramsay, L.; Revoredo-Giha, C.; Russell, J.; Steffenson, B.J.; Swanston, J.S.; et al. Crops that feed the world 4. Barley: A resilient crop? Strengths and weaknesses in the context of food security. Food Secur. 2011, 3, 141–178. [Google Scholar] [CrossRef]
- Giraldo, P.; Benavente, E.; Manzano-Agugliaro, F.; Gimenez, E. Worldwide Research Trends on Wheat and Barley: A Bibliometric Comparative Analysis. Agronomy 2019, 9, 352. [Google Scholar] [CrossRef]
- Yin, Y.; Xu, J.; He, X.; Yang, Z.; Fang, W.; Tao, J. Role of exogenous melatonin involved in phenolic acid metabolism of germinated hulless barley under NaCl stress. Plant Physiol. Biochem. 2022, 170, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Chen, Z.; Liang, F.; Zhou, W.; Li, T.; Tu, Z.; Tan, Y.; Li, J. Effects of hypoxia stress germination on nutrients, physicochemical properties and cooking characteristics of highland barley. J. Cereal Sci. 2022, 103, 103411. [Google Scholar] [CrossRef]
- Thabet, S.G.; Alomari, D.Z.; Alqudah, A.M. Exploring natural diversity reveals alleles to enhance antioxidant system in barley under salt stress. Plant Physiol. Biochem. 2021, 166, 789–798. [Google Scholar] [CrossRef]
- Zhao, H.; Ni, S.; Cai, S.; Zhang, G. Comprehensive dissection of primary metabolites in response to diverse abiotic stress in barley at seedling stage. Plant Physiol. Biochem. 2021, 161, 54–64. [Google Scholar] [CrossRef]
- Boyer, J.S. Plant Productivity and Environment. Science 1982, 218, 443–448. [Google Scholar] [CrossRef]
- Khaeim, H.; Kende, Z.; Jolánkai, M.; Kovács, G.P.; Gyuricza, C.; Tarnawa, Á. Impact of Temperature and Water on Seed Germination and Seedling Growth of Maize (Zea mays L.). Agronomy 2022, 12, 397. [Google Scholar] [CrossRef]
- Osama, S.K.; Kerr, E.D.; Yousif, A.M.; Phung, T.K.; Kelly, A.M.; Fox, G.P.; Schulz, B.L. Proteomics reveals commitment to germination in barley seeds is marked by loss of stress response proteins and mobilisation of nutrient reservoirs. J. Proteom. 2021, 242, 104221. [Google Scholar] [CrossRef] [PubMed]
- Khaeim, H.; Kende, Z.; Balla, I.; Gyuricza, C.; Eser, A.; Tarnawa, Á. The Effect of Temperature and Water Stresses on Seed Germination and Seedling Growth of Wheat (Triticum aestivum L.). Sustainability 2022, 14, 3887. [Google Scholar] [CrossRef]
- Chen, P.; Zhang, H.; Zhang, Y.; Yang, W.; Wang, L.; Wen, Z. Interactions between fruiting perianth and various abiotic factors differentially affect seed germination of Salsola brachiata. Flora 2022, 290, 152057. [Google Scholar] [CrossRef]
- Xue, X.; Du, S.; Jiao, F.; Xi, M.; Wang, A.; Xu, H.; Jiao, Q.; Zhang, X.; Jiang, H.; Chen, J.; et al. The regulatory network behind maize seed germination: Effects of temperature, water, phytohormones, and nutrients. Crops J. 2021, 9, 718–724. [Google Scholar] [CrossRef]
- Gavkosh, M.Z.; Afshari, R.T.; Jahansooz, M. Morphophysiological dormancy in smyrnium cordifolium boiss: Germination requirements and embryo growth. J. Appl. Res. Med. Aromat. Plants 2022, 30, 100385. [Google Scholar] [CrossRef]
- Aderounmu, A.F.; Nkemnkeng, F.J.; Anjah, G.M. Effects of seed provenance and growth media on the growth performance of Vitellaria paradoxa C.F. Gaertn. Int. J. Biol. Chem. Sci. 2020, 14, 2659–2669. [Google Scholar] [CrossRef]
- Mangena, P. Analysis of correlation between seed vigour, germination and multiple shoot induction in soybean (Glycine max L. Merr.). Heliyon 2021, 7, e07913. [Google Scholar] [CrossRef]
- Le, H.; Browning, K.S.; Gallie, D.R. The phosphorylation state of the wheat translation initiation factors eif4b, eif4a, and eif2 is differentially regulated during seed development and germination. J. Biol. Chem. 1998, 273, 20084–20089. [Google Scholar] [CrossRef]
- El-Badri, A.M.; Batool, M.; Wang, C.; Hashem, A.M.; Tabl, K.M.; Nishawy, E.; Kuai, J.; Zhou, G.; Wang, B. Selenium and zinc oxide nanoparticles modulate the molecular and morpho-physiological processes during seed germination of Prassica napus under salt stress. Ecotoxicol. Environ. Saf. 2021, 225, 112695. [Google Scholar] [CrossRef]
- Manz, B.; Muller, K.; Kucera, B.; Volke, F.; Leubner-Metzger, G. Water Uptake and Distribution in Germinating Tobacco Seeds Investigated In Vivo by Nuclear Magnetic Resonance Imaging. Plant Physiol. 2020, 138, 1538–1551. [Google Scholar] [CrossRef]
- Ofoe, R.; Gunupuru, L.R.; Wang-Pruski, G.; Fofana, B.; Thomas, R.H.; Abbey, L. Seed priming with pyroligneous acid mitigates aluminum stress, and promotes tomato seed germination and seedling growth. Plant Stress 2022, 4, 100083. [Google Scholar] [CrossRef]
- Liu, Y.; Han, C.; Deng, X.; Liu, D.; Liu, N.; Yan, Y. Integrated physiology and proteome analysis of embryo and endosperm highlights complex metabolic networks involved in seed germination in wheat (Triticum aestivum L.). J. Plant Physiol. 2018, 229, 63–76. [Google Scholar] [CrossRef] [PubMed]
- De Silva, Y.S.K.; Rajagopalan, U.M.; Kadono, H.; Li, D. Positive and negative phenotyping of increasing zn concentrations by biospeckle optical coherence tomography in speedy monitoring on lentil (Lens culinaris L.) seed germination and seedling growth. Plant Stress 2021, 2, 100041. [Google Scholar] [CrossRef]
- da Silva, F.T.; de Oliveira, J.P.; Fonseca, L.M.; Bruni, G.P.; Zavareze, E.D.R.; Dias, A.R.G. Physically cross-linked aerogels based on germinated and non-germinated wheat starch and PEO for application as water absorbers for food packaging. Int. J. Biol. Macromol. 2020, 155, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Yin, Y.; Liu, C.; Zhao, Z.; Guo, M. Effect of germination time on the compositional, functional and antioxidant properties of whole wheat malt and its end-use evaluation in cookie-making. Food Chem. 2021, 349, 129125. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R. Seed and seedling vigour in relation to crop growth and yield. Plant Growth Regul. 1992, 11, 249–255. [Google Scholar] [CrossRef]
- Barrero, J.M.; Jacobsen, J.V.; Talbot, M.J.; White, R.G.; Swain, S.M.; Garvin, D.F.; Gubler, F. Grain dormancy and light quality effects on germination in the model grass Brachypodium distachyon. New Phytol. 2012, 193, 376–386. [Google Scholar] [CrossRef]
- Tao, Y.; An, L.; Xiao, F.; Li, G.; Ding, Y.; Paul, M.J.; Liu, Z. Integration of embryo–endosperm interaction into a holistic and dynamic picture of seed development using a rice mutant with notched-belly kernels. Crops J. 2021, 10, 729–742. [Google Scholar] [CrossRef]
- Rizzardi, M.A.; Luiz, A.R.; Roman, E.S.; Vargas, L. Temperatura cardeal e potencial hídrico na germinação de sementes de corda-de-viola (Ipomoea triloba). Planta Daninha 2009, 27, 13–21. [Google Scholar] [CrossRef]
- Cone, J.W.; Spruit, C.J.P. Imbibition conditions and seed dormancy of Arabidopsis thaliana. Physiol. Plant. 1983, 59, 416–420. [Google Scholar] [CrossRef]
- Ozden, E.; Light, M.E.; Demir, I. Alternating temperatures increase germination and emergence in relation to endogenous hormones and enzyme activities in aubergine seeds. S. Afr. J. Bot. 2021, 139, 130–139. [Google Scholar] [CrossRef]
- Magangana, T.; Makunga, N. The effect of various factors on seed germination and the influence of abiotic stresses on growth productivity, physiology and differences in metabolite profiles (Diterpene glycosides L.) of stevia rebaudiana bertoni. S. Afr. J. Bot. 2016, 103, 324–325. [Google Scholar] [CrossRef]
- Albertos, P.; Tatematsu, K.; Mateos, I.; Sánchez-Vicente, I.; Fernández-Arbaizar, A.; Nakabayashi, K.; Nambara, E.; Godoy, M.; Franco, J.M.; Solano, R.; et al. Redox feedback regulation of ANAC089 signaling alters seed germination and stress response. Cell Rep. 2021, 35, 109263. [Google Scholar] [CrossRef] [PubMed]
- Rajora, N.; Vats, S.; Raturi, G.; Thakral, V.; Kaur, S.; Rachappanavar, V.; Kumar, M.; Kesarwani, A.K.; Sonah, H.; Sharma, T.R.; et al. Seed priming with melatonin: A promising approach to combat abiotic stress in plants. Plant Stress 2022, 4, 100071. [Google Scholar] [CrossRef]
- Lana, L.G.; de Araújo, L.M.; Silva, T.F.; Modolo, L.V. Interplay between gasotransmitters and potassium is a K+ey factor during plant response to abiotic stress. Plant Physiol. Biochem. 2021, 169, 322–332. [Google Scholar] [CrossRef]
- Bradford, K.J.; Nonogaki, H. Seed Development, Dormancy and Germination; Blackwell Publishing: Oxford, UK, 2007; pp. 1–367. [Google Scholar] [CrossRef]
- Bentsink, L.; Koornneef, M. Seed Dormancy and Germination. Arab. Book 2008, 6, e0119. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Li, Z.; Yang, X.; He, Y.; Wang, X. Production of endo-β-mannanase, superoxide radicals, hydrogen peroxide, and peroxidase in the micropylar endosperm of pepper seed during germination is determined by the endosperm itself. Sci. Hortic. 2021, 294, 110757. [Google Scholar] [CrossRef]
- Sánchez-Montesino, R.; Bouza-Morcillo, L.; Marquez, J.; Ghita, M.; Duran-Nebreda, S.; Gómez, L.; Holdsworth, M.J.; Bassel, G.; Oñate-Sánchez, L. A Regulatory Module Controlling GA-Mediated Endosperm Cell Expansion Is Critical for Seed Germination in Arabidopsis. Mol. Plant 2019, 12, 71–85. [Google Scholar] [CrossRef]
- Liu, G.; Jiang, W.; Tian, L.; Fu, Y.; Tan, L.; Zhu, Z.; Sun, C.; Liu, F. Polyamine oxidase 3 is involved in salt tolerance at the germination stage in rice. J. Genet. Genom. 2022, 49, 458–468. [Google Scholar] [CrossRef]
- Basahi, M. Seed germination with titanium dioxide nanoparticles enhances water supply, reserve mobilization, oxidative stress and antioxidant enzyme activities in pea. Saudi J. Biol. Sci. 2021, 28, 6500–6507. [Google Scholar] [CrossRef]
- Lukács, H.; Jócsák, I.; Somfalvi-Tóth, K.; Keszthelyi, S. Physiological Responses Manifested by Some Conventional Stress Parameters and Biophoton Emission in Winter Wheat as a Consequence of Cereal Leaf Beetle Infestation. Front. Plant Sci. 2022, 13, 1755. [Google Scholar] [CrossRef] [PubMed]
- Kunos, V.; Cséplő, M.; Seress, D.; Eser, A.; Kende, Z.; Uhrin, A.; Bányai, J.; Bakonyi, J.; Pál, M.; Mészáros, K. The Stimulation of Superoxide Dismutase Enzyme Activity and Its Relation with the Pyrenophora teres f. teres Infection in Different Barley Genotypes. Sustainability 2022, 14, 2597. [Google Scholar] [CrossRef]
- Kovács, Z.; Bedő, J.; Pápai, B.; Tóth-Lencsés, A.K.; Csilléry, G.; Szőke, A.; Bányai-Stefanovits, É.; Kiss, E.; Veres, A. Ripening-Induced Changes in the Nutraceutical Compounds of Differently Coloured Pepper (Capsicum annuum L.) Breeding Lines. Antioxidants 2022, 11, 637. [Google Scholar] [CrossRef] [PubMed]
- Kutasy, E.; Buday-Bódi, E.; Virág, I.C.; Forgács, F.; Melash, A.A.; Zsombik, L.; Nagy, A.; Csajbók, J. Mitigating the Negative Effect of Drought Stress in Oat (Avena sativa L.) with Silicon and Sulphur Foliar Fertilization. Plants 2022, 11, 30. [Google Scholar] [CrossRef] [PubMed]
- Polgári, D.; Mihók, E.; Sági, L. Composition and random elimination of paternal chromosomes in a large population of wheat × barley (Triticum aestivum L. × Hordeum vulgare L.) hybrids. Plant Cell Rep. 2019, 38, 767–775. [Google Scholar] [CrossRef]
- Thakur, P.S.; Chatterjee, A.; Rajput, L.S.; Rana, S.; Bhatia, V.; Prakash, S. Laser biospeckle technique for characterizing the impact of temperature and initial moisture content on seed germination. Opt. Lasers Eng. 2022, 153, 106999. [Google Scholar] [CrossRef]
- Wang, G.; Lynch, A.L.; Cruz, V.M.V.; Heinitz, C.C.; Dierig, D.A. Temperature requirements for guayule seed germination. Ind. Crops Prod. 2020, 157, 112934. [Google Scholar] [CrossRef]
- Brits, G. Influence of fluctuating temperatures and H2O2 treatment on germination of Leucospermum cordifolium and Serruina florida (Proteaceae) seeds. S. Afr. J. Bot. 1986, 52, 286–290. [Google Scholar] [CrossRef]
- Gong, M.; Chen, B.; Li, Z.-G.; Guo, L.-H. Heat-shock-induced cross adaptation to heat, chilling, drought and salt stress in maize seedlings and involvement of H2O2. J. Plant Physiol. 2001, 158, 1125–1130. [Google Scholar] [CrossRef]
- Bailly, C. The signalling role of ROS in the regulation of seed germination and dormancy. Biochem. J. 2019, 476, 3019–3032. [Google Scholar] [CrossRef]
- Poudel, R.; Finnie, S.; Rose, D.J. Effects of wheat kernel germination time and drying temperature on compositional and end-use properties of the resulting whole wheat flour. J. Cereal Sci. 2019, 86, 33–40. [Google Scholar] [CrossRef]
- del Rosario Moguel Concha, D.; Martínez, J.E.B.; Velázquez, T.G.G.; Martínez, C.J.; Ruiz, J.C.R. Impact of germination time on protein solubility and anti-inflammatory properties of Pisum sativum L. grains. Food Chem. X 2022, 13, 100219. [Google Scholar] [CrossRef] [PubMed]
- Bradford, K.J. A Water Relations Analysis of Seed Germination Rates. Plant Physiol. 1990, 94, 840–849. [Google Scholar] [CrossRef]
- Obroucheva, N.V.; Sinkevich, I.A.; Lityagina, S.V.; Novikova, G.V. Water relations in germinating seeds. Russ. J. Plant Physiol. 2017, 64, 625–633. [Google Scholar] [CrossRef]
- Khaeim, H.M.; Jeber, B.A.; Ali, M.A. Winter wheat genotypes response to different water quality. Int. J. Agric. Stat. Sci. 2019, 15, 669–676. [Google Scholar]
- Dürr, C.; Dickie, J.; Yang, X.-Y.; Pritchard, H. Ranges of critical temperature and water potential values for the germination of species worldwide: Contribution to a seed trait database. Agric. For. Meteorol. 2015, 200, 222–232. [Google Scholar] [CrossRef]
- El Hamdaoui, A.; Mechqoq, H.; El Yaagoubi, M.; Bouglad, A.; Hallouti, A.; El Mousadik, A.; El Aouad, N.; Ben Aoumar, A.A.; Msanda, F. Effect of pretreatment, temperature, gibberellin (GA3), salt and water stress on germination of Lavandula mairei Humbert. J. Appl. Res. Med. Aromat. Plants 2021, 24, 100314. [Google Scholar] [CrossRef]
- Tyagi, A.; Chen, X.; Shabbir, U.; Chelliah, R.; Oh, D.H. Effect of slightly acidic electrolyzed water on amino acid and phenolic profiling of germinated brown rice sprouts and their antioxidant potential. LWT 2022, 157, 113119. [Google Scholar] [CrossRef]
- Contreras-Negrete, G.; Pineda-García, F.; Nicasio-Arzeta, S.; De la Barrera, E.; González-Rodríguez, A. Differences in germination response to temperature, salinity, and water potential among Prosopis laevigata populations are guided by the tolerance-exploitation trade-off. Flora 2021, 285, 151963. [Google Scholar] [CrossRef]
- Aliakbari, M.; Cohen, S.P.; Lindlöf, A.; Shamloo-Dashtpagerdi, R. Rubisco activase A (RcaA) is a central node in overlapping gene network of drought and salinity in Barley (Hordeum vulgare L.) and may contribute to combined stress tolerance. Plant Physiol. Biochem. 2021, 161, 248–258. [Google Scholar] [CrossRef]
- Demirel, U.; Morris, W.L.; Ducreux, L.J.M.; Yavuz, C.; Asim, A.; Tindas, I.; Campbell, R.; Morris, J.A.; Verrall, S.R.; Hedley, P.; et al. Physiological, Biochemical, and Transcriptional Responses to Single and Combined Abiotic Stress in Stress-Tolerant and Stress-Sensitive Potato Genotypes. Front. Plant Sci. 2020, 11, 169. [Google Scholar] [CrossRef]
- Bashir, K.; Brain, R.; Institute, S.; Biswapriya, J.; Misra, B.; Gürel, F.; Öztürk, Z.N.; Uçarlı, C.; Rosellini, D. Barley Genes as Tools to Confer Abiotic Stress Tolerance in Crops. Front. Plant Sci. 2016, 7, 1137. [Google Scholar] [CrossRef]
- Drebee, H.A.; Razak, N.A.A. Measuring the efficiency of colleges at the university of al-qadisiyah-iraq: A data envelopment analysis approach. J. Ekon. Malays. 2018, 52, 163–179. [Google Scholar] [CrossRef]
- Bulmer, M.G.; Harrison, T.K. Reviewed Work: Principles of Statistics. J. R. Stat. Society. Ser. D. Stat. 1966, 16, 217. [Google Scholar] [CrossRef]
- Seefeldt, S.S.; Kidwell, K.K.; Waller, J.E. Base growth temperatures, germination rates and growth response of contemporary spring wheat (Triticum aestivum L.) cultivars from the US Pacific Northwest. Field Crops Res. 2002, 75, 47–52. [Google Scholar] [CrossRef]
- Qin, Q.; Liu, J.; Hu, S.; Dong, J.; Yu, J.; Fang, L.; Huang, S.; Wang, L. Comparative proteomic analysis of different barley cultivars during seed germination. J. Cereal Sci. 2021, 102, 103357. [Google Scholar] [CrossRef]
- Dong, X.; Sun, L.; Agarwal, M.; Maker, G.; Han, Y.; Yu, X.; Ren, Y. The Effect of Ozone Treatment on Metabolite Profile of Germinating Barley. Foods 2022, 11, 1211. [Google Scholar] [CrossRef]
- Galotta, M.F.; Pugliese, P.; Boem, F.H.G.; Veliz, C.G.; Criado, M.V.; Caputo, C.; Echeverria, M.; Roberts, I.N. Subtilase activity and gene expression during germination and seedling growth in barley. Plant Physiol. Biochem. 2019, 139, 197–206. [Google Scholar] [CrossRef]
- Al-Ansi, W.; Zhang, Y.; Alkawry, T.A.A.; Al-Adeeb, A.; Mahdi, A.A.; Al-Maqtari, Q.A.; Ahmed, A.; Mushtaq, B.S.; Fan, M.; Li, Y.; et al. Influence of germination on bread-making behaviors, functional and shelf-life properties, and overall quality of highland barley bread. LWT 2022, 159, 113200. [Google Scholar] [CrossRef]
- Benadjaoud, A.; Dadach, M.; El-Keblawy, A.; Mehdadi, Z. Impacts of osmopriming on mitigation of the negative effects of salinity and water stress in seed germination of the aromatic plant Lavandula stoechas L. J. Appl. Res. Med. Aromat. Plants 2022, 31, 100407. [Google Scholar] [CrossRef]
- Shaban, M. Effect of water and temperature on seed germination and emergence as a seed hydrothermal time model. Int. J. Adv. Biol. Biomed. Res. 2013, 1, 1686–1691. [Google Scholar]
- Gao, C.; Liu, F.; Zhang, C.; Feng, D.; Li, K.; Cui, K. Germination responses to water potential and temperature variation among provenances of Pinus yunnanensis. Flora 2021, 276–277, 151786. [Google Scholar] [CrossRef]
- Reza Yousefi, A.; Rashidi, S.; Moradi, P.; Mastinu, A. Germination and Seedling Growth Responses of Zygophyllum fabago, Salsola kali L. and Atriplex canescens to PEG-Induced Drought Stress. Environments 2020, 7, 107. [Google Scholar] [CrossRef]
- Ismaili, S.E.; Maurady, A.; Lachkar, M.; Britel, M.R.; Bakali, A.H. Effect of temperature and different pre-treatments on seed germination of Stachys mouretii Batt. & Pit. J. Appl. Res. Med. Aromat. Plants 2023, 32, 100438. [Google Scholar] [CrossRef]
- Liao, D.; Chen, Y.; Qi, J.; Zhang, H.; Sun, P.; Chen, C.; Li, X. Temporal transcriptomics reveal the molecular mechanism of dormancy and germination regulated by temperature in Paris polyphylla seed. Hortic. Plant J. 2022. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; Messéan, A.; Ricci, P. Research and innovation priorities as defined by the Ecophyto plan to address current crop protection transformation challenges in France. Adv. Agron. 2019, 154, 81–152. [Google Scholar] [CrossRef]
- Reddy, Y.N.; Ramya, V.; Suma, L.; Reddy, A.N.; Krishna, S.S. Drought adaptation: Approaches for crop improvement. In Millets Pseudo Cereals; Woodhead Publishing: Sawston, UK, 2020; pp. 143–158. [Google Scholar] [CrossRef]
- Sghaier, A.H.; Tarnawa, Á.; Khaeim, H.; Kovács, G.P.; Gyuricza, C.; Kende, Z. The Effects of Temperature and Water on the Seed Germination and Seedling Development of Rapeseed (Brassica napus L.). Plants 2022, 11, 2819. [Google Scholar] [CrossRef]
- Sghaier, A.H.; Khaeim, H.; Tarnawa, Á.; Kovács, G.P.; Gyuricza, C.; Kende, Z. Germination and Seedling Development Responses of Sunflower (Helianthus annuus L.) Seeds to Temperature and Different Levels of Water Availability. Agriculture 2023, 13, 608. [Google Scholar] [CrossRef]
- Boyce, D. Heat and moisture transfer in ventilated grain. J. Agric. Eng. Res. 1966, 11, 255–265. [Google Scholar] [CrossRef]
- Riley, G.J.P. Effects of high temperature on the germination of maize (Zea mays L.). Planta 1981, 151, 68–74. [Google Scholar] [CrossRef]
- Mukherjee, D.K.; Nair, M. Growth & Development. In Oski’s Pediatric Certification and Recertification Board Review; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2011. [Google Scholar] [CrossRef]
- Grains Research and Development Corporation. Barley—Plant growth and physiology. In GRDC Growthnotes; Grains Research and Development Corporation: Kingston, Australia, 2018; p. 4. [Google Scholar]
- Hossain, A.; da Silva, J.A.T.; Lozovskaya, M.V.; Zvolinsky, V.P. High temperature combined with drought affect rainfed spring wheat and barley in South-Eastern Russia: I. Phenology and growth. Saudi J. Biol. Sci. 2012, 19, 473–487. [Google Scholar] [CrossRef]
- Prerna, D.I.; Govindaraju, K.; Tamilselvan, S.; Kannan, M.; Vasantharaja, R.; Chaturvedi, S.; Shkolnik, D. Influence of nanoscale micro-nutrient α-Fe2O3 on seed germination, seedling growth, translocation, physiological effects and yield of rice (Oryza sativa L.) and maize (Zea mays L.). Plant Physiol. Biochem. 2021, 162, 564–580. [Google Scholar] [CrossRef]
- Basit, F.; Bhat, J.A.; Han, J.; Guan, Y.; Jan, B.L.; Shakoor, A.; Alansi, S. Screening of rice cultivars for Cr-stress response by using the parameters of seed germination, morpho-physiological and antioxidant analysis. Saudi J. Biol. Sci. 2022, 29, 3918–3928. [Google Scholar] [CrossRef]
- Matsuda, R.; Yamada, K.; Hayasaka, D.; Henmi, Y. Effects of salinity, temperature, and immersion conditions on seed germination of invasive Spartina alterniflora Loisel (smooth cordgrass) in Japan. Reg. Stud. Mar. Sci. 2023, 57, 102738. [Google Scholar] [CrossRef]
- Kong, Y.; Wang, G.; Tang, H.; Yang, J.; Yang, Y.; Wang, J.; Li, G.; Li, Y.; Yuan, J. Multi-omics analysis provides insight into the phytotoxicity of chicken manure and cornstalk on seed germination. Sci. Total. Environ. 2022, 861, 160611. [Google Scholar] [CrossRef]
- Sabouri, A.; Azizi, H.; Nonavar, M. Hydrotime model analysis of lemon balm (Melissa officinalis L.) using different distribution functions. S. Afr. J. Bot. 2020, 135, 158–163. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Zhang, D.; Jung, K.H. Molecular Basis of Pollen Germination in Cereals. Trends Plant Sci. 2019, 24, 1126–1136. [Google Scholar] [CrossRef]
- Teng, C.-F.; Gao, D.-H.; Ding, H.-C.; Yan, X.-H. Regulation of reactive oxygen species on production, release, and early germination of archeospores in Pyropia yezoensis. Algal Res. 2023, 69, 102909. [Google Scholar] [CrossRef]
- Lim, M.-N.; Lee, S.-E.; Chang, W.Y.; Yoon, I.S.; Hwang, Y.-S. Comparison of transcriptomic adjustments to availability of sugar, cellular energy, and oxygen in germinating rice embryos. J. Plant Physiol. 2021, 264, 153471. [Google Scholar] [CrossRef]
- He, J.; Hu, W.; Li, Y.; Zhu, H.; Zou, J.; Wang, Y.; Meng, Y.; Chen, B.; Zhao, W.; Wang, S.; et al. Prolonged drought affects the interaction of carbon and nitrogen metabolism in root and shoot of cotton. Environ. Exp. Bot. 2022, 197, 104839. [Google Scholar] [CrossRef]
- Peng, X.; Li, J.; Sun, L.; Gao, Y.; Cao, M.; Luo, J. Impacts of water deficit and post-drought irrigation on transpiration rate, root activity, and biomass yield of Festuca arundinacea during phytoextraction. Chemosphere 2022, 294, 133842. [Google Scholar] [CrossRef] [PubMed]
- Domańska, U.; Królikowska, M. Effect of temperature and composition on the surface tension and thermodynamic properties of binary mixtures of 1-butyl-3-methylimidazolium thiocyanate with alcohols. J. Colloid Interface Sci. 2010, 348, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Weidenhamer, J.D.; Morton, T.C.; Romeo, J.T. Solution volume and seed number: Often overlooked factors in allelopathic bioassays. J. Chem. Ecol. 1987, 13, 1481–1491. [Google Scholar] [CrossRef]
- Jiang, X.; Zhao, Z.; Jacquemyn, H.; Ding, G.; Ding, W.; Xing, X. Addition of fungal inoculum increases seed germination and protocorm formation in a terrestrial orchid. Glob. Ecol. Conserv. 2022, 38, e02235. [Google Scholar] [CrossRef]
- Aldars-García, L.; Marín, S.; Sanchis, V.; Magan, N.; Medina, A. Assessment of intraspecies variability in fungal growth initiation of Aspergillus flavus and aflatoxin B 1 production under static and changing temperature levels using different initial conidial inoculum levels. Int. J. Food Microbiol. 2018, 272, 1–11. [Google Scholar] [CrossRef]
- Krishnan, N.; Velramar, B.; Velu, R.K. Investigation of antifungal activity of surfactin against mycotoxigenic phytopathogenic fungus Fusarium moniliforme and its impact in seed germination and mycotoxicosis. Pestic. Biochem. Physiol. 2019, 155, 101–107. [Google Scholar] [CrossRef]








| Water Levels Scaled at 1 mL Intervals | Water Levels Scaled at the TKW % | ||||
|---|---|---|---|---|---|
| 1 Treatment no | 2 Water Amount mL | 1 Treatment no | 3 Proposed % of Water Level | 4 level of Water mL | 5 Rounded Level of Water mL |
| 1 | 0 | 14 | 90% | 0.69 | 0.7 |
| 2 | 1 | 15 | 180% | 1.38 | 1.4 |
| 3 | 2 | 16 | 270% | 2.07 | 2.05 |
| 4 | 3 | 17 | 360% | 2.76 | 2.75 |
| 5 | 4 | 18 | 450% | 3.45 | 3.45 |
| 6 | 5 | 19 | 540% | 4.14 | 4.15 |
| 7 | 6 | 20 | 630% | 4.83 | 4.85 |
| 8 | 7 | 21 | 720% | 5.52 | 5.5 |
| 9 | 8 | 22 | 810% | 6.21 | 6.2 |
| 10 | 9 | 23 | 900% | 6.9 | 6.9 |
| 11 | 10 | 24 | 990% | 7.59 | 7.6 |
| 12 | 11 | 25 | 1080% | 8.28 | 8.25 |
| 13 | 12 | 26 | 1170% | 8.97 | 8.95 |
| 27 | 1260% | 9.66 | 9.65 | ||
| 28 | 1350% | 10.35 | 10.35 | ||
| 29 | 1440% | 11.04 | 11.05 | ||
| 30 | 1530% | 11.73 | 11.75 | ||
| 1 Water mL | 2 Germination % | 3 Radicle cm | 4 Shoot cm | 5 Seedling cm | 6 Radicle DW g | 7 Shoot DW g | 8 Seedling DW g | 9 Corrected DW g |
|---|---|---|---|---|---|---|---|---|
| 0 | 0.0 ± 0.0 f | 0.0 ± 0.0 d | 0.0 ± 0.0 g | 0.0 ± 0.0 e | 0.000 ± 0.00 e | 0.000 ± 0.00 d | 0.000 ± 0.00 d | 0.000 ± 0.00 c |
| 1 | 90.0 ± 12.7 ab | 3.0 ± 0.9 bc | 0.7 ± 0.1 fg | 3.7 ± 0.9 de | 0.066 ± 0.02 abcd | 0.023 ± 0.01 cd | 0.090 ± 0.03 bc | 0.099 ± 0.03 b |
| 2 | 100.0 ± 0.0 a | 4.7 ± 2.9 abc | 3.1 ± 1.2 def | 7.8 ± 4.0 bcd | 0.094 ± 0.03 a | 0.068 ± 0.03 abc | 0.162 ± 0.05 ab | 0.162 ± 0.05 ab |
| 3 | 91.0 ± 5.5 ab | 2.2 ± 1.2 cd | 1.6 ± 1.4 efg | 3.8 ± 2.6 de | 0.038 ± 0.02 cd | 0.077 ± 0.09 ab | 0.115 ± 0.09 abc | 0.130 ± 0.11 b |
| 4 | 97.0 ± 2.7 a | 6.8 ± 3.8 a | 6.1 ± 3.1 abc | 12.9 ± 6.9 ab | 0.091 ± 0.05 ab | 0.081 ± 0.04 ab | 0.171 ± 0.09 a | 0.177 ± 0.09 ab |
| 5 | 84.0 ± 9.6 bc | 4.9 ± 3.3 abc | 5.8 ± 3.7 abcd | 10.7 ± 7.0 abc | 0.070 ± 0.04 abcd | 0.080 ± 0.04 ab | 0.150 ± 0.08 abc | 0.174 ± 0.07 ab |
| 6 | 79.0 ± 8.9 cd | 5.4 ± 3.1 ab | 5.9 ± 3.1 abc | 11.3 ± 6.2 abc | 0.063 ± 0.04 abcd | 0.075 ± 0.05 ab | 0.138 ± 0.09 abc | 0.173 ± 0.11 ab |
| 7 | 83.0 ± 12.0 bc | 6.8 ± 2.8 a | 7.3 ± 2.3 ab | 14.1 ± 5.0 a | 0.072 ± 0.04 abc | 0.077 ± 0.03 ab | 0.149 ± 0.07 abc | 0.175 ± 0.06 ab |
| 8 | 90.0 ± 7.9 ab | 5.2 ± 2.6 abc | 5.2 ± 2.7 abcd | 10.4 ± 5.3 abc | 0.059 ± 0.03 bcd | 0.064 ± 0.04 abc | 0.123 ± 0.07 abc | 0.137 ± 0.07 b |
| 9 | 77.0 ± 4.5 cd | 2.9 ± 1.2 bcd | 3.6 ± 2.2 cde | 6.5 ± 3.3 cd | 0.037 ± 0.01 d | 0.037 ± 0.02 bcd | 0.074 ± 0.04 cd | 0.096 ± 0.05 b |
| 10 | 66.0 ± 13.4 e | 6.3 ± 1.8 a | 7.9 ± 1.9 a | 14.1 ± 3.6 a | 0.067 ± 0.02 abcd | 0.095 ± 0.03 a | 0.162 ± 0.05 ab | 0.244 ± 0.04 a |
| 11 | 60.0 ± 6.1 e | 4.1 ± 2.1 abc | 4.8 ± 1.7 bcd | 8.9 ± 3.8 abcd | 0.047 ± 0.02 cd | 0.061 ± 0.02 abc | 0.109 ± 0.04 abc | 0.178 ± 0.05 ab |
| 12 | 69.0 ± 4.2 de | 6.4 ± 0.7 a | 0.0 ± 0.0 g | 14.0 ± 1.3 a | 0.070 ± 0.01 abcd | 0.085 ± 0.01 ab | 0.154 ± 0.01 ab | 0.224 ± 0.02 a |
| L.S.D * | 10.17 | 2.94 | 2.74 | 5.58 | 0.035 | 0.049 | 0.077 | 0.084 |
| 1 Wml | 2 Wof TKW | 3 Germi % | 4 Radicle cm | 5 Shoot cm | 6 Seedling cm | 7 Radicle DW g | 8 Plumule DW g | 9 Seedling DW g | 10 Corrected DW g |
|---|---|---|---|---|---|---|---|---|---|
| 0.7 | 90% | 0 ± 0.0 e | 0.0 ± 0.0 h | 0.0 ± 0.0 i | 0.0 ± 0.0 k | 0.000 ± 0.000 g | 0 ± 0.000 h | 0 ± 0.000 i | 0 ± 0.000 i |
| 1.4 | 180% | 74 ± 8.3 bc | 3.1 ± 41.4 g | 1.0 ± 0.6 hi | 4.1 ± 2.5 jk | 0.051 ± 0.029 def | 0.021 ± 0.012 gh | 0.073 ± 0.041 gh | 0.079 ± 0.044 h |
| 2.05 | 270% | 94 ± 1.3 a | 5.4 ± 6.5 cdefg | 4.2 ± 0.7 fg | 9.5 ± 2.6 efghi | 0.082 ± 0.024 abcd | 0.074 ± 0.020 cd | 0.156 ± 0.044 bcde | 0.166 ± 0.045 efg |
| 2.75 | 3605 | 92 ± 0.5 ab | 4.2 ± 2.7 efg | 3.2 ± 2.1 gh | 7.4 ± 5.2 hij | 0.061 ± 0.042 bcde | 0.042 ± 0.035 fg | 0.104 ± 0.076 efgh | 0.113 ± 0.085 gh |
| 3.45 | 450% | 92 ± 1.3 ab | 6.5 ± 6.7 abcde | 5.7 ± 1.9 cdef | 12.2 ± 3.7 cdefg | 0.088 ± 0.021 abc | 0.076 ± 0.019 cd | 0.164 ± 0.039 abcd | 0.179 ± 0.041 def |
| 4.15 | 540% | 89 ± 1.5 ab | 7.2 ± 7.4 abc | 6.4 ± 1.4 bcdef | 13.6 ± 4.4 bcde | 0.093 ± 0.036 a | 0.081 ± 0.018 cd | 0.174 ± 0.054 abcd | 0.196 ± 0.060 bcde |
| 4.85 | 630% | 79 ± 1.8 abc | 3.9 ± 8.9 fg | 4.3 ± 2.3 efg | 8.2 ± 4.4 ghij | 0.048 ± 0.023 ef | 0.045 ± 0.025 efg | 0.092 ± 0.047 fgh | 0.123 ± 0.074 fgh |
| 5.5 | 720% | 85 ± 2.1 abc | 6.8 ± 10.6 abcd | 6.6 ± 2.0 bcd | 13.4 ± 4.2 bcdef | 0.081 ± 0.023 abcd | 0.080 ± 0.027 cd | 0.161 ± 0.049 abcd | 0.187 ± 0.048 cde |
| 6.2 | 810% | 84 ± 0.8 abc | 9.0 ± 4.2 a | 9.2 ± 1.0 a | 18.2 ± 2.2 a | 0.099 ± 0.007 a | 0.109 ± 0.008 ab | 0.207 ± 0.015 ab | 0.247 ± 0.015 abc |
| 6.9 | 900% | 84 ± 3.3 abc | 6.6± 16.4 abcde | 7.7 ± 1.2 abc | 14.3 ± 1.8 abcd | 0.074± 0.010 abcde | 0.091± 0.011 bcd | 0.165 ± 0.018 abcd | 0.201 ± 0.030 bcde |
| 7.6 | 990% | 81 ± 3.6 abc | 7.9 ± 17.8 ab | 8.5 ± 1.4 ab | 16.4 ± 3.4 abc | 0.082 ± 0.027 abcd | 0.096 ± 0.015 abc | 0.177 ± 0.041 abc | 0.223± 0.044 abcde |
| 8.25 | 1080% | 80 ± 2.9 abc | 7.3 ± 14.6 abc | 8.4 ± 1.5 ab | 15.7 ± 3.2 abcd | 0.090 ± 0.021 ab | 0.111 ± 0.020 ab | 0.201 ± 0.038 ab | 0.255 ± 0.046 ab |
| 8.95 | 1170% | 78 ± 1.1 abc | 4.3 ± 5.7 defg | 4.6 ± 1.8 defg | 8.9 ± 3.0 fghi | 0.059 ± 0.018 cde | 0.070 ± 0.023 cde | 0.129 ± 0.040 cdef | 0.165 ± 0.049 efg |
| 9.65 | 1260% | 75 ± 4.1 abc | 5.7 ± 20.6 bcdef | 6.6 ± 3.1 bcde | 12.2 ± 5.4 cdefg | 0.060 ± 0.025 bcde | 0.065 ± 0.027 def | 0.125 ± 0.044 cdefg | 0.165 ± 0.024 efg |
| 10.35 | 1350% | 82 ± 2.7 abc | 8.2 ± 13.5 ab | 9.8 ± 2.4 a | 18.0 ± 5.3 ab | 0.094 ± 0.037 a | 0.121 ± 0.028 a | 0.215 ± 0.065 a | 0.263 ± 0.072 a |
| 11.05 | 1440% | 52 ± 3.1 d | 4.5 ± 15.7 defg | 6.9 ± 2.8 bc | 11.4± 3.9 defgh | 0.052 ± 0.023 def | 0.069 ± 0.029 def | 0.121 ± 0.051 defg | 0.230 ± 0.054 abcd |
| 11.75 | 15,305 | 69 ± 4.4 cd | 2.8 ± 22.2 g | 3.0 ± 0.9 gh | 5.9 ± 1.5 ij | 0.026 ± 0.004 fg | 0.035 ± 0.015 g | 0.061 ± 0.019 h | 0.095 ± 0.033 h |
| L.S.D * | 19.96 | 2.57 | 2.25 | 4.59 | 0.031 | 0.027 | 0.055 | 0.062 | |
| 1 Seeds no | 2 Inactive Seeds % | 3 Initiated % | 4 Radicle-Bearing Seedlings % | 5 Short Shoot Seedling % | 6 Regular Shoot Seedling % | 7 Aggregated Value % |
|---|---|---|---|---|---|---|
| 10 | 0.110 ± 0.057 a | 0.010 ± 0.032 a | 0.010 ± 0.032 b | 0.110 ± 0.099 a | 0.760 ± 0.040 a | 0.838 ± 0.022 a |
| 15 | 0.133 ± 0.118 a | 0.013 ± 0.028 a | 0.027 ± 0.034 ab | 0.180 ± 0.109 a | 0.660 ± 0.058 a | 0.791 ± 0.042 a |
| 20 | 0.090 ± 0.088 a | 0.035 ± 0.047 a | 0.055 ± 0.060 a | 0.145 ± 0.314 a | 0.675 ± 0.087 a | 0.794 ± 0.038 a |
| 25 | 0.048 ± 0.056 a | 0.028 ± 0.060 a | 0.008 ± 0.017 b | 0.248 ± 0.239 a | 0.608 ± 0.117 a | 0.780 ± 0.076 a |
| L.S.D | 0.075 N.S | 0.040 N.S | 0.035 | 0.191 N.S | 0.232 N.S | 0.140 N.S |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarnawa, Á.; Kende, Z.; Sghaier, A.H.; Kovács, G.P.; Gyuricza, C.; Khaeim, H. Effect of Abiotic Stresses from Drought, Temperature, and Density on Germination and Seedling Growth of Barley (Hordeum vulgare L.). Plants 2023, 12, 1792. https://doi.org/10.3390/plants12091792
Tarnawa Á, Kende Z, Sghaier AH, Kovács GP, Gyuricza C, Khaeim H. Effect of Abiotic Stresses from Drought, Temperature, and Density on Germination and Seedling Growth of Barley (Hordeum vulgare L.). Plants. 2023; 12(9):1792. https://doi.org/10.3390/plants12091792
Chicago/Turabian StyleTarnawa, Ákos, Zoltán Kende, Asma Haj Sghaier, Gergő Péter Kovács, Csaba Gyuricza, and Hussein Khaeim. 2023. "Effect of Abiotic Stresses from Drought, Temperature, and Density on Germination and Seedling Growth of Barley (Hordeum vulgare L.)" Plants 12, no. 9: 1792. https://doi.org/10.3390/plants12091792