
Extracts and Residues of Common Ragweed (Ambrosia artemisiifolia L.) Cause Alterations in Root and Shoot Growth of Crops
 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Chemical Compounds of Common Ragweed
2.2. Laboratory Experiment: Ragweed Had Significant Impact on Root Growth
2.2.1. Germination
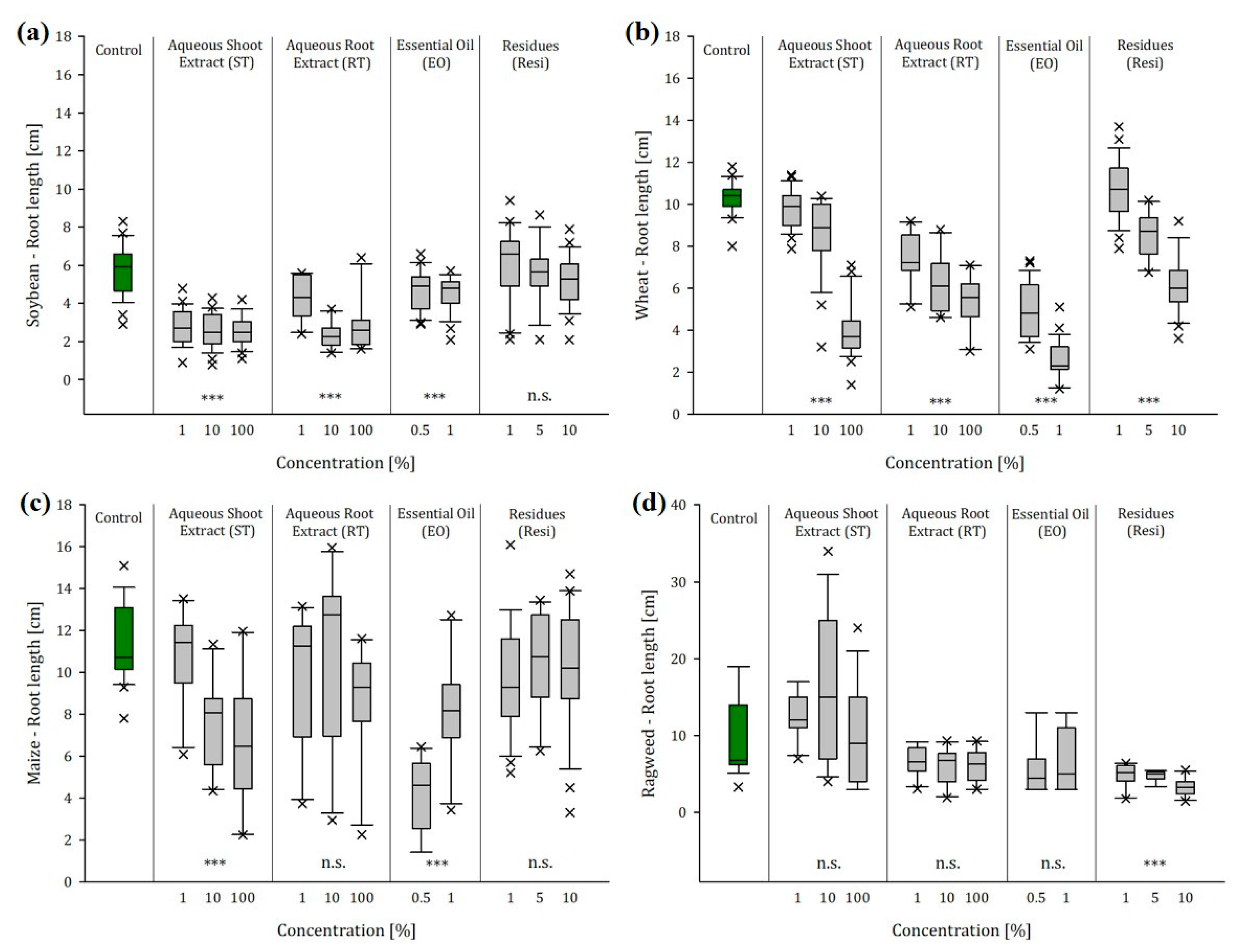
2.2.2. Root Length
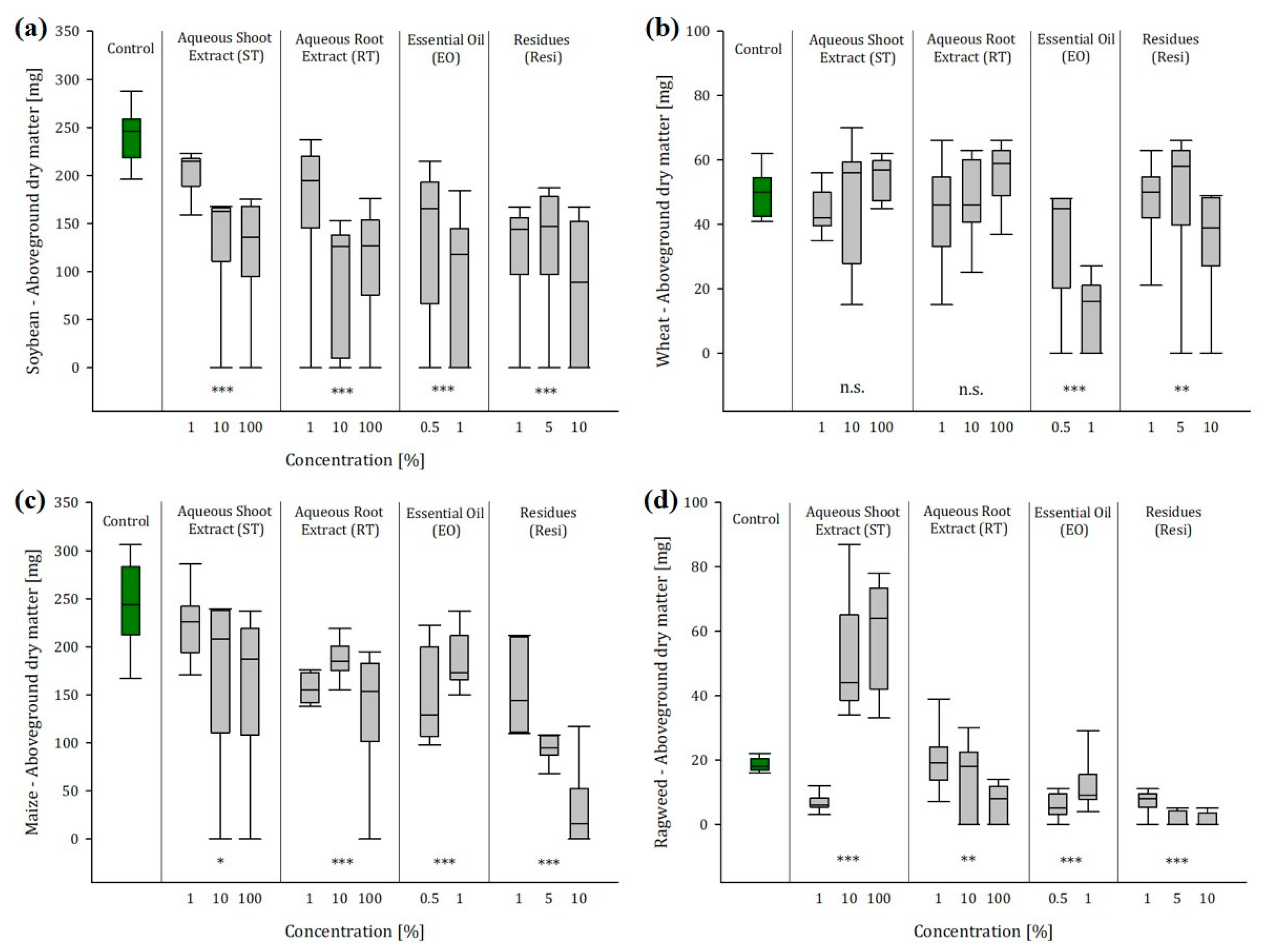
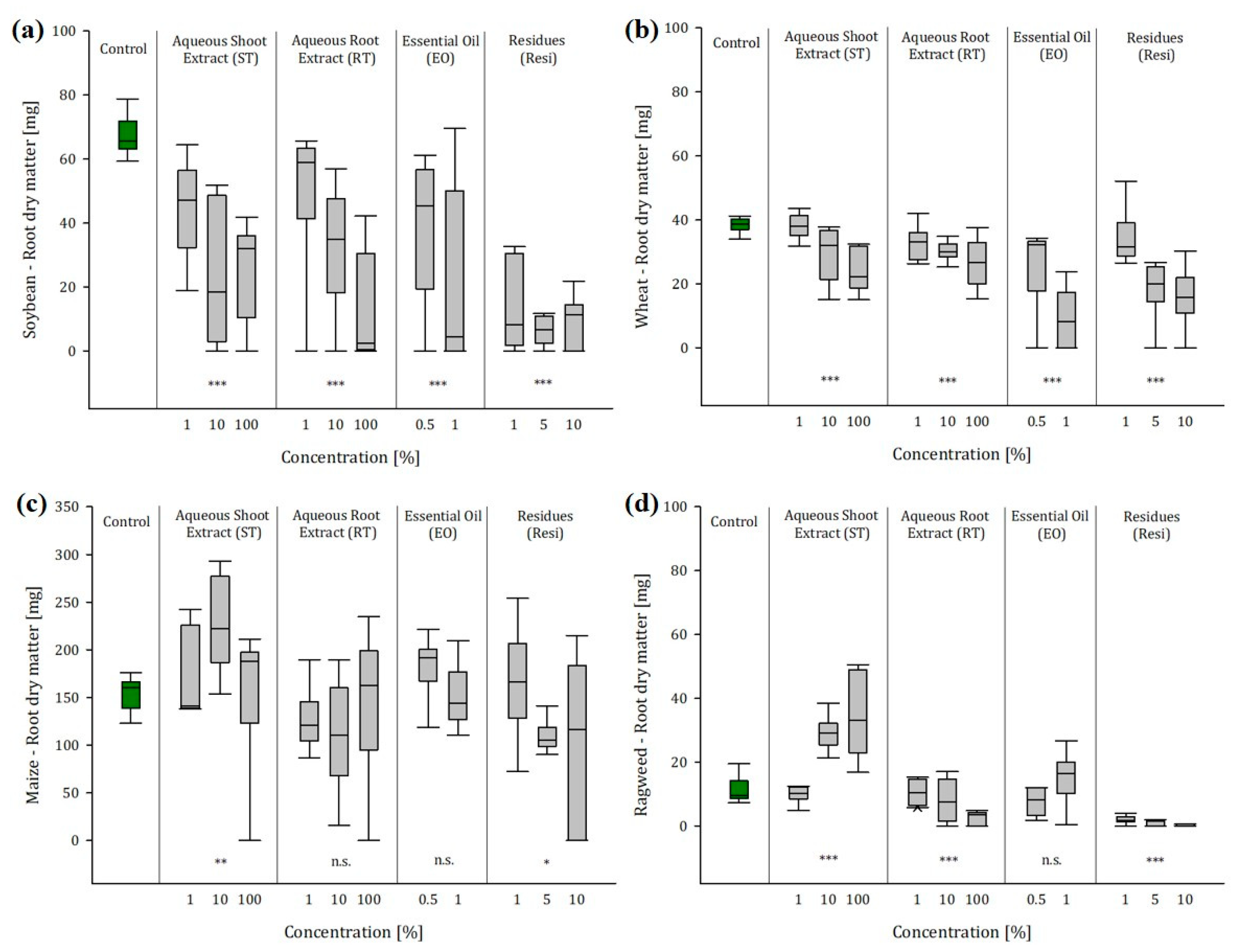
2.3. Greenhouse Experiment 1: Soil Conditions Enhanced the Effects of Ragweed Extracts and Residues
2.3.1. Ragweed Shoot Extract
2.3.2. Ragweed Shoot Extract
2.3.3. Ragweed Essential Oil
2.3.4. Ragweed Residues
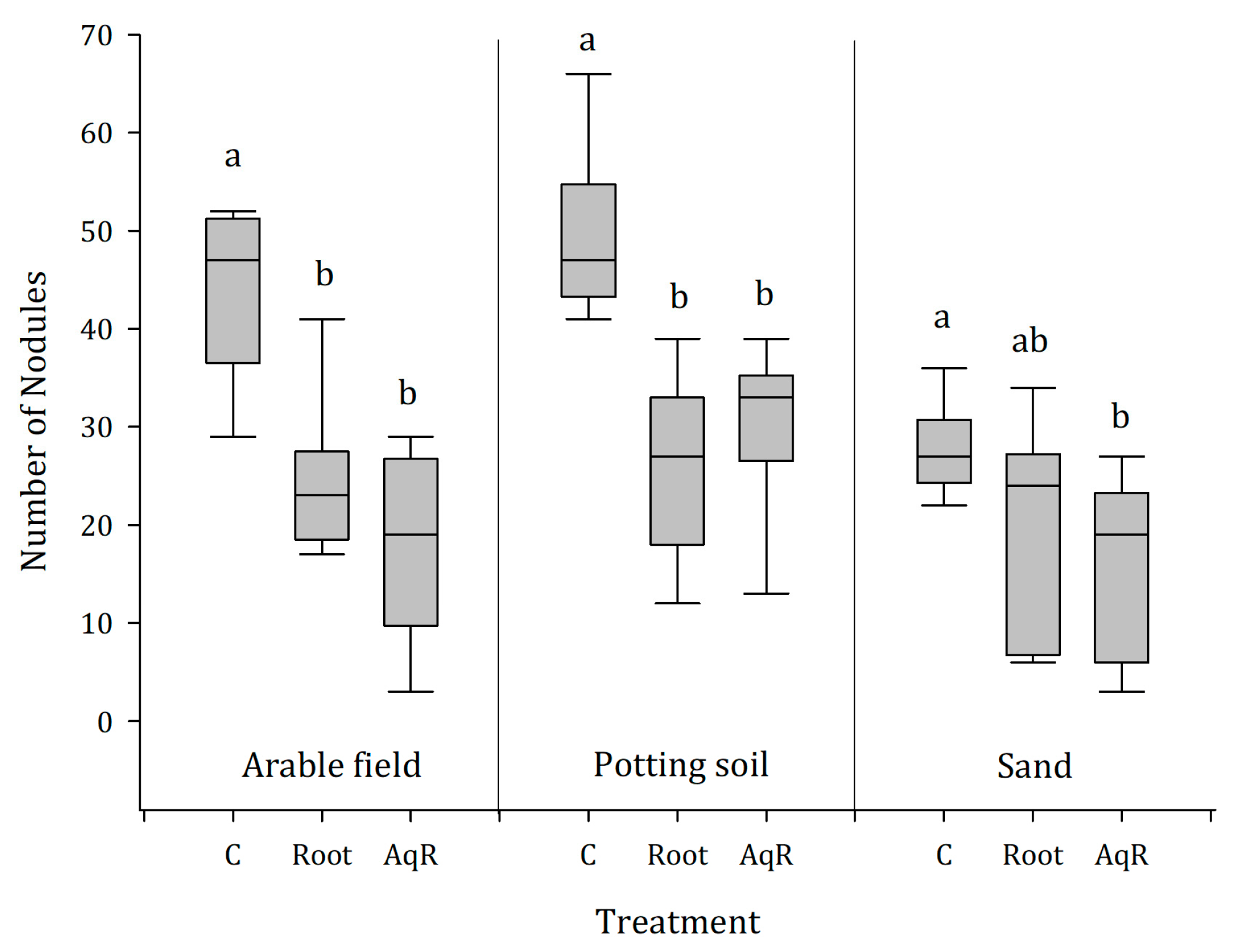
2.4. Greenhouse Experiment 2: Roots and Root Extracts Reduce Nodulation of Soybean, Irrespective of the Soil Type
3. Discussion
3.1. Effects of the Ragweed Extracts and Residues
3.2. Effects on the Nodulation with Bradyrhizobium japonicum on Soybean Roots
4. Materials and Methods
4.1. Plant Material
4.2. Extract Preparation and Chemical Analysis
4.3. Laboratory Experiment
4.4. Greenhouse Experiment 1
4.5. Greenhouse Experiment 2
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilcove, D.S.; Rothstein, D.; Dubow, J.; Phillips, A.; Losos, E. Quantifying Threats to Imperiled Species in the United States. BioScience 1998, 48, 607–615. [Google Scholar] [CrossRef]
- Pimentel, D.; Lach, L.; Zuniga, R.; Morrison, D. Environmental and economic costs of nonindigenous species in the United States. BioScience 1999, 50, 53–65. [Google Scholar] [CrossRef]
- Thorpe, A.S.; Thelen, G.C.; Diaconu, A.; Callaway, R.M. Root exudate is allelopathic in invaded community but not in native community: Field evidence for the novel weapons hypothesis. J. Ecol. 2009, 97, 641–645. [Google Scholar] [CrossRef]
- Keane, R.M.; Crawley, M.J. Exotic plant invasions and the enemy release hypothesis. Trends Ecol. Evol. 2002, 17, 164–170. [Google Scholar] [CrossRef]
- Blossey, B.; Notzold, R. Evolution of Increased Competitive Ability in Invasive Nonindigenous Plants: A Hypothesis. J. Ecol. 1995, 83, 887–889. [Google Scholar] [CrossRef]
- Callaway, R.M.; Aschehoug, E.T. Invasive plants versus their new and old neighbors: A mechanism for exotic invasion. Science 2000, 290, 521–523. [Google Scholar] [CrossRef]
- Müller-Schärer, H.; Schaffner, U.; Steinger, T. Evolution in invasive plants: Implications for biological control. Trends Ecol. Evol. 2004, 19, 417–422. [Google Scholar] [CrossRef]
- Callaway, R.M.; Ridenour, W.M. Novel weapons: Invasive success and the evolution of increased competitive ability. Front. Ecol. Environ. 2004, 2, 436–443. [Google Scholar] [CrossRef]
- Molisch, H. Der Einfluss Einer Pflanze Auf Die Andere: Allelopathie; Verlag Gustav Fischer: Jena, Germany, 1937. [Google Scholar]
- Basset, I.J.; Crompton, C.W. Biology of Canadian Weeds—Ambrosia artemisiifolia L. and Ambrosia psilostachya DC. Can. J. Plant Sci. 1975, 55, 463–476. [Google Scholar] [CrossRef]
- Karrer, G.; Milakovic, I.; Kropf, M.; Blöch, C.; Dlugosch, A.; Leitsch-Vitalos, M.; Hackl, G.; Follak, S.; Fertsak, S.; Schwab, M.; et al. Ausbreitungsbiologie und Management Einer Extrem Allergenen, Eingeschleppten Pflanze—Wege und Ursachen der Ausbreitung von Ragweed (Ambrosia artemisiifolia) Sowie Möglichkeiten Seiner Bekämpfung; Endbericht; BMLFUW: Wien, Austria, 2011. [Google Scholar]
- Smith, M.; Cecchi, L.; Skjøth, C.A.; Karrer, G.; Šikoparija, B. Common ragweed: A threat to environmental health in Europe. Environ. Int. 2013, 61, 115–126. [Google Scholar] [CrossRef]
- Varga, P.; Kazinczi, G.; Béres, I.; Kovács, I. Competition between sunflower and Ambrosia artemisiifolia in additive experiments. Cereal Res. Commun. 2006, 34, 701–704. [Google Scholar] [CrossRef]
- Kazinczi, G.; Béres, I.; Novák, R.; Bíró, K.; Pathy, Z. Common ragweed (Ambrosia artemisiifolia): A review with special regards to the results in Hungary. I. Taxonomy, origin and distribution, morphology, life cycle and reproduction strategy. Herbologia 2008, 9, 55–91. [Google Scholar]
- Hall, R.M.; Urban, B.; Wagentristl, H.; Karrer, G.; Winter, A.; Czerny, R.; Kaul, H.-P. Common ragweed (Ambrosia artemisiifolia L.) causes severe yield losses in soybean and impairs Bradyrhizobium japonicum infection. Agronomy 2021, 11, 1616. [Google Scholar] [CrossRef]
- Wopfner, N.; Gadermaier, G.; Egger, M.; Asero, R.; Ebner, C.; Jahn-Schmid, B.; Ferreira, F. The spectrum of allergens in ragweed and mugwort pollen. Int. Arch. Allergy Immunol. 2005, 138, 337–346. [Google Scholar] [CrossRef]
- Ziska, L.H.; George, K.; Frenz, D.A. Establishment and persistence of common ragweed (Ambrosia artemisiifolia L.) in disturbed soil as a function of an urban-rural macro-environment. Glob. Chang. Biol. 2007, 13, 266–274. [Google Scholar] [CrossRef]
- Prank, M.; Chapman, D.S.; Bullock, J.M.; Belmonte, J.; Berger, U.; Dahl, A.; Jäger, S.; Kovtunenko, I.; Magyar, D.; Niemelä, S.; et al. An operational model for forecasting ragweed pollen release and dispersion in Europe. Agric. For. Meteorol. 2013, 182, 43–53. [Google Scholar] [CrossRef]
- Johnson, B.; Loux, M.; Nordby, D.; Sprague, C.; Nice, G.; Westhoven, A.; Stachler, J. Biology and Management of Giant Ragweed; GWC; University of Missouri: Columbia, MO, USA, 2007. [Google Scholar]
- Fumanal, B.; Chauvel, B.; Sabatier, A.; Bretagnolle, F. Variability and cryptic heteromorphism of Ambrosia artemisiifolia seeds: What consequences for its invasion in France? Ann. Bot. 2007, 100, 305–313. [Google Scholar] [CrossRef]
- Essl, F.; Biró, K.; Brandes, D.; Broennimann, O.; Bullock, J.M.; Chapman, D.S.; Chauvel, B.; Dullinger, S.; Fumanal, B.; Guisan, A.; et al. Biological Flora of the British Isles: Ambrosia artemisiifolia. J. Ecol. 2015, 103, 1069–1098. [Google Scholar] [CrossRef]
- Han, C.; Shao, H.; Zhou, S.; Mei, Y.; Cheng, Z.; Huang, L.; Lv, G. Chemical composition and phytotoxicity of essential oil from invasive plant, Ambrosia artemisiifolia L. Ecotoxicol. Environ. Saf. 2021, 211, 111879. [Google Scholar] [CrossRef]
- Šućur, J.; Konstantinović, B.; Crnković, M.; Bursić, V.; Samardžić, N.; Malenčić, Đ.; Prvulović, D.; Popov, M.; Vuković, G. Chemical composition of Ambrosia trifida L. and its allelopathic influence on crops. Plants 2021, 10, 2222. [Google Scholar] [CrossRef]
- Janaćković, P.; Rajčević, N.; Gavrilović, M.; Novaković, J.; Radulović, M.; Miletić, M.; Janakiev, T.; Dimkić, I.; Marin, P.D. Essential oil composition of Ambrosia artemisiifolia and its antibacterial activity against phytopathogens. Biol. Life Sci. Forum. 2022, 15, 22. [Google Scholar] [CrossRef]
- Pajević, S.; Borišev, M.; Orčić, D.; Boža, P.; Nikolić, N. Photosynthetic and biochemical characteristics of invasive species (Ambrosia artemisiifolia L., Ambrosia trifida L. and Iva xanthifolia Nutt.) depending on soil humidity and phenological phase. Russ. J. Ecol. 2010, 41, 498–505. [Google Scholar] [CrossRef]
- Vidotto, F.; Tesio, F.; Ferrero, A. Allelopathic effects of Ambrosia artemisiifolia L. in the invasive process. J. Crop Prot. 2013, 54, 161–167. [Google Scholar] [CrossRef]
- Bonea, D.; Bonciu, E.; Niculescu, M.; Olaru, A.L. The allelopathic, cytotoxic and genotoxic effect of Ambrosia artemisiifolia on the germination and root meristems of Zea mays. Caryologia 2018, 71, 24–28. [Google Scholar] [CrossRef]
- Molinaro, F.; Monterumici, C.M.; Ferrero, A.; Tabasso, S.; Negre, M. Bioherbicidal activity of a germacranolide sesquiterpene dilactone from Ambrosia artemisiifolia L.—Part B Pesticides, Food Contaminants, and Agricultural Wastes. J. Environ. Sci. Health 2016, 51, 847–852. [Google Scholar] [CrossRef]
- Zhao, F.; Elkelish, A.; Durner, J.; Lindermayr, C.; Winkler, J.B.; Ruioff, F.; Behrendt, H.; Traidl-Hoffmann, C.; Holzinger, A.; Kofler, W.; et al. Common ragweed (Ambrosia artemisiifolia L.): Allergenicity and molecular characterization of pollen after plant exposure to elevated NO2. Plant Cell Environ. 2016, 39, 147–164. [Google Scholar] [CrossRef]
- Zhang, W.; Ren, C.; Deng, J.; Zhao, F.; Yang, G.; Han, X.; Tong, X.; Feng, Y. Plant functional composition and species diversity affect soil C, N, and P during secondary succession of abandoned farmland on the Loess Plateau. Ecol. Eng. 2018, 122, 91–99. [Google Scholar] [CrossRef]
- Liu, S.-H.; Kazemi, S.; Karrer, G.; Bellaire, A.; Weckwert, W.; Damkjaer, J.; Hoffmann, O.; Epstein, M.M. Influence of the environment on ragweed pollen and their sensitizing capacity in a mouse model of allergic lung inflammation. Front. Allergy 2022, 3, 854038. [Google Scholar] [CrossRef]
- Dudley, S.A.; Murphy, G.P.; File, A.L. Kin recognition and competition in plants. Funct. Ecol. 2013, 27, 898–906. [Google Scholar] [CrossRef]
- Ninkovic, V.; Markovic, D.; Dahlin, I. Decoding neighbour volatiles in preparation for future competition and implications for tritrophic interactions. Perspect. Plant Ecol. Evol. Syst. 2016, 23, 11–17. [Google Scholar] [CrossRef]
- Su, P.; Liu, X.; Wang, R.; Liu, T.; Zhao, W.; Sun, M.; Wang, H.; Liu, Y.; Wu, Q. Autotoxicity of Ambrosia artemisiifolia and Ambrosia trifida and its significance for the regulation of intraspecific populations density. Sci. Rep. 2022, 12, 17424. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Wu, B.; Jiang, K.; Zhou, J.; Liu, J.; Lv, Y. Canada goldenrod invasion cause significant shifts in the taxonomic diversity and community stability of plant communities in heterogeneous landscapes in urban ecosystems in East China. Ecol. Eng. 2018, 301, 1–6. [Google Scholar] [CrossRef]
- Casiglia, S.; Bruno, M.; Bramucci, M.; Quassinti, L.; Lupidi, G.; Fiorini, D.; Maggi, F. Kundmannia sicula (L.) DC: A rich source of germacrene D. J. Essent. Oil Res. 2017, 29, 437–442. [Google Scholar] [CrossRef]
- Nijenstein, J.H.; Ester, A. Phytotoxicity and control of the field slug Deroceras reticulatum by seed applied pesticides in wheat, barley and perennial rygrass. Seed Sci. Technol. 1998, 26, 501–513. [Google Scholar]
- Ibrahim, M.A.; Oksanen, E.J.; Holopainen, J.K. Effects of limonene on the growth and physiology of cabbage (Brassica oleracea L.) and carrot (Daucus carota L.) plants. J. Sci. Food Agric. 2004, 84, 1319–1326. [Google Scholar] [CrossRef]
- Shao, H.; Zhang, Y.M.; Nan, P.; Huang, X.L.; Zhang, C. Chemical composition and phytotoxic activity of the volatile oil of invasive Xanthium italicum Moretti from Xinjiang, China. J. Arid. Land 2013, 5, 324–330. [Google Scholar] [CrossRef]
- Zunino, M.P.; Zygadlo, J.A. Effect of monoterpenes on lipid oxidation in maize. Planta 2004, 219, 303–309. [Google Scholar] [CrossRef]
- Singh, H.P.; Kaur, S.; Mittal, S.; Batish, D.R.; Kohli, R.K. Essential oil of Artemisia scoparia inhibits plant growth by generating reactive oxygen species and causing oxidative damage. J. Chem. Ecol. 2009, 35, 154–162. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Kaur, S.; Arora, K.; Kohli, R.K. α-Pinene inhibits growth and induces oxidative stress in roots. Ann. Bot. 2006, 98, 1261–1269. [Google Scholar] [CrossRef]
- Nishida, N.; Tamotsu, S.; Nagata, N.; Saito, C.; Sakai, A. Allelopathic effects of volatile monoterpenoids produced by Salvia leucophylla: Inhibition of cell proliferation and DNA synthesis in the root apical meristem of Brassica campestris seedlings. J. Chem. Ecol. 2005, 31, 1187–1203. [Google Scholar] [CrossRef]
- Chowhan, N.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Phytotoxic effects of β-pinene on early growth and associated biochemical changes in rice. Acta Physiol. Plant 2011, 33, 2369–2376. [Google Scholar] [CrossRef]
- Nagashima, A.; Higaki, T.; Koeduka, T.; Ishigami, K.; Hosokawa, S.; Watanabe, H.; Matsui, K.; Hasezawa, S.; Touhara, K. Transcriptional regulators involved in responses to volatile organic compounds in plants. J. Biol. Chem. 2019, 294, 2256–2266. [Google Scholar] [CrossRef]
- Frank, L.; Wenig, M.; Ghirardo, A.; van der Krol, A.; Vlot, A.C.; Schnitzler, J.-P.; Rosenkranz, M. Isoprene and β-caryophyllene confer plant resistance via different plant internal signalling pathways. Plant Cell Environ. 2021, 44, 1151–1164. [Google Scholar] [CrossRef]
- Ditengou, F.A.; Müller, A.; Rosenkranz, M.; Felten, J.; Lasok, H.; Miloradovic van Doorn, M.; Legué, V.; Palme, K.; Schnitzler, J.-P.; Polle, A. Volatile signalling by sesquiterpenes from ectomycorrhizal fungi reprogramms root architecture. Nat. Commun. 2015, 6, 6279. [Google Scholar] [CrossRef]
- Araniti, F.; Sánchez-Moreiras, A.M.; Graña, E.; Reigosa, M.J.; Abenavoli, M.R. Terpenoid trans-caryophyllene inhibits weed germination and induces plant water status alteration and oxidative damage in adult Arabidopsis. Plant Biol. 2016, 19, 79–89. [Google Scholar] [CrossRef]
- Verma, M.; Rao, P.B. Allelopathic effect of four weed species extracts on germination, growth and protein in different varieties of Glycine max (L.) Merrill. J. Environ. Biol. 2006, 27, 571–577. [Google Scholar]
- Kincel, K.; Ramona, Ș.; Alin, C. Influence of Ambrosia artemisiifolia extract on germination and growth of Amaranthus retroflexus and Zea mays. Res. J. Agric. Sci. 2019, 51, 127–135. [Google Scholar]
- Maksimović, T.; Marković, L.; Hasanagić, D. Influence of Ambrosia artemisiifolia extract on germination and growth of Pisum sativum L. and Phaseolus vulgaris L. seedlings. Zemlj biljka. Res. J. Agric. Sci. 2021, 70, 33–41. [Google Scholar] [CrossRef]
- Hall, R.M.; Markovic, D.; Kaul, H.-P.; Urban, B.; Durec, N.; Renner-Martin, K.; Ninkovic, V. Airborne volatile organic compounds of the invasive common ragweed (Ambrosia artemisiifolia L.) alter growth of wheat, soybean, and maize. In What Makes Common Ragweed a Fierce Competitor with Crops? University of Natural Resources and Life Science: Vienna, Austria, 2022. [Google Scholar]
- Savić, A.; Oveisi, M.; Božić, D.; Pavlović, D.; Saulić, M.; Schärer, H.M.; Vrbničanin, S. Competition between Ambrosia artemisiifolia and Ambrosia trifida: Is there a threat of a stronger competitor? Weed Res. 2021, 61, 298–306. [Google Scholar] [CrossRef]
- Vaou, N.; Stavropoulou, E.; Voidarou, C.; Tsigalou, C.; Bezirtzoglou, E. Towards advances in medicinal plant antimicrobial activity: A review study on challenges and future perspectives. Microorganisms 2021, 9, 2041. [Google Scholar] [CrossRef]
- Puppo, A.; Pauly, N.; Boscari, A.; Mandon, K.; Brouquisse, R. Hydrogen peroxide and nitric oxide: Key regulators of the legume—Rhizobium and mycorrhizal symbioses. Antioxid. Redox Signal. 2013, 18, 2202–2219. [Google Scholar] [CrossRef] [PubMed]
- Gal, A.; Clarence, J.; Gal, J.; Afifi, M.; Lee, E.; Lukens, L.; Swanton, C.J. Detection of neighboring weeds alters soybean seedling roots and nodulation. Weed Sci. 2015, 63, 888–900. [Google Scholar] [CrossRef]
- Atkinson, J.H.; Harris, R.F. Atmospheric degradation of volatile organic compounds. Chem. Rev. 2003, 103, 4605–4638. [Google Scholar] [CrossRef] [PubMed]
- McInnis, S.M.; Desikan, R.; Hancock, J.T.; Hiscock, S.J. Production of reactive oxygen species and reactive nitrogen species by angiosperm stigmas and pollen: Potential signalling crosstalk? New Phytol. 2006, 172, 221–228. [Google Scholar] [CrossRef]
- Peláez-Vico, M.A.; Bernabéu-Roda, L.; Kohlen, W.; Soto, M.J.; López-Ráez, J.A. Strigolactones in the Rhizobium-legume symbiosis: Stimulatory effect on bacterial surface motility and down-regulation of their levels in nodulated plants. Plant Sci. 2016, 245, 119–127. [Google Scholar] [CrossRef]
- Hong, N.H.; Xuan, T.D.; Tsuzuki, E.; Terao, H.; Matsuo, M.; Khanh, T.D. Weed Control of Four Higher Plant Species in Paddy Rice Fields in Southeast Asia. J. Agron. Crop. Sci. 2004, 190, 59–64. [Google Scholar] [CrossRef]
- Khanh, T.D.; Hong, N.H.; Xuan, T.D.; Chung, I.M. Paddy weed control by medicinal and leguminous plants from Southeast Asia. Crop. Prot. 2005, 24, 421–431. [Google Scholar] [CrossRef]
- Kazinczi, G.; Béres, I.; Onofir, A.; Nádasy, E.; Takács, A.; Horvát, J.; Torma, M. Allelopathic effects of plant extracts on common ragweed. J. Plant Dis. Prot. 2008, 21, 335–340. [Google Scholar]
- Ritz, C.; Baty, F.; Steibig, J.; Gerhard, D. Dose-response analysis using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hall, R.M.; Wagentristl, H.; Renner-Martin, K.; Urban, B.; Durec, N.; Kaul, H.-P. Extracts and Residues of Common Ragweed (Ambrosia artemisiifolia L.) Cause Alterations in Root and Shoot Growth of Crops. Plants 2023, 12, 1768. https://doi.org/10.3390/plants12091768
Hall RM, Wagentristl H, Renner-Martin K, Urban B, Durec N, Kaul H-P. Extracts and Residues of Common Ragweed (Ambrosia artemisiifolia L.) Cause Alterations in Root and Shoot Growth of Crops. Plants. 2023; 12(9):1768. https://doi.org/10.3390/plants12091768
Chicago/Turabian StyleHall, Rea Maria, Helmut Wagentristl, Katharina Renner-Martin, Bernhard Urban, Nora Durec, and Hans-Peter Kaul. 2023. "Extracts and Residues of Common Ragweed (Ambrosia artemisiifolia L.) Cause Alterations in Root and Shoot Growth of Crops" Plants 12, no. 9: 1768. https://doi.org/10.3390/plants12091768