
Variation in Seed Dormancy of Chaco Seasonally Dry Forest Species: Effects of Seed Traits and Population Environmental Conditions
 , and
, and
Abstract
:1. Introduction
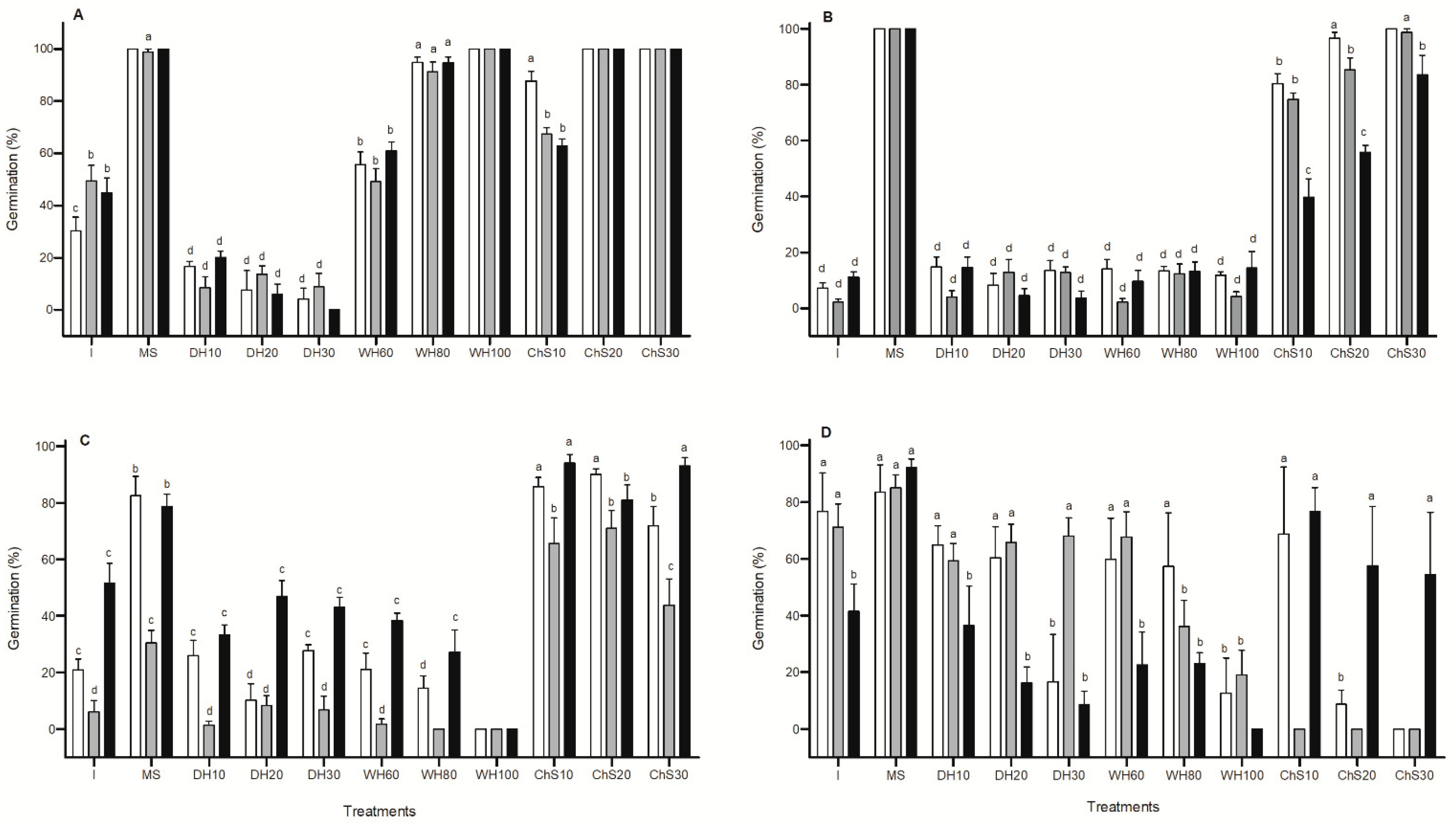
2. Results
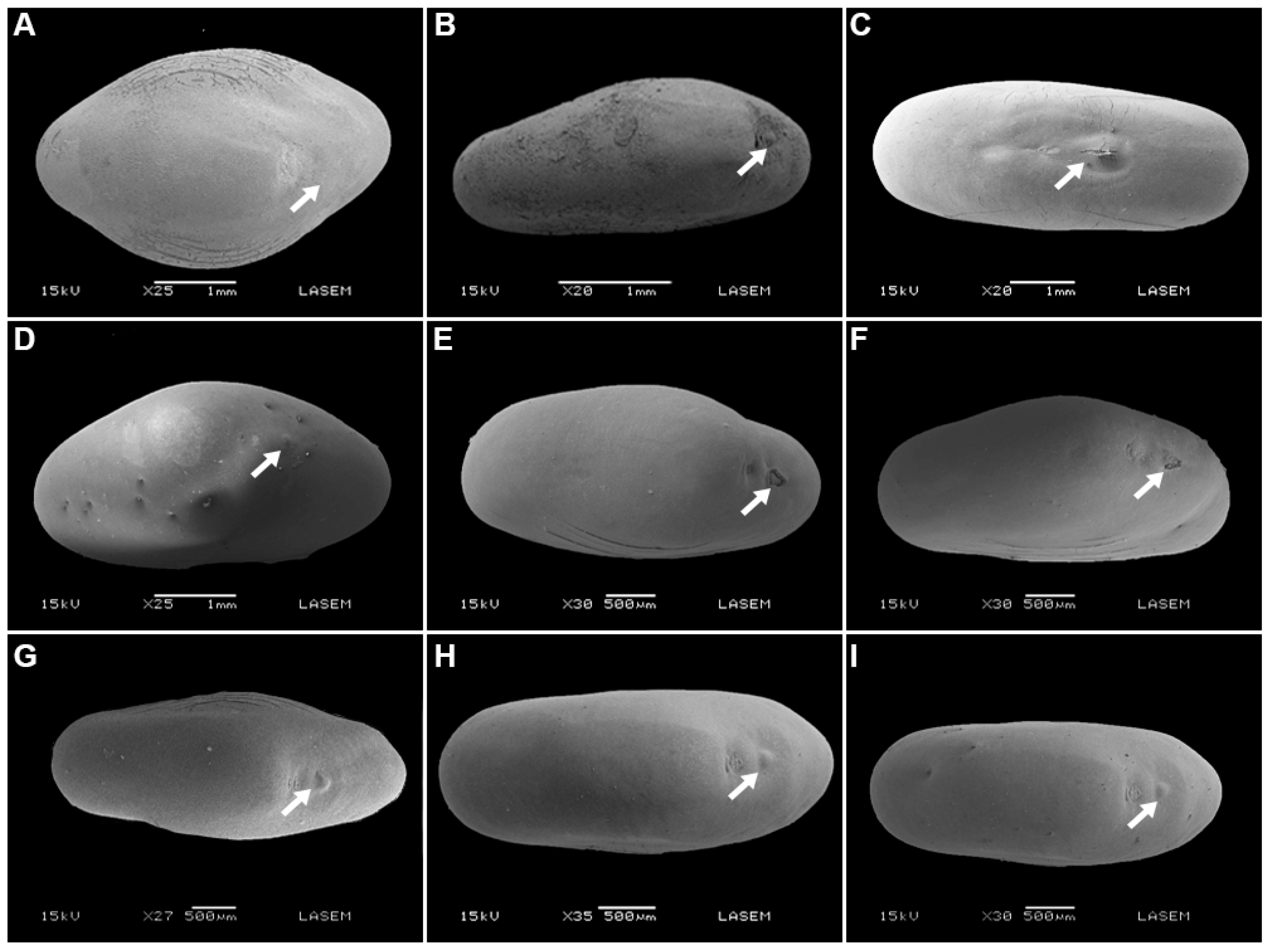
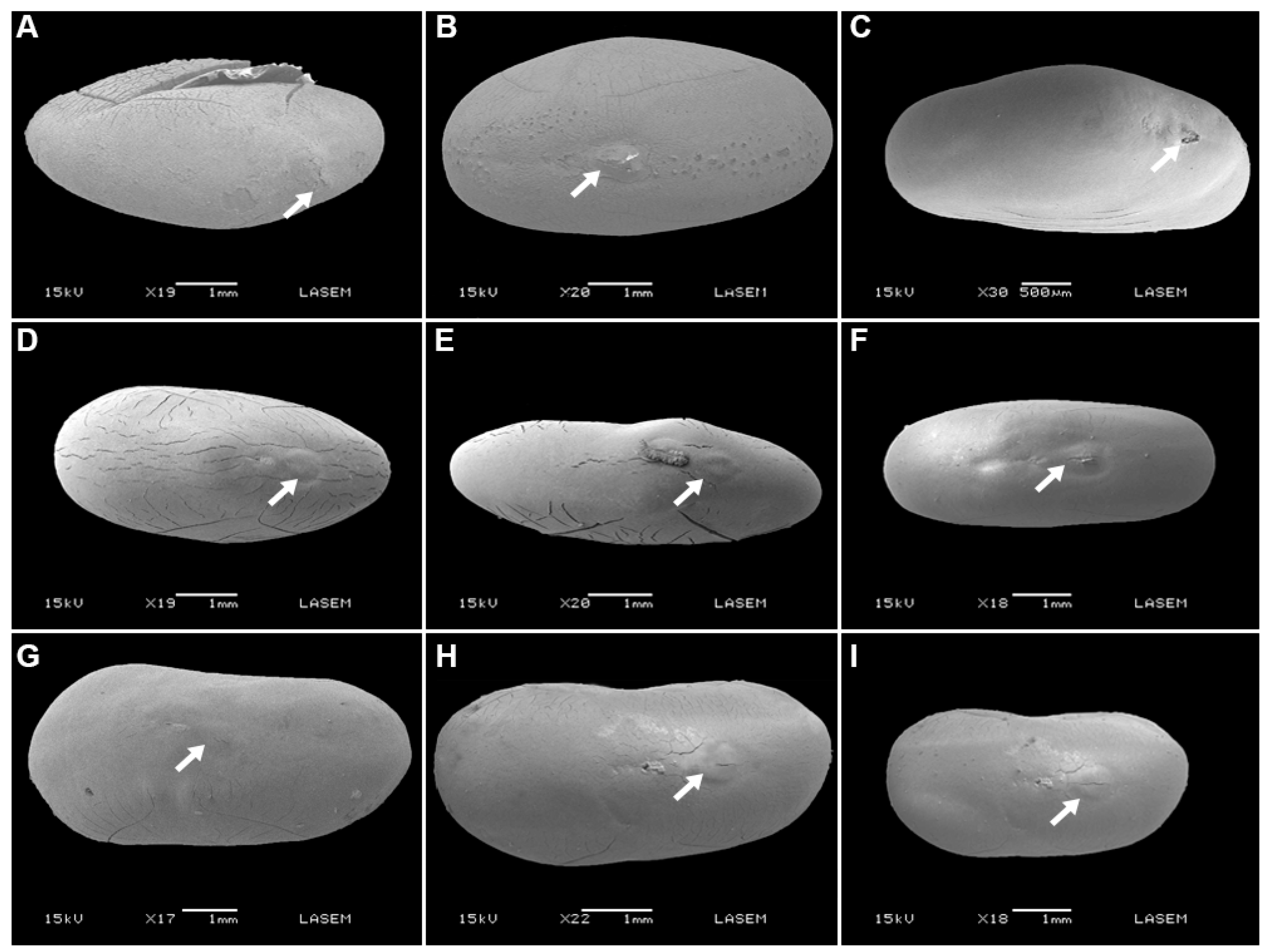
2.1. Seed Traits
2.2. Seed Dormancy
2.2.1. Imbibition Curve
2.2.2. Dormancy-Breaking Treatments
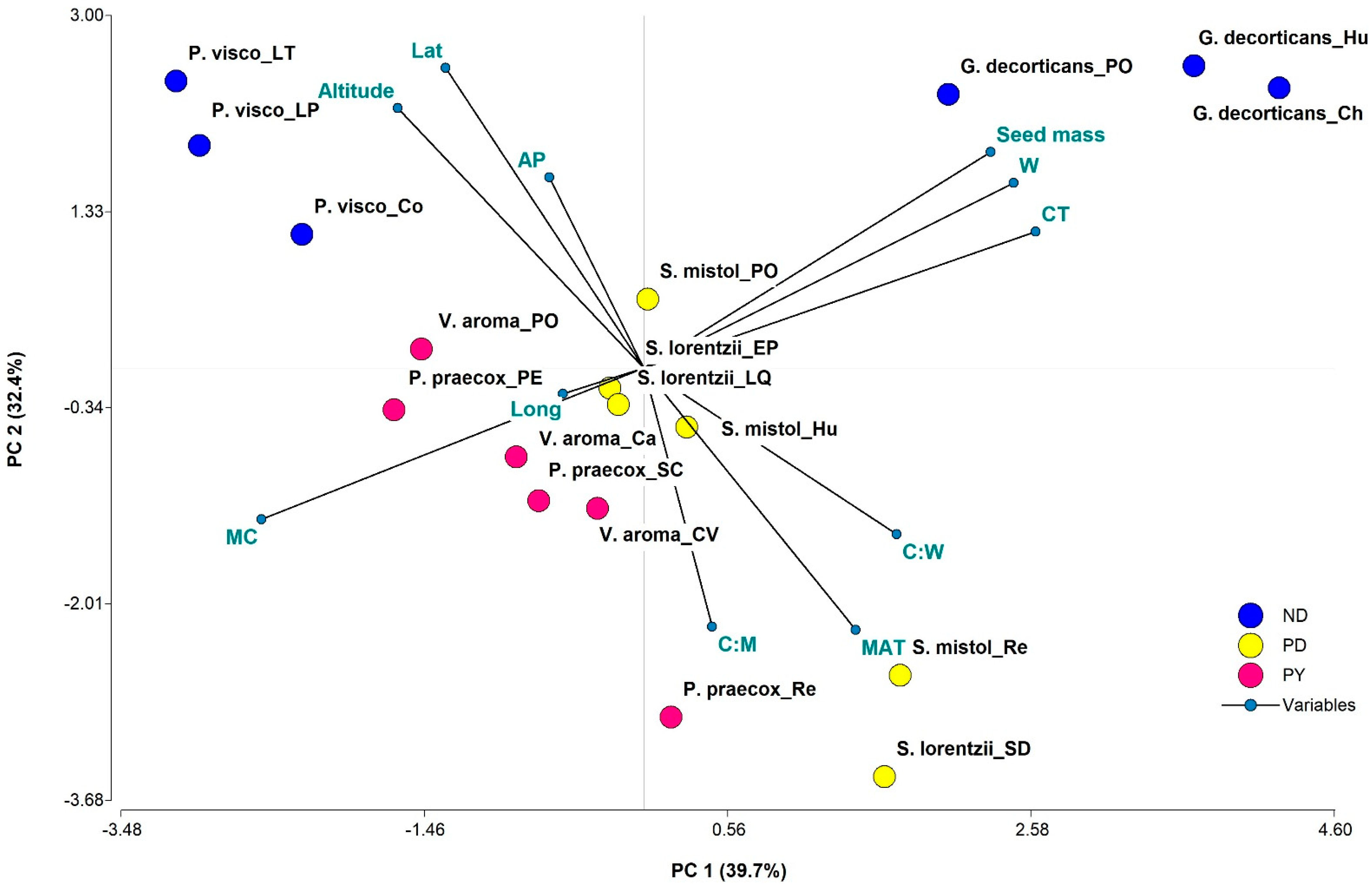
2.3. Associations between MC, Seed Mass, Morpho-Anatomical Traits, and Geographic and Environmental Variables
3. Discussion
3.1. Seed Traits and Seed Dormancy
3.2. Associations between Seed Traits, Seed Dormancy, and Environmental Variables
4. Conclusions
5. Materials and Methods
5.1. Studied Species and Seed Collection
5.2. Seed Traits
5.3. Seed Dormancy
5.3.1. Imbibition Curve
5.3.2. Dormancy-Breaking Treatments
5.4. Statistical Analyses
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vallejos, M.; Volante, J.N.; Mosciaro, M.J.; Vale, L.M.; Bustamante, M.L.; Paruelo, J.M. Transformation dynamics of the natural cover in the Dry Chaco ecoregion: A plot level geo-database from 1976 to 2012. J. Arid Environ. 2014, 123, 3–11. [Google Scholar] [CrossRef]
- Volante, J.N.; Alcaraz-Segura, D.; Mosciaro, M.J.; Viglizzo, E.F.; Paruelo, J.M. Ecosystem functional changes associated with land clearing in NW Argentina. Agric. Ecosyste. Environ. 2012, 154, 12–22. [Google Scholar] [CrossRef]
- Gold, K.; Way, M. Seed Conservation of the Latinamerican flora–An international opportunity. Royal Botanic Gardens. Lyonia 2004, 6, 19–24. [Google Scholar]
- Kameswara Rao, N.K.; Hanson, J.; Dulloo, M.E.; Ghosh, K.; Nowell, A. Manual of Seed Handling in Genebanks; No.8; Bioversity International: Rome, Italy, 2006; p. 147. [Google Scholar]
- Baskin, C.C.; Baskin, J.M. Breaking seed dormancy during dry storage: A useful tool or major problem for successful restoration via direct seeding? Plants 2020, 9, 636. [Google Scholar] [CrossRef] [PubMed]
- Kildisheva, O.A.; Dixon, K.W.; Silveira, F.A.; Chapman, T.; Di Sacco, A.; Mondoni, A.; Cross, A.T. Dormancy and germination: Making every seed count in restoration. Restor. Ecol. 2020, 28, S256–S265. [Google Scholar] [CrossRef]
- Seglias, A.E.; Williams, E.; Bilge, A.; Kramer, A.T. Phylogeny and source climate impact seed dormancy and germination of restoration relevant forb species. PLoS ONE 2018, 13, e0191931. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Elsevier/Academic Press: San Diego, CA, USA, 2014; pp. 145–181. [Google Scholar]
- Baskin, J.M.; Baskin, C.C. Classification, biogeography, and phylogenetic relationships of seed dormancy. In Seed Conservation: Turning Science into Practice; Royal Botanic Gardens, Kew: Richmond, UK, 2003; pp. 518–544. [Google Scholar]
- Baskin, J.M.; Baskin, C.C. A classification system for seed dormancy. Seed Sci. Res. 2004, 14, 1–16. [Google Scholar] [CrossRef]
- Jaganathan, G.K.; Song, D.; Liu, B. Diversity and distribution of physical dormant species in relation to ecosystem and life-forms. Plant Sci. Today 2017, 4, 55–63. [Google Scholar] [CrossRef]
- Rubio de Casas, R.; Willis, C.G.; Pearse, W.D.; Baskin, C.C.; Baskin, J.M.; Cavender-Bares, J. Global biogeography of seed dormancy is determined by seasonality and seed size: A case study in the legumes. New Phytol. 2017, 214, 1527–1536. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C.; Li, X. Taxonomy, anatomy and evolution of physical dormancy in seeds. Plant Species Biol. 2000, 15, 139–152. [Google Scholar] [CrossRef]
- Gama-Arachchige, N.S.; Baskin, J.M.; Geneve, R.L.; Baskin, C.C. Identification and characterization of ten new water gaps in seeds and fruits with physical dormancy and classification of water-gap complexes. Ann. Bot. 2013, 112, 69–84. [Google Scholar] [CrossRef]
- Li, X.; Baskin, J.M.; Baskin, C.C. Anatomy of two mechanisms of breaking physical dormancy by experimental treatments in seeds of two North American Rhus species (Anacardiaceae). Am. J. Bot. 1999, 86, 1505–1511. [Google Scholar] [CrossRef]
- Galíndez, G.; Ortega-Baes, P.; Seal, C.E.; Daws, M.I.; Scopel, A.L.; Pritchard, H.W. Physical seed dormancy in Collaea argentina (Fabaceae) and Abutilon pauciflorum (Malvaceae) after 4 years storage. Seed Sci. Technol. 2010, 38, 777–782. [Google Scholar] [CrossRef]
- Galíndez, G.; Ceccato, D.; Malagrina, G.; Pidal, B.; Chilo, G.; Bach, H.; Ortega-Baes, P. Physical seed dormancy in native legume species of Argentina. Bol. Soc. Argent. Bot. 2016, 51, 73–78. [Google Scholar] [CrossRef]
- Jaganathan, G.K. Influence of maternal environment in developing different levels of physical dormancy and its ecological significance. Plant Ecol. 2015, 217, 71–79. [Google Scholar] [CrossRef]
- Rodrigues-Junior, A.G.; Mello, A.C.M.; Baskin, C.C.; Baskin, J.M.; Oliveira, D.M.; Garcia, Q.S. Why large seeds with physical dormancy become nondormant earlier than small ones. PLoS ONE 2018, 13, e0202038. [Google Scholar] [CrossRef]
- Donohue, K.; Dorn, L.; Griffith, C.; Kim, E.; Aguilera, A.; Polisetty, C.R.; Schmitt, J. Environmental and genetic influences on the germination of Arabidopsis thallana in the field. Evolution 2005, 59, 740–757. [Google Scholar] [CrossRef] [PubMed]
- Renzi, J.P.; Duchoslav, M.; Brus, J.; Hradilová, I.; Pechanec, V.; Václavek, T.; Machalová, J.; Hron, K.; Verdier, J.; Smýkal, P. Physical dormancy release in Medicago truncatula seeds is related to environmental variations. Plants 2020, 9, 503. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Pascual, E.; Jiménez-Alfaro, B.; Caujapé-Castells, J.; Jaén-Molina, R.; Díaz, T.E. A local dormancy cline is related to the seed maturation environment, population genetic composition and climate. Ann. Bot. 2013, 112, 937–945. [Google Scholar] [CrossRef] [PubMed]
- Hay, F.R.; Smith, R.D.; Ellis, R.H.; Butler, L. Developmental changes in the germinability, desiccation tolerance, hardseededness, and longevity of individual seeds of Trifolium ambiguum. Ann. Bot. 2010, 105, 1035–1052. [Google Scholar] [CrossRef]
- Oyarzabal, M.; Clavijo, J.; Oakley, L.; Biganzoli, F.; Tognetti, P.; Barberis, I.; León, R.J. Unidades de vegetación de la Argentina. Ecol. Austral 2018, 28, 40–63. [Google Scholar] [CrossRef]
- Karlin, M.; Karlin, U.O.; Coirini, R.; Contreras, A. Cambio climático en el Chaco Seco. In Proceedings of the Congreso Internacional de Cambio Climático; Publicaciones CCC: Córdoba, Argentina, 2019. [Google Scholar]
- Morláns, M.C. Regiones naturales de Catamarca: Provincias geológicas y provincias fitogeográficas. Rev. Cienc. Téc. 1995, 2, 1–42. [Google Scholar]
- Renison, D.; Valladares, G.; Martella, M.B. The effect of passage through the gut of the Greater Rhea (Rhea americana) on germination of tree seeds: Implications for forest restoration. Emu-Aust. Ornithol. 2010, 110, 125–131. [Google Scholar] [CrossRef]
- Funes, G.; Díaz, S.; Venier, P. La temperatura como principal determinante de la germinación en especies del Chaco seco de Argentina. Ecol Austral. 2009, 19, 129–138. [Google Scholar]
- Paredes, D.A.; Rodriguez Araujo, M.E.; Pérez, D.R. Germinación de tres especies de Fabaceae de interés para la restauración ecológica en el Monte Austral, Patagonia, Argentina. Quebracho 2018, 26, 68–78. [Google Scholar]
- Rodriguez Araujo, M.E.; Pérez, D.; Aronson, J.; Cross, A. Filling gaps on seed germination and species selection in drylands of Argentina: Work in progress and reflections on intelligent tinkering. Multequina 2021, 30, 165–180. [Google Scholar]
- Araoz, S.D.; Del Longo, O.T. Tratamientos pregerminativos para romper la dormición física impuesta por el endocarpo en Ziziphus mistol Grisebach. Quebracho 2006, 13, 56–65. [Google Scholar]
- Ibañez Moro, A.V.; Bravo, S.J.; Abdala, N.R.; Borghetti, F.; Chaib, A.M.; Galetto, L. Heat shock effects on germination and seed survival of five woody species from the Chaco region. Flora 2021, 275, 151751. [Google Scholar] [CrossRef]
- Venier, P.; Funes, G.; García, C.C. Physical dormancy and histological features of seeds of five Acacia species (Fabaceae) from xerophytic forests in central Argentina. Flora 2012, 207, 39–46. [Google Scholar] [CrossRef]
- Ferreras, A.E.; Zeballos, S.R.; Funes, G. Inter-and intra-population variability in physical dormancy along a precipitation gradient. Acta Bot Bras. 2017, 31, 141–146. [Google Scholar] [CrossRef]
- Jaureguiberry, P.; Díaz, S. Post-burning regeneration of the Chaco seasonally dry forest: Germination response of dominant species to experimental heat shock. Oecologia 2015, 177, 689–699. [Google Scholar] [CrossRef]
- Del Longo, O.T.; Aráoz, S.D. Criteria to determine the duration of the germination test for seeds of mistol (Ziziphus mistol Grisebach). Seed Sci. Technol. 2009, 37, 507–511. [Google Scholar] [CrossRef]
- Baskin, J.M.; Baskin, C.C. Seed (true seed plus endocarp) dormancy in Anacardiaceae in relation to infrafamilial taxonomy and endocarp anatomy. Seed Sci. Res. 2022, 32, 187–199. [Google Scholar] [CrossRef]
- Bertuzzi, T.; Pastrana-Ignes, V.; Curti, R.N.; Batlla, D.; Baskin, C.C.; Sühring, S.; Galíndez, G. Variation in thermal and hydrotime requirements for seed germination of Chaco seasonally dry forest species in relation to population environmental conditions and seed mass. Austral Ecol. 2022, 47, 1232–1247. [Google Scholar] [CrossRef]
- Saatkamp, A.; Cochrane, A.; Commander, L.; Guja, L.K.; Jimenez-Alfaro, B.; Larson, J.; Nicotra, A.; Poschlod, P.; Silveira, F.A.O.; Cross, A.T.; et al. A research agenda for seed-trait functional ecology. New Phytol. 2018, 221, 1764–1775. [Google Scholar] [CrossRef]
- Funes, G.; Venier, P. Dormancy and germination in three Acacia (Fabaceae) species from central Argentina. Seed Sci. Res. 2006, 16, 77–82. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Mimicking the natural thermal environments experienced by seeds to break physiological dormancy to enhance seed testing and seedling production. Seed Sci. Technol. 2022, 50, 21–29. [Google Scholar] [CrossRef]
- Ooi, M.K.; Auld, T.D.; Denham, A.J. Projected soil temperature increase and seed dormancy response along an altitudinal gradient: Implications for seed bank persistence under climate change. Plant Soil 2012, 353, 289–303. [Google Scholar] [CrossRef]
- Cochrane, J.A.; Hoyle, G.L.; Yates, C.J.; Wood, J.; Nicotra, A.B. Evidence of population variation in drought tolerance during seed germination in four Banksia (Proteaceae) species from Western Australia. Aust. J. Bot. 2014, 62, 481–489. [Google Scholar] [CrossRef]
- Hradilová, I.; Duchoslav, M.; Brus, J.; Pechanec, V.; Hýbl, M.; Kopecký, P.; Smýkal, P. Variation in wild pea (Pisum sativum subsp. elatius) seed dormancy and its relationship to the environment and seed coat traits. PeerJ 2019, 7, e6263. [Google Scholar] [CrossRef]
- Klupczyńska, E.A.; Pawłowski, T.A. Regulation of seed dormancy and germination mechanisms in a changing environment. Int. J. Mol. Sci. 2021, 22, 1357. [Google Scholar] [CrossRef] [PubMed]
- Berger, J.D.; Shrestha, D.; Ludwig, C. Reproductive strategies in Mediterranean legumes: Trade-offs between phenology, seed size and vigor within and between wild and domesticated Lupinus species collected along aridity gradients. Front. Plant Sci. 2017, 8, 548. [Google Scholar] [CrossRef] [PubMed]
- Demaio, P.; Karlin, U.O.; Medina, M. Árboles Nativos de Argentina. Tomo 3: Noroeste; Ecoval Editorial: Córdoba, Argentina, 2015; p. 410. [Google Scholar]
- International Seed Testing Association-ISTA. International Rules for Seed Testing; The International Seed Testing Association: Bassersdorf, Switzerland, 2017. [Google Scholar]
- Rasband, W.S.; ImageJ, U.S. National Institutes of Health, Bethesda, Maryland, USA, 1997–2018. Available online: https://imagej.nih.gov/ij/ (accessed on 1 November 2022).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
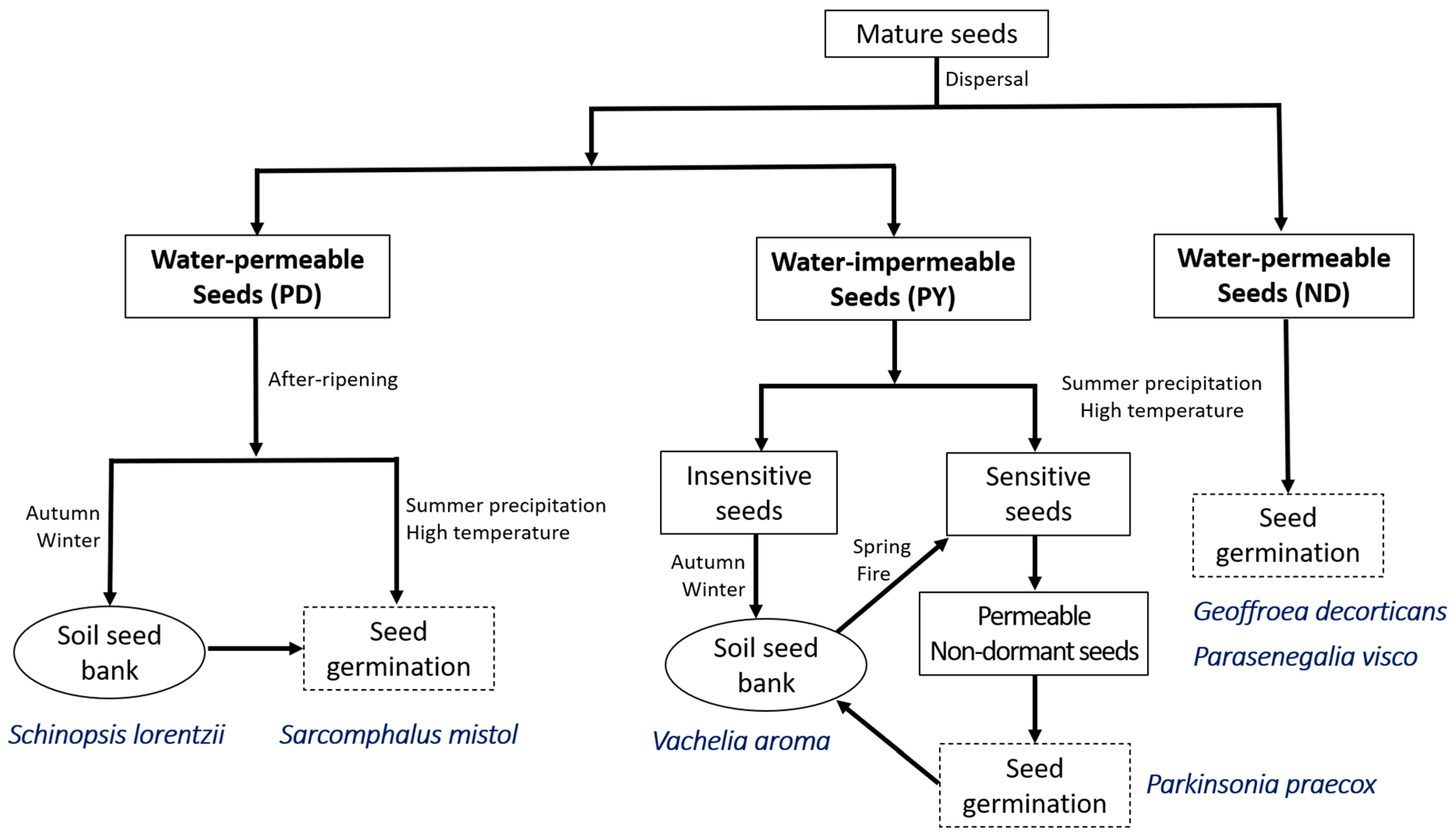{kind=link}
| Species | Population | MC (%) | Seed Mass (mg) | Germination (%) | Water Uptake (%) | ||||
|---|---|---|---|---|---|---|---|---|---|
| Light | Darkness | Intact | Scarified | ||||||
| 25 °C | 30/20 °C | 25 °C | 30/20 °C | ||||||
| Geoffroea decorticans | Chacabuco | 5.4 (0.3) Ca | 1005.2 (355) Aa | 16 (9) Cc | 7 (8) Cb | 83 (17) Ca | 77 (32) Ca | 71.2 (30) Ba | 63.9 (12) Ba |
| Huaycama | 5.7 (0.5) Ca | 909.3 (115) Aa | 12 (5) Bc | 74 (28) Bb | 78 (19) Ba | 61 (41) Ba | 76.5 (37) Ba | 63.3 (11) Ba | |
| Pomancillo Oeste | 6.3 (0.1) Ca | 700.9 (246) Ab | 36 (18) Ac | 52 (31) Ab | 98 (5) Aa | 100 | 85.7 (16) Aa | 80.4 (19) Aa | |
| Parasenegalia visco | Concepción | 8.9 (0.2) Ab | 111.3 (18) Ca | 100 | 100 | 100 | 100 | 233.4 (32) Ba | 223.0 (54) Aa |
| La Puerta | 9.5 (0.04) Aa | 99.0 (15) Cb | 100 | 99 (3) | 100 | 100 | 195.6 (193) Ca | 252.6 (45) Aa | |
| Los Túneles | 9.4 (0.3) Aa | 101.8 (18) Cb | 100 | 99 (3) | 100 | 100 | 276.4 (46) Aa | 232.1 (38) Ab | |
| Parkinsonia praecox | Pomancillo Este | 8.7 (0.2) Aa | 46.5 (5) Da | 44 (6) Aa | 50 (20) Aa | 38 (8) Ab | 26 (7) Ab | 64.2 (77) Ab | 150.2 (17) Aa |
| Recreo | 9.0 (0.2) Aa | 42.5 (7) Da | 35 (10) Aa | 42 (14) Aa | 32 (17) Ab | 34 (15) Ab | 56.8 (86) Ab | 150.3 (36) Aa | |
| Santa Cruz | 8.7 (0.1) Aa | 46.2 (9) Da | 47 (5) Aa | 46 (15) Aa | 34 (6) Ab | 24 (18) Ab | 46.6 (72) Ab | 144.1 (39) Aa | |
| Sarcomphalus mistol | Huaycama | 8.4 (0.1) Ba | 234.4 (38) Ba | 48 (7) Aa | 48 (8) Aa | 13 (18) Ab | 12 (13) Ab | 28.5 (5) Aa | 29.1 (3) Aa |
| Pomancillo Oeste | 7.6 (0.1) Bc | 233.7 (37) Ba | 4 (4) Ca | 2(3) Ca | 0 | 0 | 19.1 (3) Ba | 19.7 (9) Ba | |
| Recreo | 7.8 (0.05) Bb | 176.9 (31) Bb | 28 (18) Ba | 29 (18) Ba | 5 (6) Bb | 8 (7) Bb | 26.5 (3) Aa | 29.7 (6) Aa | |
| Schinopsis lorentzii | El Portezuelo | 8.6 (0.1) Aa | 118.8 (24) Ca | 37 (26) Ba | 40 (33) Bb | 26 (13) Ba | 25 (24) Bb | 76.5 (17) Aa | 67.9 (11) Ab |
| La Quebrada | 8.5 (0.1) Aa | 128.2 (30) Ca | 55 (13) Aa | 41 (9) Ab | 43 (7) Aa | 39 (33) Ab | 78.2 (23) Aa | 77.6 (18) Aa | |
| Santo Domingo | 8.7 (0.1) Aa | 117.1 (27) Ca | 24 (13) Ca | 10 (4) Cb | 15 (15) Ca | 9 (4) Cb | 61.4 (9) Ba | 49.6 (9) Bb | |
| Vachellia | Capital | 8.8 (0.1) Aa | 61.2 (7) Da | 8 (1) Ba | 4 (5) Ba | 8 (7) Ba | 4 (5) Ba | 18.0 (47) Ba | 152.8 (20) Ab |
| aroma | Colonia del Valle | 8.2 (0.2) Ab | 83.6 (10) Db | 10 (3) Ba | 9 (3) Ba | 8 (7) Ba | 5 (4) Ba | 2.9 (5) Ba | 167.0 (49) Ab |
| Pomancillo Oeste | 9.3 (0.4) Aa | 66.7 (17) Da | 14 (6) Aa | 14 (4) Aa | 15 (18) Aa | 6 (8) Aa | 53.1 (82) Aa | 124.2 (22) Bb |
| Species | Population | Pl (mm) | CT (mm) | W (mm) | C:W | C:M |
|---|---|---|---|---|---|---|
| Geoffroea decorticans | Chacabuco | - | 4.21 (0.6) Aa | 14.69 (2.8) Aa | 0.29 (0.02) Ca | 0.004 (0.0003) Da |
| Huaycama | - | 3.87 (0.1) Aa | 13.15 (0.6) Aa | 0.29 (0.01) Ca | 0.004 (0.001) Da | |
| Pomancillo Oeste | - | 2.49 (0.5) Ab | 10.09 (1.5) Aa | 0.24 (0.01) Cb | 0.003 (0.001) Da | |
| Parasenegalia visco | Concepción | 0.036 (0.01) Ca | 0.14 (0.02) Fa | 1.54 (0.3) Da | 0.09 (0.01) Eb | 0.001(0.0003) Ea |
| La Puerta | 0.029 (0.005) Ca | 0.11 (0.03) Fa | 1.46 (0.3) Da | 0.08 (0.01) Eb | 0.001 (0.0003) Ea | |
| Los Túneles | 0.030 (0.004) Ca | 0.14 (0.02) Fa | 1.19 (0.1) Da | 0.12 (0.03) Ea | 0.001 (0.0002) Ea | |
| Parkinsonia praecox | Pomancillo Este | 0.063 (0.005) Ba | 0.32 (0.04) Ea | 1.47 (0.3) Da | 0.23 (0.05) Cb | 0.007 (0.001) Ba |
| Recreo | 0.061 (0.003) Ba | 0.34 (0.05) Ea | 1.25 (0.2) Da | 0.28 (0.02) Ca | 0.008 (0.002) Ba | |
| Santa Cruz | 0.063 (0.01) Ba | 0.40 (0.07) Ea | 1.48 (0.2) Da | 0.27 (0.01) Ca | 0.009 (0.002) Ba | |
| Sarcomphalus mistol | Huaycama | - | 1.15 (0.03) Ca | 3.72 (0.05) Ba | 0.31 (0.01) Bb | 0.005 (0.001) Ca |
| Pomancillo Oeste | - | 1.13 (0.1) Ca | 3.32 (0.3) Ba | 0.34 (0.02) Ba | 0.004 (0.001) Ca | |
| Recreo | - | 1.29 (0.009) Ca | 3.62 (0.1) Ba | 0.36 (0.01) Ba | 0.007 (0.0003) Ca | |
| Schinopsis lorentzii | El Portezuelo | - | 1.59 (0.4) Ba | 3.99 (0.9) Ba | 0.40 (0.03) Aa | 0.014 (0.004) Aa |
| La Quebrada | - | 1.4 (0.3) Ba | 3.54 (0.6) Ba | 0.40 (0.02) Aa | 0.010 (0.003) Aa | |
| Santo Domingo | - | 1.46 (0.2) Ba | 3.52 (0.5) Ba | 0.42 (0.03) Aa | 0.013 (0.003) Aa | |
| Vachellia aroma | Capital | 0.153 (0.03) Aa | 0.47 (0.1) Da | 2.62 (0.4) Ca | 0.18 (0.03) Da | 0.008 (0.003) Ba |
| Colonia del Valle | 0.097 (0.02) Ab | 0.44 (0.09) Da | 2.55 (0.4) Ca | 0.17 (0.05) Da | 0.004 (0.0005) Ba | |
| Pomancillo Oeste | 0.106 (0.01) Ab | 0.46 (0.04) Da | 2.77 (0.4) Ca | 0.17 (0.01) Da | 0.007 (0.002) Ba |
| Species (Family) | Population | Vegetation Unit | Latitude | Longitude | Altitude (m a.s.l.) | MAT (°C) | AP (mm) |
|---|---|---|---|---|---|---|---|
| Geoffroea decorticans (Fabaceae) | Chacabuco | Arid Chaco | −28°26′ | −65°43′ | 525 | 20.1 | 529 |
| Huaycama | Arid Chaco | −28°30′ | −65°39′ | 578 | 19.0 | 529 | |
| Pomancillo Oeste | Chaco Serrano | −28°18′ | −65°43′ | 648 | 19.4 | 545 | |
| Parasenegalia visco (Fabaceae) | Concepción | Chaco Serrano | −28°40′ | −66°04′ | 881 | 18.7 | 441 |
| La Puerta | Chaco Serrano | −28°11′ | −65°46′ | 880 | 18.2 | 547 | |
| Los Túneles | Chaco Serrano | −28°08′ | −65°38′ | 955 | 18.3 | 613 | |
| Parkinsonia praecox (Fabaceae) | Pomancillo Este | Chaco Serrano | −28°18′ | −65°42′ | 677 | 19.4 | 545 |
| Recreo | Semiarid Chaco | −29°05′ | −65°4′ | 243 | 20.5 | 491 | |
| Santa Cruz | Arid Chaco | −28°29′ | −65°40′ | 566 | 20.1 | 530 | |
| Sarcomphalus mistol (Rhamnaceae) | Huaycama | Arid Chaco | −28°31′ | −65°41′ | 538 | 20.1 | 524 |
| Pomancillo Oeste | Chaco Serrano | −28°18′ | −65°43′ | 646 | 19.4 | 545 | |
| Recreo | Semiarid Chaco | −29°08′ | −65°05′ | 235 | 20.6 | 483 | |
| Schinopsis lorentzii (Anacardiaceae) | El Portezuelo | Chaco Serrano | −28°27′ | −65°36′ | 760 | 18.5 | 553 |
| La Quebrada | Chaco Serrano | −28°27′ | −65°51′ | 887 | 19.2 | 502 | |
| Santo Domingo | Semiarid Chaco | −29°09′ | −65°12′ | 254 | 20.5 | 483 | |
| Vachellia aroma (Fabaceae) | Capital | Arid Chaco | −28°26′ | −65°45′ | 539 | 20.1 | 529 |
| Colonia del Valle | Arid Chaco | −28°38′ | −65°54′ | 476 | 20.2 | 494 | |
| Pomancillo Oeste | Chaco Serrano | −28°18′ | −65°43′ | 653 | 19.4 | 545 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertuzzi, T.; López-Spahr, D.; Gómez, C.A.; Sühring, S.; Malagrina, G.; Baskin, C.C.; Galíndez, G. Variation in Seed Dormancy of Chaco Seasonally Dry Forest Species: Effects of Seed Traits and Population Environmental Conditions. Plants 2023, 12, 1790. https://doi.org/10.3390/plants12091790
Bertuzzi T, López-Spahr D, Gómez CA, Sühring S, Malagrina G, Baskin CC, Galíndez G. Variation in Seed Dormancy of Chaco Seasonally Dry Forest Species: Effects of Seed Traits and Population Environmental Conditions. Plants. 2023; 12(9):1790. https://doi.org/10.3390/plants12091790
Chicago/Turabian StyleBertuzzi, Tania, Diego López-Spahr, Carlos A. Gómez, Silvia Sühring, Gisela Malagrina, Carol C. Baskin, and Guadalupe Galíndez. 2023. "Variation in Seed Dormancy of Chaco Seasonally Dry Forest Species: Effects of Seed Traits and Population Environmental Conditions" Plants 12, no. 9: 1790. https://doi.org/10.3390/plants12091790