
Features of Photosynthesis in Arabidopsis thaliana Plants with Knocked Out Gene of Alpha Carbonic Anhydrase 2
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Measurement of Chlorophyll a Fluorescence and P700 Absorption Changes at Room Temperature
2.3. Measurements of Electron Transport Rate in Thylakoids
2.4. Evaluation of State Transitions
2.5. Electrochromic Carotenoid Absorbance Shift Measurements
2.6. Western Blot Analysis
2.7. Determination of Starch Content
2.8. Quantitative Reverse Transcription PCR
2.9. Statistical Analysis
3. Results
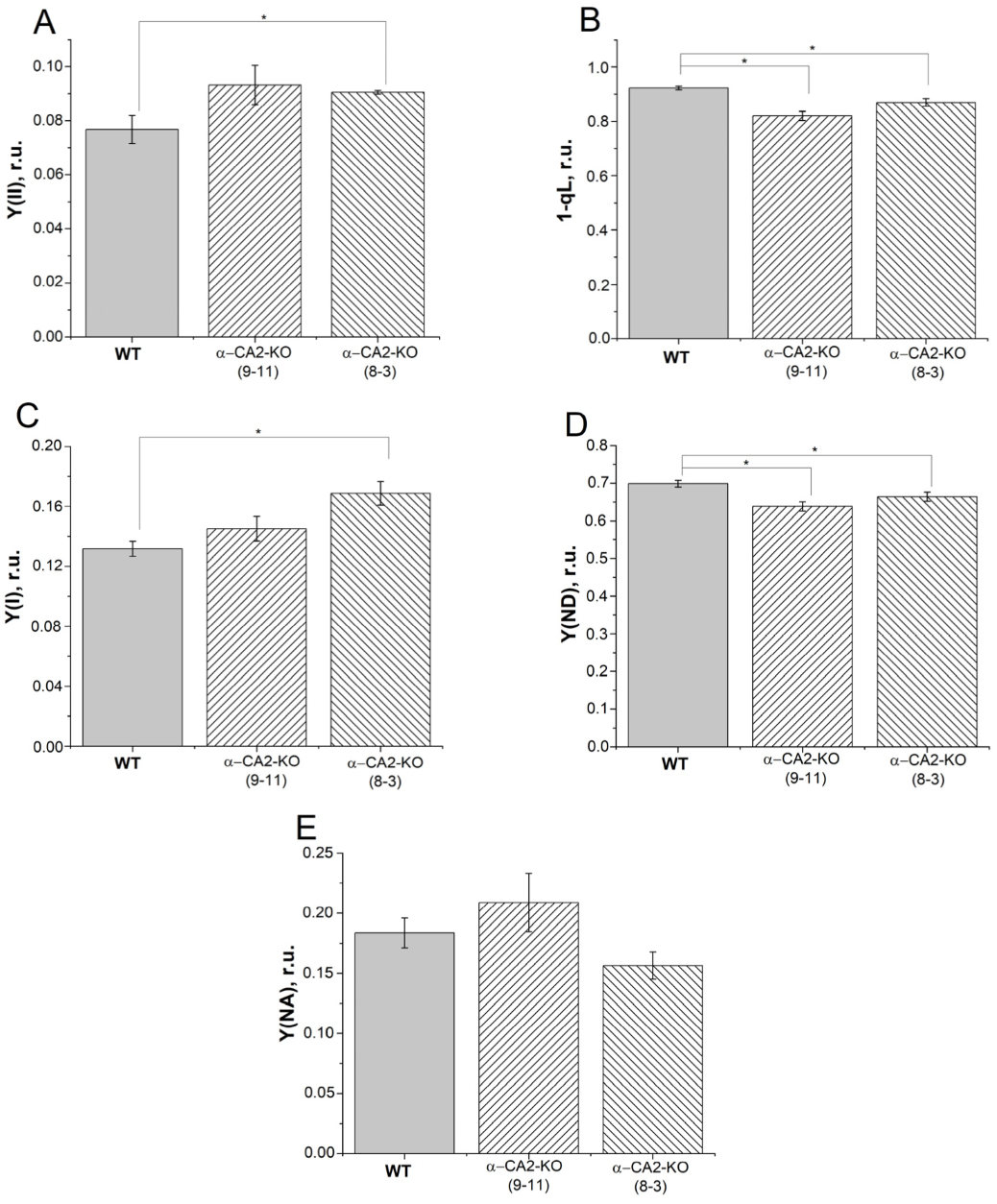
3.1. Effect of α-CA2 Absence on the Photosynthetic Electron Transport in Intact Leaves
3.2. Effect of α-CA2 Absence on the Electron Transport in Isolated Thylakoids
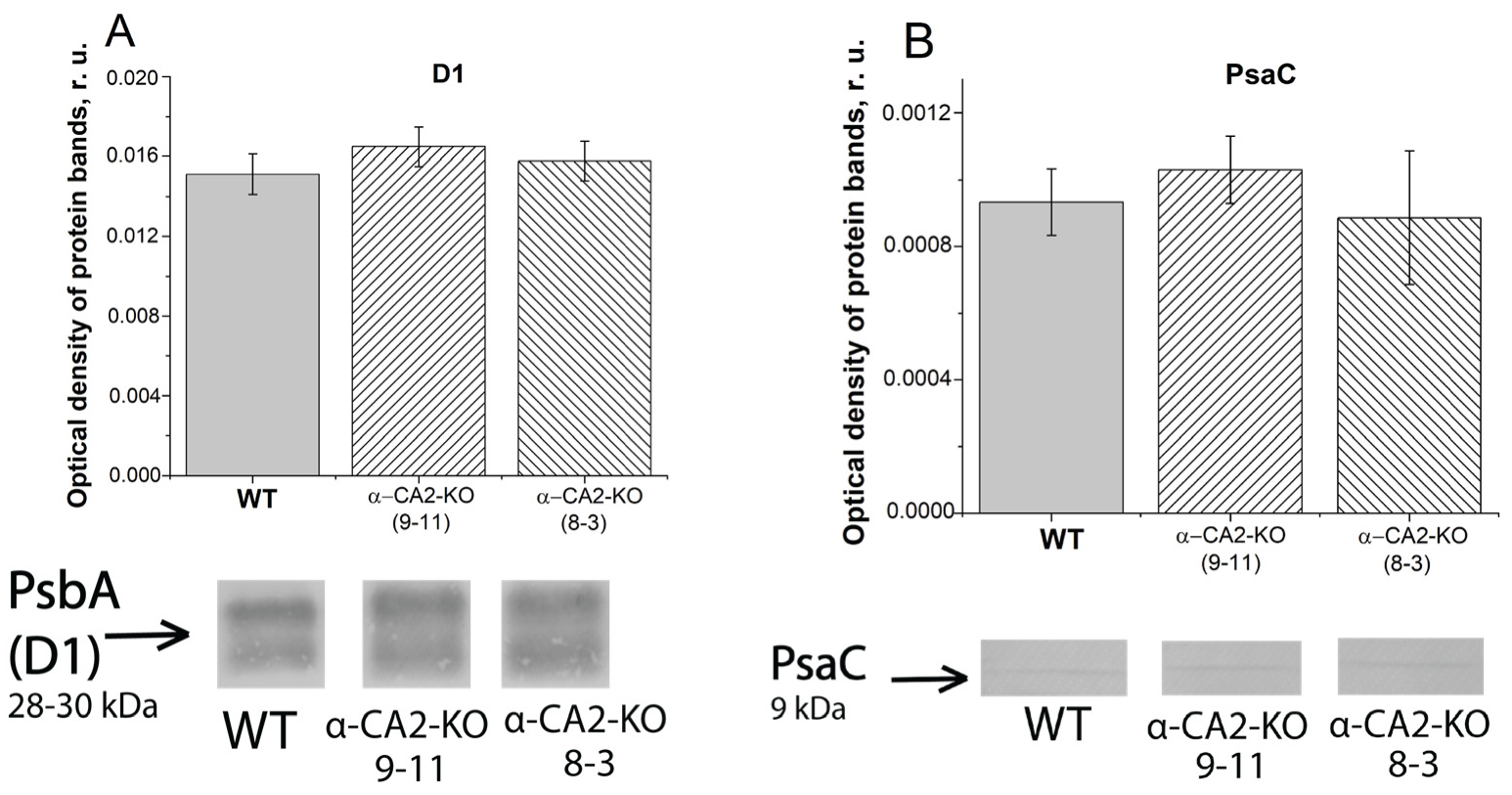
3.3. Effect of α-CA2 Absence on the Content of Photosynthetic Reaction Centers Proteins
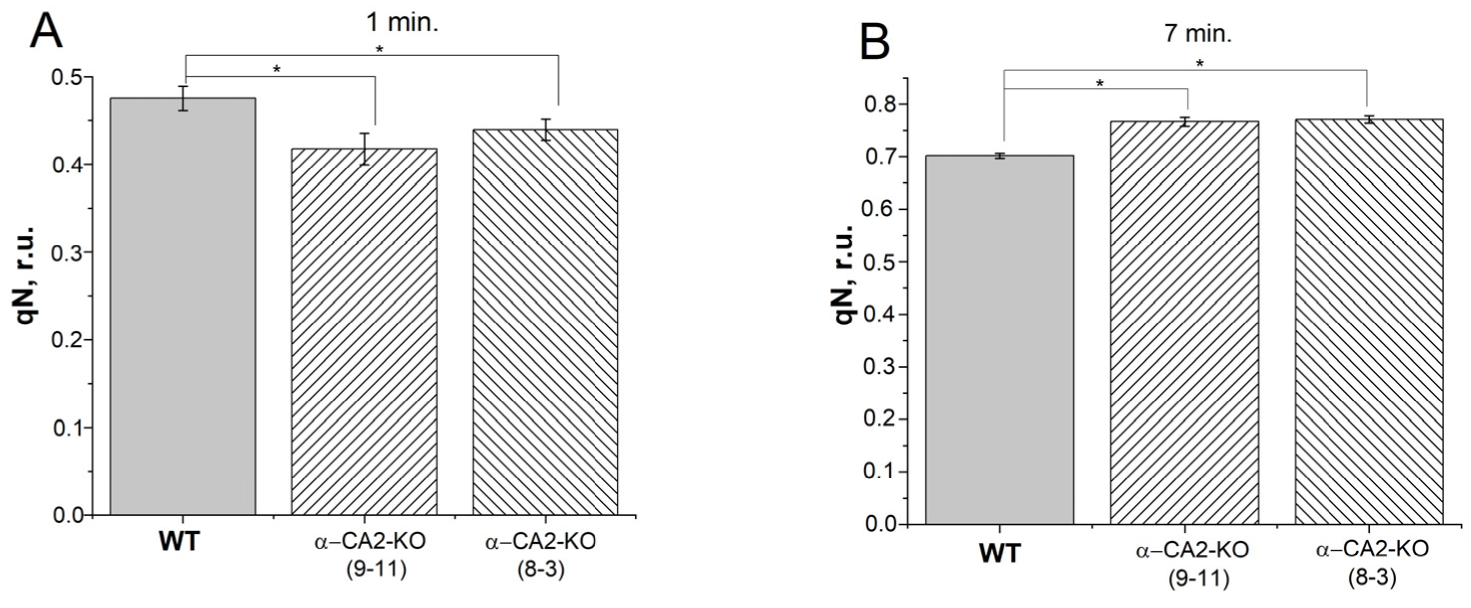
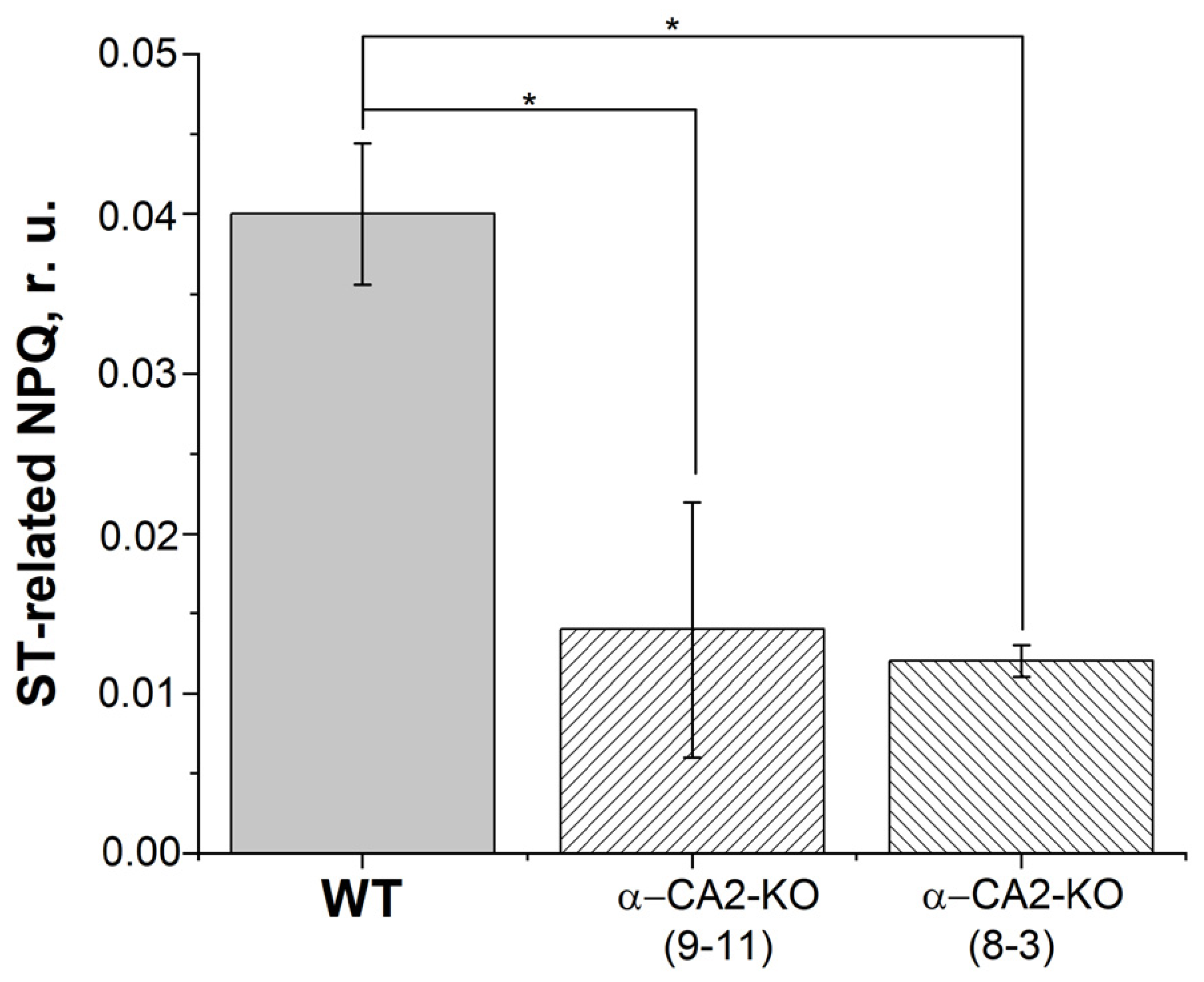
3.4. Effect of α-CA2 Absence on Non-Photochemical Quenching of Chlorophyll Fluorescence
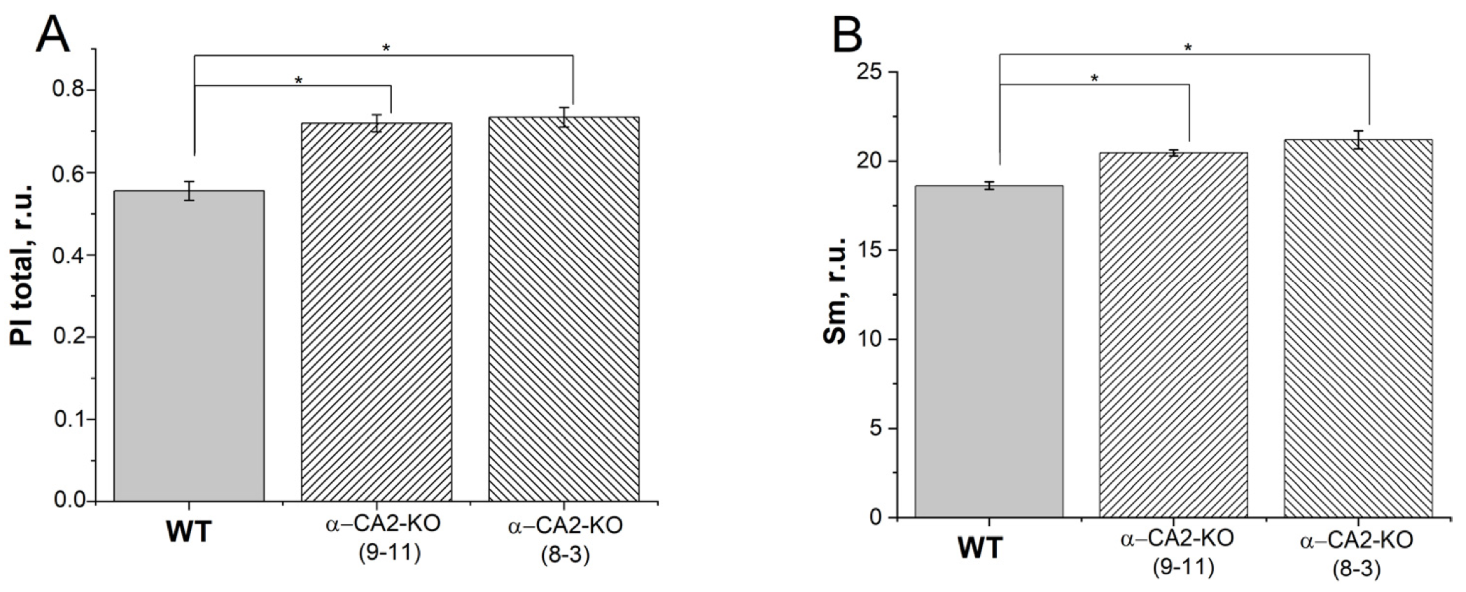
3.5. Effect of α-CA2 Absence on the Susceptibility to Photoinhibition
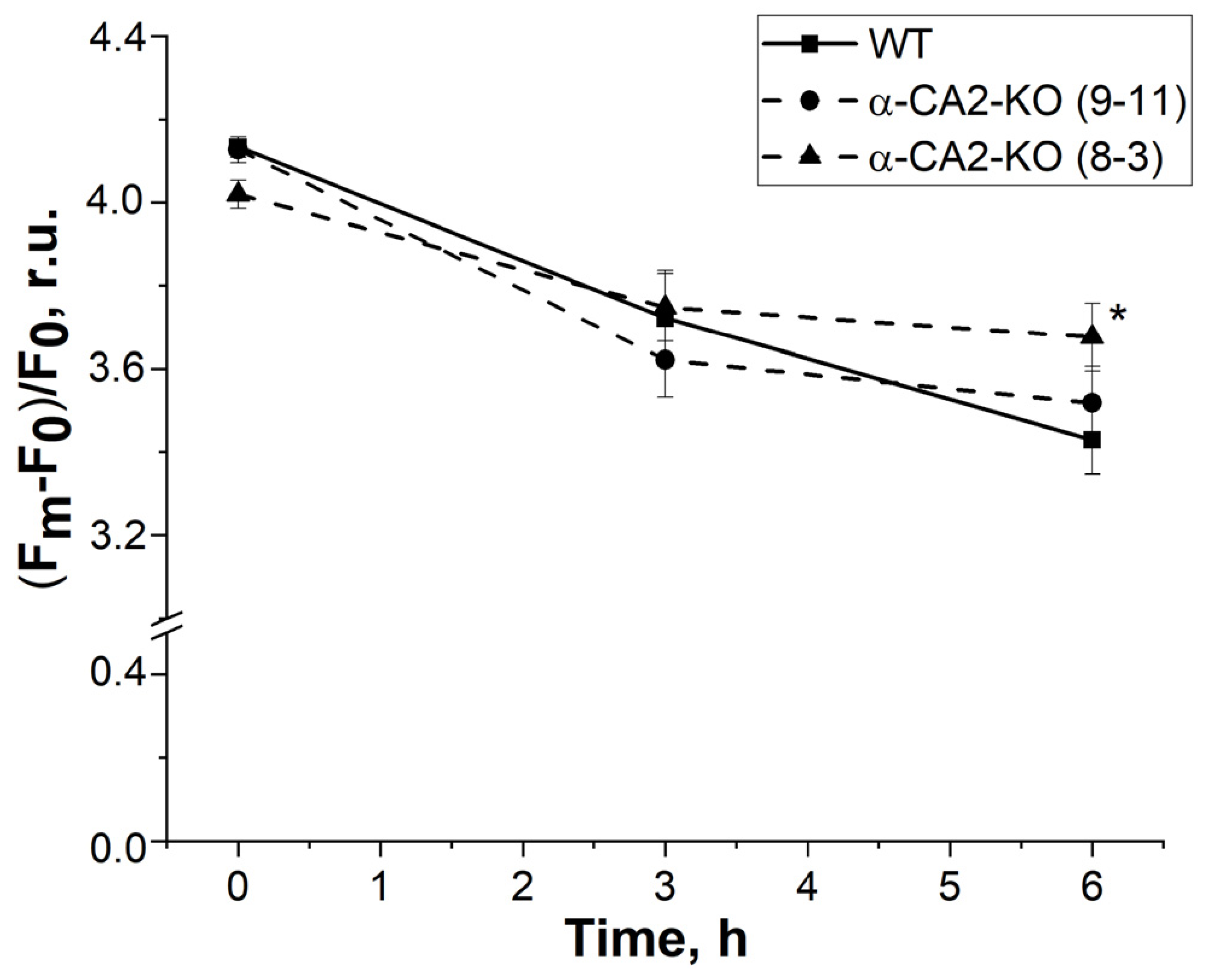
3.6. Effect of α-CA2 Absence on State Transitions
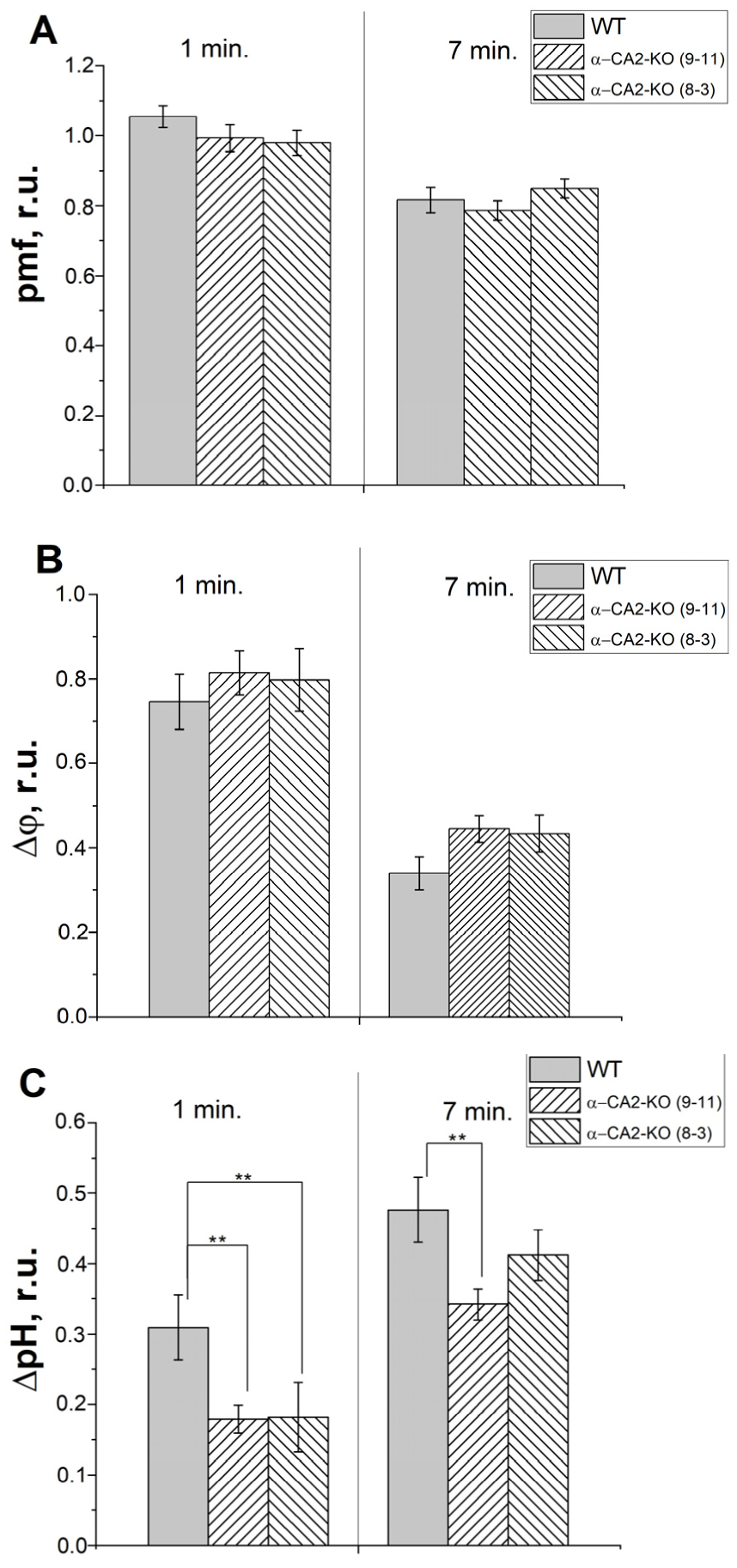
3.7. Effect of α-CA2 Absence on the Characteristics of Electrochromic Shift in Intact Leaves
3.8. Effect of α-CA2 Absence on the Amount of Starch in Leaves
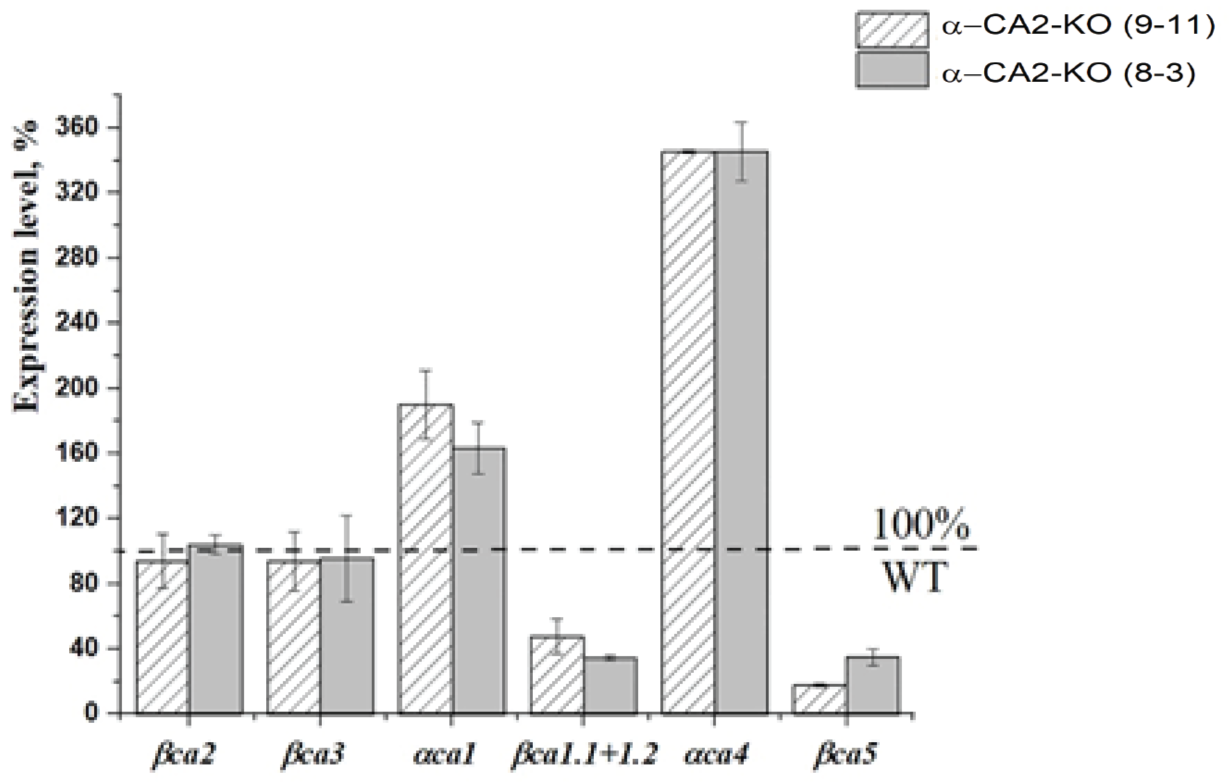
3.9. Effect of α-CA2 Absence on the Expression Levels of the Cytoplasmic and Chloroplast Carbonic Anhydrases
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| α-CA4-KO | α-CA4 knockout |
| α-CA2-KO | α-CA2 knockout |
| CA | carbonic anhydrase |
| ECS | electrochromic carotenoid absorbance shift |
| KEA3 | K+/H+ exchange antiporter |
| LHCII | light-harvesting complex of photosystem II |
| MV | methyl viologen |
| NPQ | non-photochemical quenching of chlorophyll a fluorescence |
| PETC | photosynthetic electron transport chain |
| pmf | proton motive force |
| PQ | plastoquinone |
| PQH2 | plastohydroquinone |
| PSII | photosystem II |
| PSI | photosystem I |
| Rubisco | ribulose bisphosphate carboxylase/oxygenase |
| WT | wild type |
References
- Johansson, I.-M.; Forsman, C. Kinetic Studies of Pea Carbonic Anhydrase. Eur. J. Biochem. 1993, 218, 439–446. [Google Scholar] [CrossRef]
- Villarejo, A.; Burén, S.; Larsson, S.; Déjardin, A.; Monné, M.; Rudhe, C.; Karlsson, J.; Jansson, S.; Lerouge, P.; Rolland, N.; et al. Evidence for a Protein Transported through the Secretory Pathway En Route to the Higher Plant Chloroplast. Nat. Cell Biol. 2005, 7, 1224–1231. [Google Scholar] [CrossRef] [PubMed]
- Fabre, N.; Reiter, I.M.; Becuwe-Linka, N.; Genty, B.; Rumeau, D. Characterization and Expression Analysis of Genes Encoding alfaand beta Carbonic Anhydrases in Arabidopsis. Plant Cell Environ. 2007, 30, 617–629. [Google Scholar] [CrossRef]
- Shen, J.; Li, Z.; Fu, Y.; Liang, J. Identification and Molecular Characterization of the Alternative Spliced Variants of Beta Carbonic Anhydrase 1 (ΒCA1) from Arabidopsis thaliana. PeerJ 2021, 9, e12673. [Google Scholar] [CrossRef]
- Friso, G.; Giacomelli, L.; Ytterberg, A.J.; Peltier, J.-B.; Rudella, A.; Sun, Q.; van Wijk, K.J. In-Depth Analysis of the Thylakoid Membrane Proteome of Arabidopsis Thaliana Chloroplasts: New Proteins, New Functions, and a Plastid Proteome Database. Plant Cell 2004, 16, 478–499. [Google Scholar] [CrossRef] [PubMed]
- Ignatova, L.; Zhurikova, E.; Ivanov, B. The Presence of the Low Molecular Mass Carbonic Anhydrase in Photosystem II of C3 Higher Plants. J. Plant Physiol. 2019, 232, 94–99. [Google Scholar] [CrossRef]
- Fedorchuk, T.P.; Kireeva, I.A.; Opanasenko, V.K.; Terentyev, V.V.; Rudenko, N.N.; Borisova-Mubarakshina, M.M.; Ivanov, B.N. Alpha Carbonic Anhydrase 5 Mediates Stimulation of ATP Synthesis by Bicarbonate in Isolated Arabidopsis Thylakoids. Front. Plant Sci. 2021, 12, 82. [Google Scholar] [CrossRef]
- Rudenko, N.N.; Ignatova, L.K.; Ivanov, B.N. Multiple Sources of Carbonic Anhydrase Activity in Pea Thylakoids: Soluble and Membrane-Bound Forms. Photosynth. Res. 2007, 91, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Fedorchuk, T.; Rudenko, N.; Ignatova, L.; Ivanov, B. The Presence of Soluble Carbonic Anhydrase in the Thylakoid Lumen of Chloroplasts from Arabidopsis Leaves. J. Plant Physiol. 2014, 171, 903–906. [Google Scholar] [CrossRef]
- Rudenko, N.N.; Ivanov, B.N. Unsolved Problems of Carbonic Anhydrases Functioning in Photosynthetic Cells of Higher C3 Plants. Biochem. Mosc. 2021, 86, 1243–1255. [Google Scholar] [CrossRef]
- Fedorchuk, T.P.; Opanasenko, V.K.; Rudenko, N.N.; Ivanov, B.N. Bicarbonate-induced stimulation of photophosphorylation in isolated thylakoids: Effects of carbonic anhydrase inhibitors. Biol. Membr. 2018, 35, 34–41. [Google Scholar] [CrossRef]
- Punnett, T.; Iyer, R.V. The Enhancement of Photophosphorylation and the Hill Reaction by Carbon Dioxide. J. Biol. Chem. 1964, 239, 2335–2339. [Google Scholar] [CrossRef] [PubMed]
- Khristin, M.S.; Ignatova, L.K.; Rudenko, N.N.; Ivanov, B.N.; Klimov, V.V. Photosystem II Associated Carbonic Anhydrase Activity in Higher Plants Is Situated in Core Complex. FEBS Lett. 2004, 577, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Ignatova, L.K.; Rudenko, N.N.; Khristin, M.S.; Ivanov, B.N. Heterogeneous Origin of Carbonic Anhydrase Activity of Thylakoid Membranes. Biochem. Mosc. 2006, 71, 525–532. [Google Scholar] [CrossRef]
- Ignatova, L.K.; Rudenko, N.N.; Mudrik, V.A.; Fedorchuk, T.P.; Ivanov, B.N. Carbonic Anhydrase Activity in Arabidopsis Thaliana Thylakoid Membrane and Fragments Enriched with PSI or PSII. Photosynth. Res. 2011, 110, 89–98. [Google Scholar] [CrossRef]
- Zhurikova, Е.М.; Ignatova, L.K.; Semenova, G.A.; Rudenko, N.N.; Mudrik, V.A.; Ivanov, B.N. Effect of Knockout of α-Carbonic Anhydrase 4 Gene on Photosynthetic Characteristics and Starch Accumulation in Leaves of Arabidopsis Thaliana. Russ. J. Plant 2015, 62, 602–608. [Google Scholar] [CrossRef]
- Rudenko, N.N.; Fedorchuk, T.P.; Vetoshkina, D.V.; Zhurikova, E.M.; Ignatova, L.K.; Ivanov, B.N. Influence of Knockout of At4g20990 Gene Encoding α-CA4 on Photosystem II Light-Harvesting Antenna in Plants Grown under Different Light Intensities and Day Lengths. Protoplasma 2018, 255, 69–78. [Google Scholar] [CrossRef]
- Rudenko, N.N.; Permyakova, N.V.; Ignatova, L.K.; Nadeeva, E.M.; Zagorskaya, A.A.; Deineko, E.V.; Ivanov, B.N. The Role of Carbonic Anhydrase ACA4 in Photosynthetic Reactions in Arabidopsis Thaliana Studied, Using the Cas9 and T-DNA Induced Mutations in Its Gene. Plants 2022, 11, 3303. [Google Scholar] [CrossRef]
- Rudenko, N.N.; Fedorchuk, T.P.; Terentyev, V.V.; Dymova, O.V.; Naydov, I.A.; Golovko, T.K.; Borisova-Mubarakshina, M.M.; Ivanov, B.N. The Role of Carbonic Anhydrase α-CA4 in the Adaptive Reactions of Photosynthetic Apparatus: The Study with α-CA4 Knockout Plants. Protoplasma 2020, 257, 489–499. [Google Scholar] [CrossRef]
- Rudenko, N.N.; Vetoshkina, D.V.; Fedorchuk, T.P.; Ivanov, B.N. Effect of Light Intensity under Different Photoperiods on Expression Level of Carbonic Anhydrase Genes of the α- and β-Families in Arabidopsis Thaliana Leaves. Biochem. Mosc. 2017, 82, 1025–1035. [Google Scholar] [CrossRef]
- Zhurikova, E.M.; Ignatova, L.K.; Rudenko, N.N.; Mudrik, V.A.; Vetoshkina, D.V.; Ivanov, B.N. Participation of Two Carbonic Anhydrases of the Alpha Family in Photosynthetic Reactions in Arabidopsis Thaliana. Biochem. Mosc. 2016, 81, 1182–1187. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, A.M.; Yamamoto, H.Y. Zeaxanthin Formation and Energy-Dependent Fluorescence Quenching in Pea Chloroplasts under Artificially Mediated Linear and Cyclic Electron Transport. Plant Physiol. 1991, 96, 635–643. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C.; Knapp, M. How to Correctly Determine the Different Chlorophyll Fluorescence Parameters and the Chlorophyll Fluorescence Decrease Ratio RFd of Leaves with the PAM Fluorometer. Photosynthetica 2005, 43, 379–393. [Google Scholar] [CrossRef]
- Klughammer, C.; Schreiber, U. Complementary PS II Quantum Yields Calculated from Simple Fluorescence Parameters Measured by PAM Fluorometry and the Saturation Pulse Method. PAM Appl. Notes 2008, 1, 201–247. [Google Scholar]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The Fluorescence Transient as a Tool to Characterize and Screen Photosynthetic Samples. In Probing Photosynthesis: Mechanisms, Regulation and Adaptation; Yunus, M., Pathre, U., Mohanty, P., Eds.; Taylor and Francis: London, UK, 2000; pp. 445–483. [Google Scholar]
- Casazza, A.P.; Tarantino, D.; Soave, C. Preparation and Functional Characterization of Thylakoids from Arabidopsis Thaliana. Photosynth. Res. 2001, 68, 175–180. [Google Scholar] [CrossRef]
- McCormac, D.J.; Bruce, D.; Greenberg, B.M. State Transitions, Light-Harvesting Antenna Phosphorylation and Light-Harvesting Antenna Migration In Vivo in the Higher Plant Spirodela Oligorrhiza. Biochim. Biophys. Acta BBA Bioenerg. 1994, 1187, 301–312. [Google Scholar] [CrossRef]
- Vetoshkina, D.V.; Kozuleva, M.A.; Terentyev, V.V.; Zhurikova, E.M.; Borisova- Mubarakshina, M.M.; Ivanov, B.N. Comparison of State Transitions of the Photosynthetic Antennae in Arabidopsis and Barley Plants upon Illumination with Light of Various Intensity. Biochem. Mosc. 2019, 84, 1065–1073. [Google Scholar] [CrossRef]
- Kaplan, F.; Zhao, W.; Richards, J.T.; Wheeler, R.M.; Guy, C.L.; Levine, L.H. Transcriptional and Metabolic Insights into the Differential Physiological Responses of Arabidopsis to Optimal and Supraoptimal Atmospheric CO2. PLoS ONE 2012, 7, e43583. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The Relationship between the Quantum Yield of Photosynthetic Electron Transport and Quenching of Chlorophyll Fluorescence. Biochim. Biophys. Acta BBA Gen. Subj. 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Klughammer, C.; Schreiber, U. An Improved Method, Using Saturating Light Pulses, for the Determination of Photosystem I Quantum Yield via P700+-Absorbance Changes at 830 Nm. Planta 1994, 192, 261–268. [Google Scholar] [CrossRef]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New Fluorescence Parameters for the Determination of QA Redox State and Excitation Energy Fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef]
- Tóth, S.Z.; Schansker, G.; Strasser, R.J. A Non-Invasive Assay of the Plastoquinone Pool Redox State Based on the OJIP-Transient. Photosynth. Res. 2007, 93, 193. [Google Scholar] [CrossRef] [PubMed]
- Puthiyaveetil, S. A Mechanism for Regulation of Chloroplast LHC II Kinase by Plastoquinol and Thioredoxin. FEBS Lett. 2011, 585, 1717–1721. [Google Scholar] [CrossRef]
- Leong, T.-Y.; Anderson, J.M. Adaptation of the Thylakoid Membranes of Pea Chloroplasts to Light Intensities. II. Regulation of Electron Transport Capacities, Electron Carriers, Coupling Factor (CF1) Activity and Rates of Photosynthesis. Photosynth. Res. 1984, 5, 117–128. [Google Scholar] [CrossRef]
- Hangarter, R.; Ort, D.R. The Relationship between Light-Induced Increases in the H+ Conductivity of Thylakoid Membranes and Activity of the Coupling Factor. Eur. J. Biochem. 1986, 158, 7–12. [Google Scholar] [CrossRef]
- Ruban, A.V.; Rees, D.; Pascal, A.A.; Horton, P. Mechanism of ΔpH-Dependent Dissipation of Absorbed Excitation Energy by Photosynthetic Membranes. II. The Relationship between LHCII Aggregation In Vitro and QE in Isolated Thylakoids. Biochim. Biophys. Acta BBA Bioenerg. 1992, 1102, 39–44. [Google Scholar] [CrossRef]
- Ruban, A.V.; Johnson, M.P.; Duffy, C.D.P. The Photoprotective Molecular Switch in the Photosystem II Antenna. Biochim. Biophys. Acta BBA Bioenerg. 2012, 1817, 167–181. [Google Scholar] [CrossRef]
- Zito, F. The Qo Site of Cytochrome b6f Complexes Controls the Activation of the LHCII Kinase. EMBO J. 1999, 18, 2961–2969. [Google Scholar] [CrossRef]
- Vener, A.V. Environmentally Modulated Phosphorylation and Dynamics of Proteins in Photosynthetic Membranes. Biochim. Biophys. Acta BBA Bioenerg. 2007, 1767, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Sterling, D.; Alvarez, B.V.; Casey, J.R. The Extracellular Component of a Transport Metabolon. J. Biol. Chem. 2002, 277, 25239–25246. [Google Scholar] [CrossRef]
- Bruce, L.J. A Band 3-Based Macrocomplex of Integral and Peripheral Proteins in the RBC Membrane. Blood 2003, 101, 4180–4188. [Google Scholar] [CrossRef]
- Wang, C.; Yamamoto, H.; Narumiya, F.; Munekage, Y.N.; Finazzi, G.; Szabo, I.; Shikanai, T. Fine-Tuned Regulation of the K+/H+ Antiporter KEA 3 Is Required to Optimize Photosynthesis during Induction. Plant J. 2017, 89, 540–553. [Google Scholar] [CrossRef]
- Borisova, M.M.M.; Kozuleva, M.A.; Rudenko, N.N.; Naydov, I.A.; Klenina, I.B.; Ivanov, B.N. Photosynthetic Electron Flow to Oxygen and Diffusion of Hydrogen Peroxide through the Chloroplast Envelope via Aquaporins. Biochim. Biophys. Acta BBA Bioenerg. 2012, 1817, 1314–1321. [Google Scholar] [CrossRef] [PubMed]
- Perez-Martin, A.; Michelazzo, C.; Torres-Ruiz, J.M.; Flexas, J.; Fernández, J.E.; Sebastiani, L.; Diaz-Espejo, A. Regulation of Photosynthesis and Stomatal and Mesophyll Conductance under Water Stress and Recovery in Olive Trees: Correlation with Gene Expression of Carbonic Anhydrase and Aquaporins. J. Exp. Bot. 2014, 65, 3143–3156. [Google Scholar] [CrossRef]
- Wang, C.; Shikanai, T. Modification of Activity of the Thylakoid H + /K + Antiporter KEA3 Disturbs ∆pH-Dependent Regulation of Photosynthesis. Plant Physiol. 2019, 181, 762–773. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plants | The Electron Transfer Rate, µmol O2/mg Chl × h |
|---|---|
| WT | 13.7 ± 1.4 |
| α-CA2-KO (9–11) | 16.8 ± 0.9 ** |
| α-CA2-KO (8–3) | 18.0 ± 0.8 * |
| Plants | PSI/PSII Ratio at the 77 K Emission Spectra After Dark Adaptation | PSI/PSII Ratio at the 77 K Emission Spectra After 20 min Illumination with Light (λ = 640 nm) |
|---|---|---|
| WT | 3.6 ± 0.2 (100%) | 4.4 ± 0.1 (122%) |
| α-CA2-KO (9–11) | 3.0 ± 0.2 (100%) | 3.4 ± 0.1 (113%) |
| α-CA2-KO (8–3) | 3.6 ± 0.1(100%) | 3.9 ± 0.1 (108%) |
| Plants | Starch Content (mg g−1 Fresh Weight) |
|---|---|
| WT | 2.05 ± 0.13 |
| α-CA2-KO (9–11) | 1.48 ± 0.13 |
| α-CA2-KO (8–3) | 0.92 ± 0.10 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nadeeva, E.M.; Ignatova, L.K.; Rudenko, N.N.; Vetoshkina, D.V.; Naydov, I.A.; Kozuleva, M.A.; Ivanov, B.N. Features of Photosynthesis in Arabidopsis thaliana Plants with Knocked Out Gene of Alpha Carbonic Anhydrase 2. Plants 2023, 12, 1763. https://doi.org/10.3390/plants12091763
Nadeeva EM, Ignatova LK, Rudenko NN, Vetoshkina DV, Naydov IA, Kozuleva MA, Ivanov BN. Features of Photosynthesis in Arabidopsis thaliana Plants with Knocked Out Gene of Alpha Carbonic Anhydrase 2. Plants. 2023; 12(9):1763. https://doi.org/10.3390/plants12091763
Chicago/Turabian StyleNadeeva, Elena M., Lyudmila K. Ignatova, Natalia N. Rudenko, Daria V. Vetoshkina, Ilya A. Naydov, Marina A. Kozuleva, and Boris N. Ivanov. 2023. "Features of Photosynthesis in Arabidopsis thaliana Plants with Knocked Out Gene of Alpha Carbonic Anhydrase 2" Plants 12, no. 9: 1763. https://doi.org/10.3390/plants12091763