
Moderate Salinity Stress Increases the Seedling Biomass in Oilseed Rape (Brassica napus L.)
 and
and
Abstract
:1. Introduction
2. Results
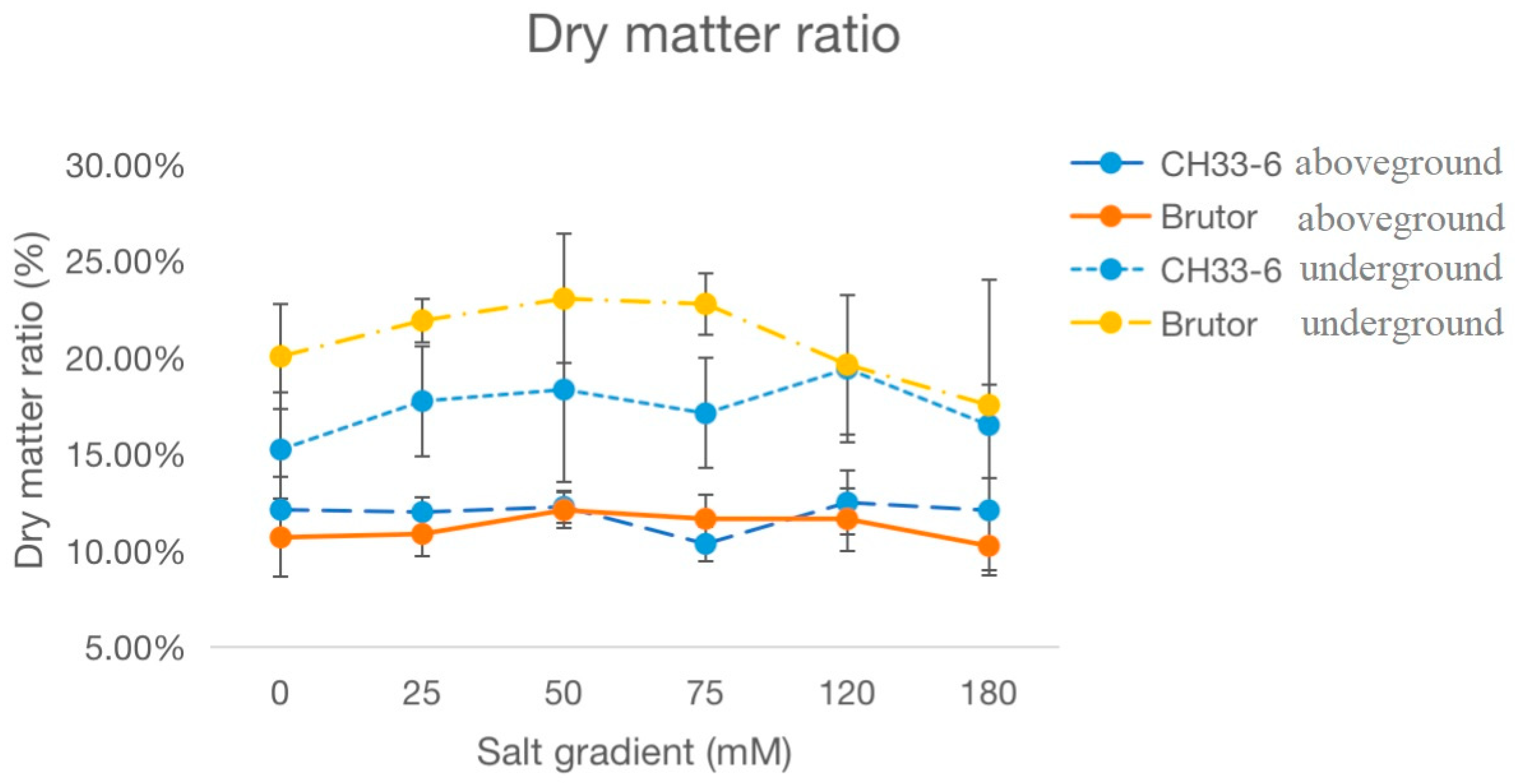
2.1. Response of Oilseed Rape Seedlings to the Treatments of Different Salinity Levels
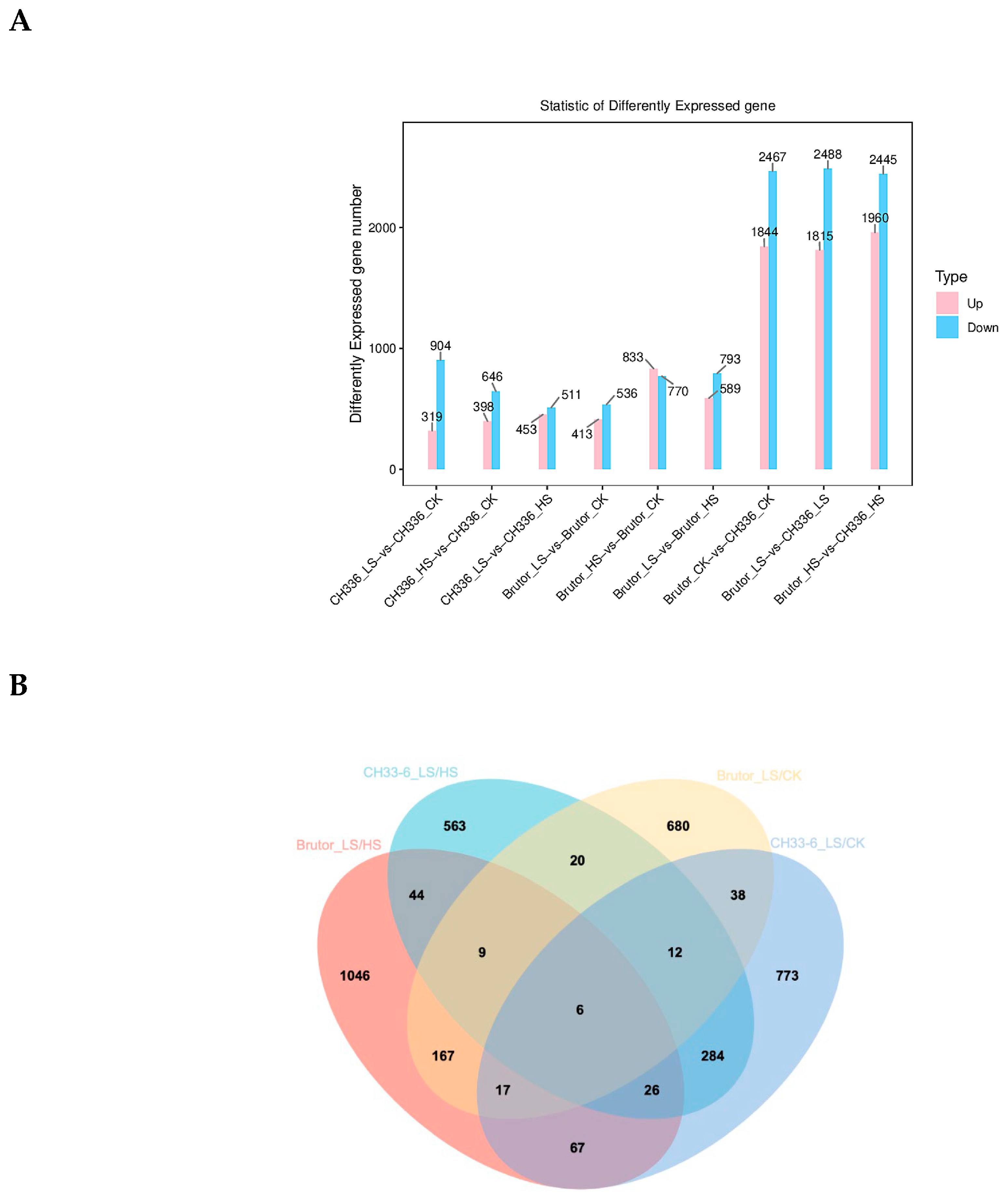
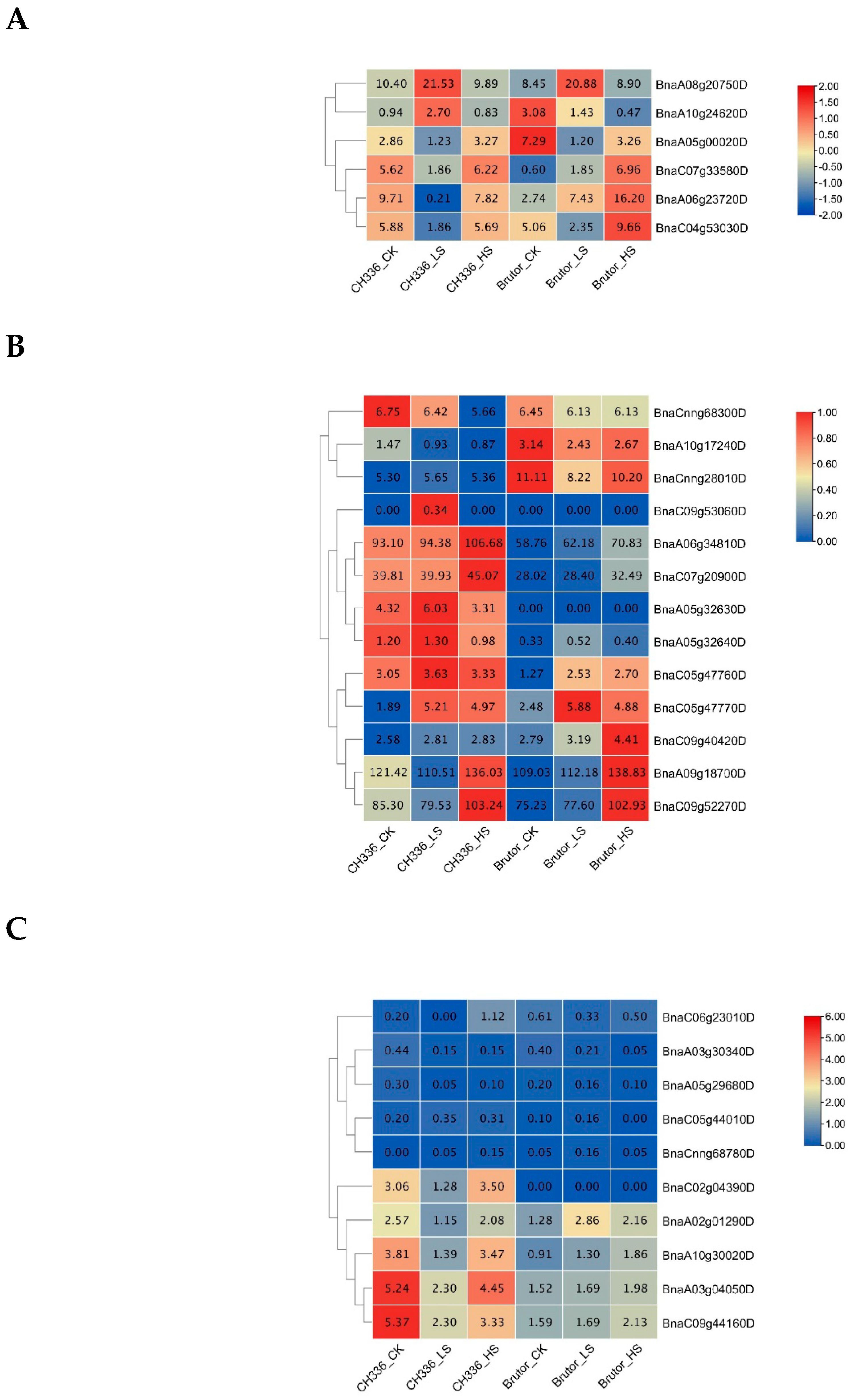
2.2. RNA-seq Analysis and Identification of Differently Expressed Genes (DEGs)
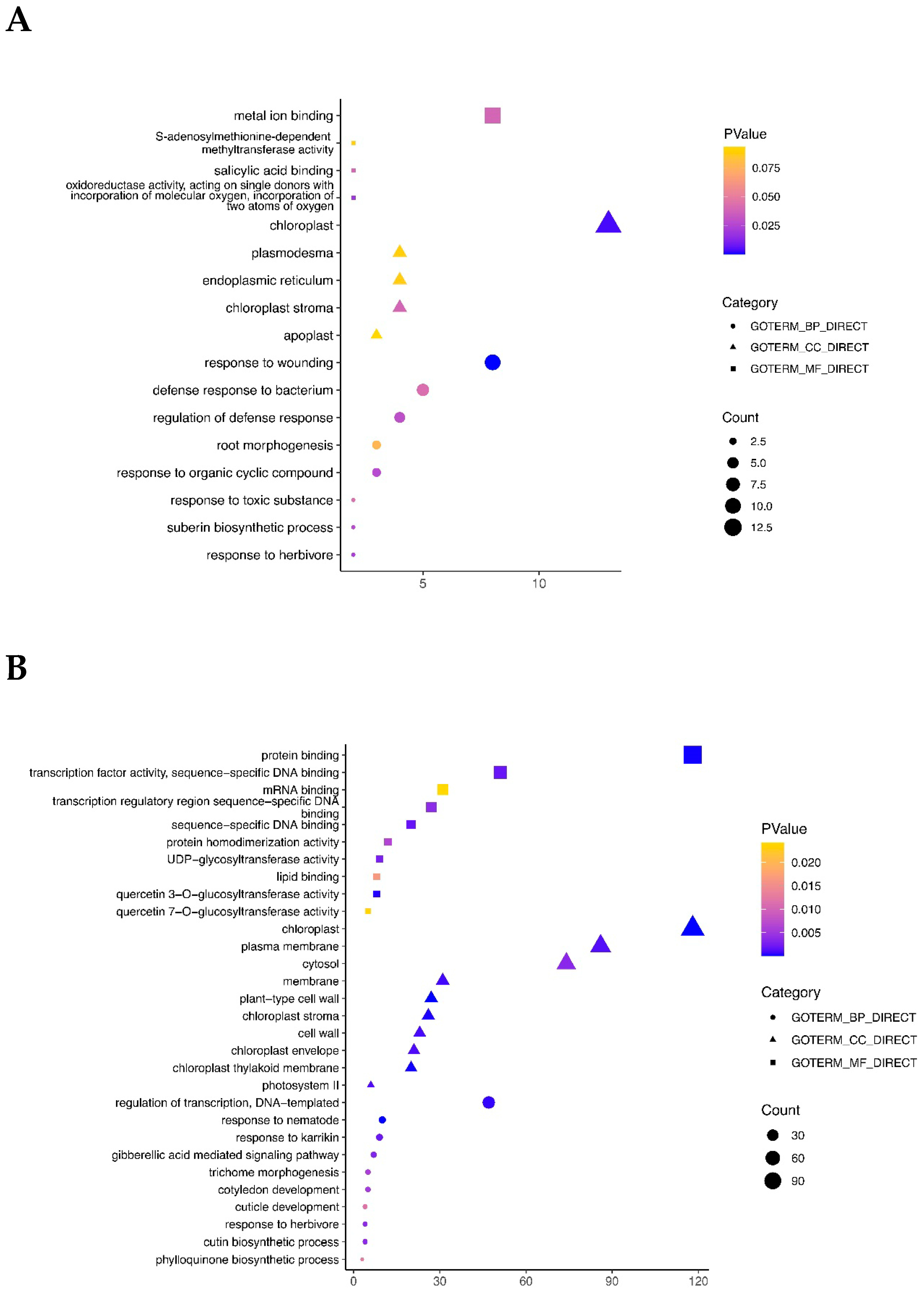
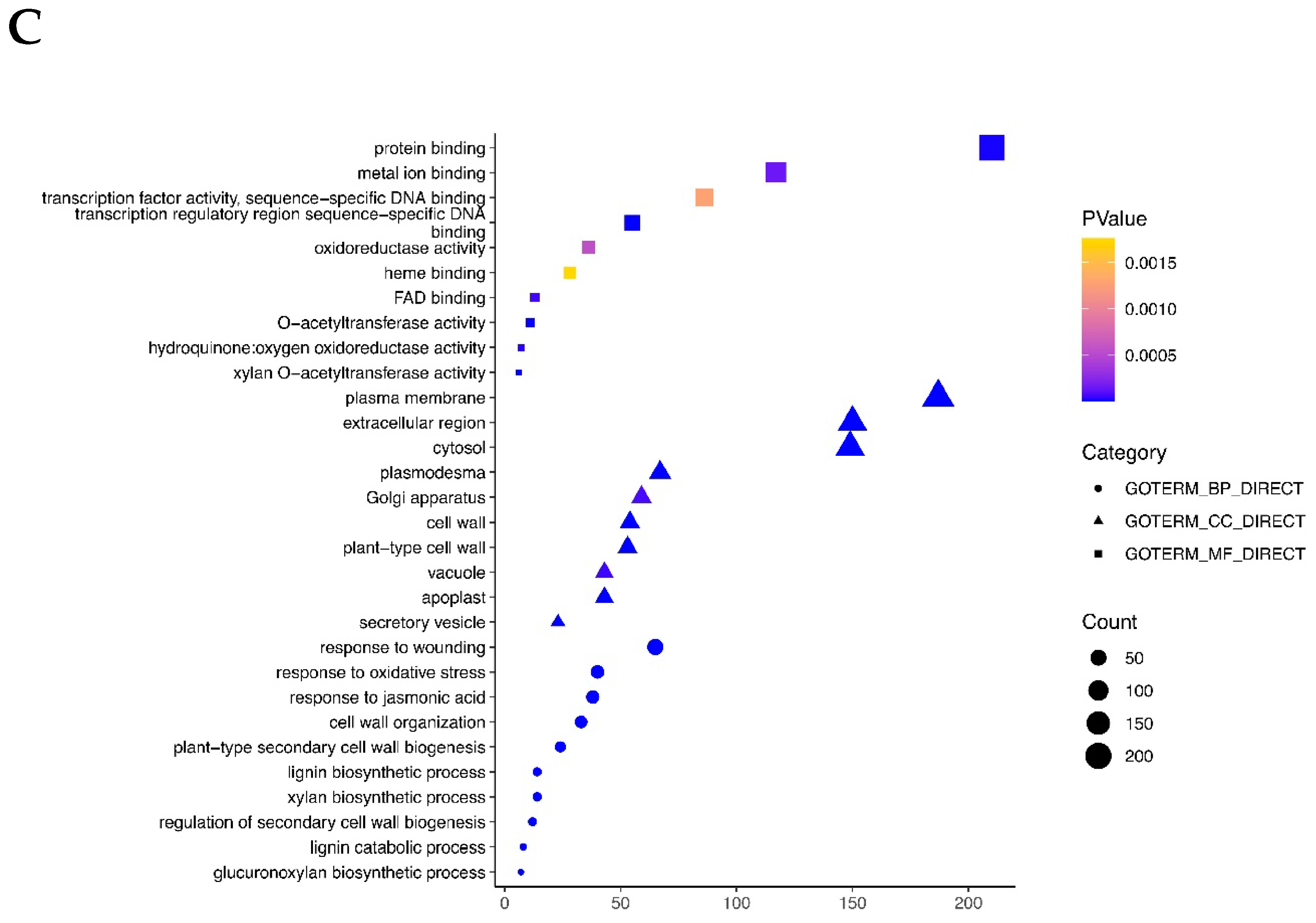
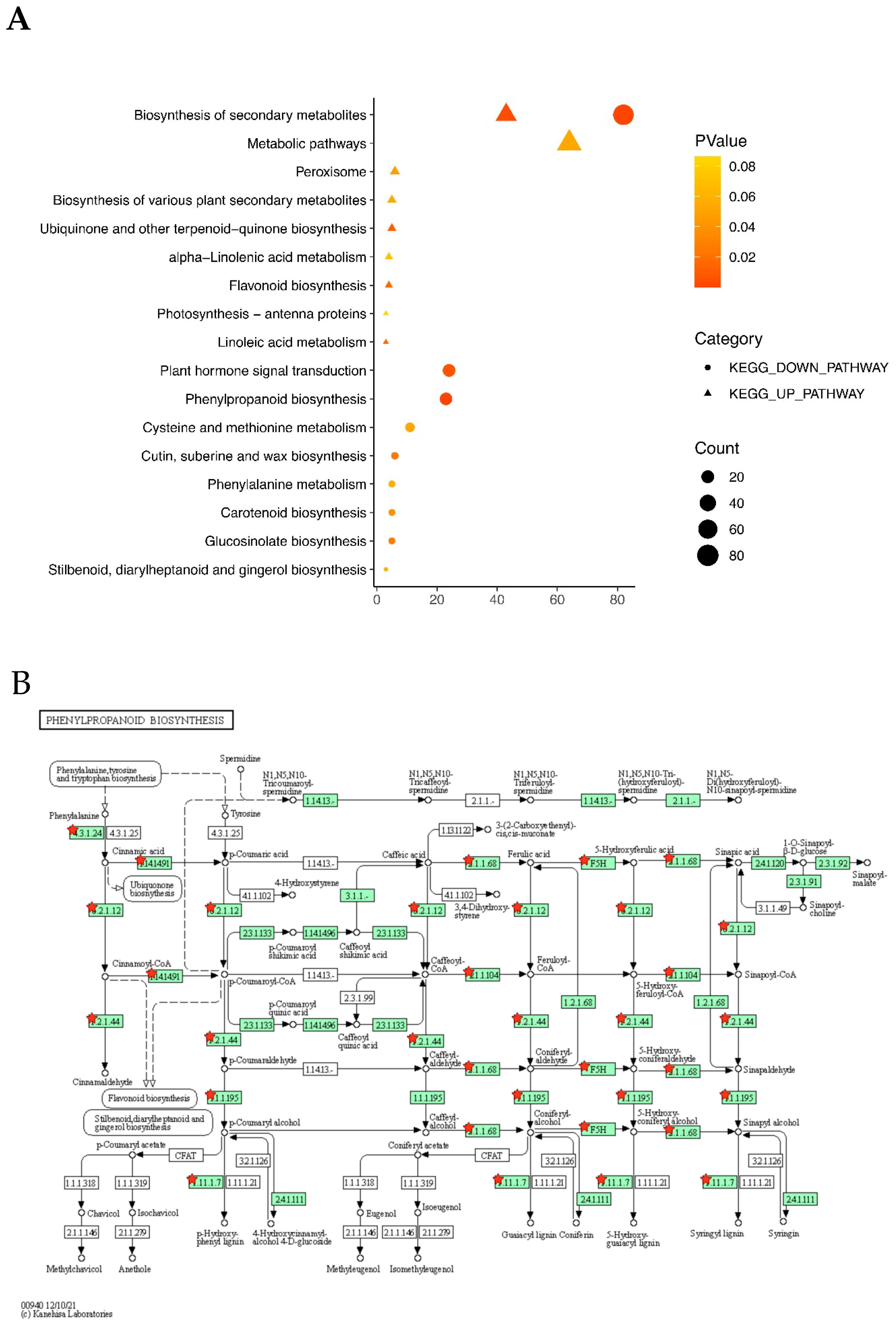
2.3. GO and KEGG Enrichment Analyses of DEGs
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Salt Solution Treatments
4.3. Measurements of Plant Growth Parameters
4.4. RNA Isolation
4.5. Construction of cDNA Libraries and RNA Sequencing
4.6. Identification of Differentially Expressed Genes (DEGs) and Functional Analysis
4.7. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, A. Soil Salinity: A Global Threat to Sustainable Development. Soil Use Manag. 2022, 38, 39–67. [Google Scholar] [CrossRef]
- Haj-Amor, Z.; Araya, T.; Kim, D.G.; Bouri, S.; Lee, J.; Ghiloufi, W.; Yang, Y.; Kang, H.; Jhariya, M.K.; Banerjee, A.; et al. Soil Salinity and Its Associated Effects on Soil Microorganisms, Greenhouse Gas Emissions, Crop Yield, Biodiversity and Desertification: A Review. Sci. Total Environ. 2022, 843, 156946. [Google Scholar] [CrossRef]
- Hopmans, J.W.; Qureshi, A.S.; Kisekka, I.; Munns, R.; Taleisnik, E. Critical Knowledge Gaps and Research Priorities in Global Soil Salinity. Adv. Agron. 2021, 9, 1–191. [Google Scholar]
- Singh, A. Alternative Management Options for Irrigation-Induced Salinization and Waterlogging Under Different Climatic Conditions. Ecol. Indic. 2018, 90, 184–192. [Google Scholar] [CrossRef]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity Induced Physiological and Biochemical Changes in Plants: An Omic Approach Towards Salt Stress Tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef]
- Hameed, A.; Ahmed, M.Z.; Hussain, T.; Aziz, I.; Ahmad, N.; Gul, B.; Nielsen, B.L. Effects of Salinity Stress on Chloroplast Structure and Function. Cells 2021, 10, 2023. [Google Scholar] [CrossRef] [PubMed]
- Zörb, C.; Geilfus, C.M.; Dietz, K.J. Salinity and Crop Yield. Plant Biol. 2019, 21 (Suppl. 1), 31–38. [Google Scholar] [CrossRef] [PubMed]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.K.; Shabala, S. Mechanisms of Plant Responses and Adaptation to Soil Salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef]
- Rahman, M.M.; Mostofa, M.G.; Keya, S.S.; Siddiqui, M.N.; Ansary, M.M.U.; Das, A.K.; Rahman, M.A.; Tran, L.S. Adaptive Mechanisms of Halophytes and Their Potential in Improving Salinity Tolerance in Plants. Int. J. Mol. Sci. 2021, 22, 10733. [Google Scholar] [CrossRef]
- Chourasia, K.N.; More, S.J.; Kumar, A.; Kumar, D.; Singh, B.; Bhardwaj, V.; Kumar, A.; Das, S.K.; Singh, R.K.; Zinta, G.; et al. Salinity Responses and Tolerance Mechanisms in Underground Vegetable Crops: An Integrative Review. Planta 2022, 255, 68. [Google Scholar] [CrossRef] [PubMed]
- Bartels, D.; Dinakar, C. Balancing Salinity Stress Responses in Halophytes and Non-Halophytes: A Comparison between Thellungiella and Arabidopsis thaliana. Funct. Plant Biol. 2013, 40, 819–831. [Google Scholar] [CrossRef]
- Prerostova, S.; Dobrev, P.I.; Gaudinova, A.; Hosek, P.; Soudek, P.; Knirsch, V.; Vankova, R. Hormonal Dynamics During Salt Stress Responses of Salt-Sensitive Arabidopsis thaliana and Salt-Tolerant Thellungiella salsuginea. Plant Sci. 2017, 264, 188–198. [Google Scholar] [CrossRef]
- Muthuramalingam, P.; Jeyasri, R.; Rakkammal, K.; Satish, L.; Shamili, S.; Karthikeyan, A.; Valliammai, A.; Priya, A.; Selvaraj, A.; Gowri, P.; et al. Multi-Omics and Integrative Approach Towards Understanding Salinity Tolerance in Rice: A Review. Biology 2022, 11, 1022. [Google Scholar] [CrossRef]
- Abuslima, E.; Kanbar, A.; Raorane, M.L.; Eiche, E.; Junker, B.H.; Hause, B.; Riemann, M.; Nick, P. Gain Time to Adapt: How Sorghum Acquires Tolerance to Salinity. Front. Plant Sci. 2022, 13, 008172. [Google Scholar] [CrossRef]
- Trono, D.; Pecchioni, N. Candidate Genes Associated with Abiotic Stress Response in Plants as Tools to Engineer Tolerance to Drought, Salinity and Extreme Temperatures in Wheat: An Overview. Plants 2022, 11, 3358. [Google Scholar] [CrossRef]
- Mwando, E.; Angessa, T.T.; Han, Y.; Li, C. Salinity Tolerance in Barley during Germination- Homologs and Potential Genes. J. Zhejiang Univ. Scien. B 2020, 21, 93–121. [Google Scholar]
- Maryum, Z.; Luqman, T.; Nadeem, S.; Khan, S.M.U.D.; Wang, B.; Ditta, A.; Khan, M.K.R. An Overview of Salinity Stress, Mechanism of Salinity Tolerance and Strategies for Its Management in Cotton. Front. Plant Sci. 2022, 13, 907937. [Google Scholar] [CrossRef] [PubMed]
- Geng, G.; Li, R.; Stevanato, P.; Lv, C.; Lu, Z.; Yu, L.; Wang, Y. Physiological and Transcriptome Analysis of Sugar Beet Reveals Different Mechanisms of Response to Neutral Salt and Alkaline Salt Stresses. Front. Plant Sci. 2020, 11, 571864. [Google Scholar] [CrossRef] [PubMed]
- Acharya, B.R.; Sandhu, D.; Dueñas, C.; Ferreira, J.F.S.; Grover, K.K. Deciphering Molecular Mechanisms Involved in Salinity Tolerance in Guar (Cyamopsis tetragonoloba (L.) Taub.) Using Transcriptome Analyses. Plants 2022, 11, 291. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.A.; Helal, M.M.U.; Tang, M.; Hu, M.; Tong, C.; Liu, S. High-Throughput Association Mapping in Brassica napus L.: Methods and Applications. Methods Mol. Biol. 2023, 2638, 67–91. [Google Scholar] [PubMed]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Early Allopolyploid Evolution in the Post-Neolithic Brassica napus Oilseed Genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [Green Version]
- Sun, F.; Fan, G.; Hu, Q.; Zhou, Y.; Guan, M.; Tong, C.; Li, J.; Du, D.; Qi, C.; Jiang, L.; et al. The High-Quality Genome of Brassica napus Cultivar ’ZS11’ Reveals the Introgression History in Semi-Winter Morphotype. Plant J. 2017, 92, 452–468. [Google Scholar] [CrossRef] [Green Version]
- Song, J.M.; Guan, Z.; Hu, J.; Guo, C.; Yang, Z.; Wang, S.; Liu, D.; Wang, B.; Lu, S.; Zhou, R.; et al. Eight High-Quality Genomes Reveal Pan-Genome Architecture and Ecotype Differentiation of Brassica napus. Nat. Plants 2020, 6, 34–45. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Liang, Z.; Yan, T.; Xu, Y.; Xuan, L.; Tang, J.; Zhou, G.; Lohwasser, U.; Hua, S.; Wang, H.; et al. Whole-Genome Resequencing of a Worldwide Collection of Rapeseed Accessions Reveals the Genetic Basis of Ecotype Divergence. Mol. Plant 2019, 12, 30–43. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Li, F.; Xu, K.; Gao, G.; Chen, B.; Yan, G.; Wang, N.; Qiao, J.; Li, J.; Li, H.; et al. Assessing and Broadening Genetic Diversity of a Rapeseed Germplasm Collection. Breed Sci. 2014, 64, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Hu, Q.; Hua, W.; Yin, Y.; Zhang, X.; Liu, L.; Shi, J.; Zhao, Y.; Qin, L.; Chen, C.; Wang, H. Rapeseed Research and Production in China. Crop J. 2017, 5, 127–135. [Google Scholar] [CrossRef] [Green Version]
- Raza, A. Eco-Physiological and Biochemical Responses of Rapeseed (Brassica napus L.) to Abiotic Stresses: Consequences and Mitigation Strategies. J. Plant Growth Regul. 2021, 40, 1368–1388. [Google Scholar] [CrossRef]
- Shahzad, B.; Rehman, A.; Tanveer, M.; Wang, L.; Park, S.K.; Ali, A. Salt Stress in Brassica: Effects, Tolerance Mechanisms, and Management. J. Plant Growth Regul. 2022, 41, 781–795. [Google Scholar] [CrossRef]
- Naeini, Z. Effects of Salinity Stress on the Morphology and Yield of Two Cultivars of Canola (Brassica napus L.). J. Agron. 2007, 6, 409–414. [Google Scholar]
- Arif, M.R.; Islam, M.T.; Robin, A.H.K. Salinity Stress Alters Root Morphology and Root Hair Traits in Brassica napus. Plants 2019, 8, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohamed, I.A.A.; Shalby, N.; Bai, C.; Qin, M.; Agami, R.A.; Jie, K.; Wang, B.; Zhou, G. Stomatal and Photosynthetic Traits Are Associated with Investigating Sodium Chloride Tolerance of Brassica Napus L. Cultivars. Plants 2020, 9, 62. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, I.A.A.; Shalby, N.; El-Badri, A.M.; Batool, M.; Wang, C.; Wang, Z.; Salah, A.; Rady, M.M.; Jie, K.; Wang, B.; et al. RNA-Seq Analysis Revealed Key Genes Associated with Salt Tolerance in Rapeseed Germination through Carbohydrate Metabolism, Hormone, and MAPK Signaling Pathways. Ind. Crops Prod. 2022, 176, 114262–114280. [Google Scholar] [CrossRef]
- Yong, H.Y.; Wang, C.; Bancroft, I.; Li, F.; Wu, X.; Kitashiba, H.; Nishio, T. Identification of a Gene Controlling Variation in the Salt Tolerance of Rapeseed (Brassica napus L.). Planta 2015, 242, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Dolatabadi, N.; Toorchi, M. Rapeseed (Brassica napus L.) Genotypes Response to NaCl Salinity. J. Biodiv. Environ. Sci. 2017, 10, 2220–6663. [Google Scholar]
- Wan, H.; Chen, L.; Guo, J.; Li, Q.; Wen, J.; Yi, B.; Ma, C.; Tu, J.; Fu, T.; Shen, J. Genome-Wide Association Study Reveals the Genetic Architecture Underlying Salt Tolerance-Related Traits in Rapeseed (Brassica napus L). Front. Plant Sci. 2017, 8, 593. [Google Scholar] [CrossRef] [Green Version]
- Wan, H.; Wei, Y.; Qian, J.; Gao, Y.; Wen, J.; Yi, B.; Ma, C.; Tu, J.; Fu, T.; Shen, J. Association Mapping of Salt Tolerance Traits at Germination Stage of Rapeseed (Brassica napus L). Euphytica 2018, 214, 190. [Google Scholar] [CrossRef]
- Wu, H.; Guo, J.; Wang, C.; Li, K.; Zhang, X.; Yang, Z.; Li, M.; Wang, B. An Effective Screening Method and a Reliable Screening Trait for Salt Tolerance of Brassica Napus at the Germination Stage. Front. Plant Sci. 2019, 10, 530. [Google Scholar] [CrossRef]
- Wassan, G.M.; Khanzada, H.; Zhou, Q.; Mason, A.S.; Keerio, A.A.; Khanzada, S.; Solangi, A.M.; Faheem, M.; Fu, D.; He, H. Identification of Genetic Variation for Salt Tolerance in Brassica napus Using Genome-Wide Association Mapping. Mol. Genet. Genom. 2021, 296, 391–408. [Google Scholar] [CrossRef]
- Davière, J.M.; Achard, P. A pivotal role of DELLAs in regulating multiple hormone signals. Mol. Plant. 2016, 9, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Novaes, E.; Kirst, M.; Chiang, V.; Winter-Sederoff, H.; Sederoff, R. Lignin and Biomass: A Negative Correlation for Wood Formation and Lignin Content in Trees. Plant Physiol. 2010, 154, 555–561. [Google Scholar] [CrossRef] [Green Version]
- Dong, N.; Lin, H. Contribution of Phenylpropanoid Metabolism to Plant Development and Plant–Environment Interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef]
- Cho, J.S.; Jeon, H.W.; Kim, M.H.; Vo, T.K.; Kim, J.; Park, E.J.; Choi, Y.I.; Lee, H.; Han, K.H.; Ko, J.H. Wood Forming Tissue-Specific Bicistronic Expression of Pdga20ox1 and Ptrmyb221 Improves Both the Quality and Quantity of Woody Biomass Production in a Hybrid Poplar. Plant Biotechnol. J. 2019, 17, 1048–1057. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Hernández, M.D.C.; Garmendia, I. Optimum Growth and Quality of the Edible Ice Plant Under Saline Conditions. J. Sci. Food Agric. 2022, 102, 2686–2692. [Google Scholar] [CrossRef] [PubMed]
- Niinemets, Ü. Components of Leaf Dry Mass per Area-Thickness and Density-Alter Leaf Photosynthetic Capacity in Reverse Directions in Woody Plants. New Phytol. 1999, 144, 35–47. [Google Scholar] [CrossRef]
- Cheng, J.F.; Dai, T.B.; Jiang, H.Y.; Pan, X.Y.; Cao, W.X. Characterization of Leaf Carbon and Nitrogen Assimilation in Different Rice Genotypes at Jointing Stage and Their Relationships with Nitrogen Utilization Efficiency. Chin. J. Rice Sci. 2012, 26, 101–108. [Google Scholar]
- Rosati, A.; Esparza, G.; DeJong, T.M.; Pearcy, R.W. Influence of Canopy Light Environment and Nitrogen Availability on Leaf Photosynthetic Characteristics and Photosynthetic Nitrogen-Use Efficiency of Field-Grown Nectarine Trees. Tree Physiol. 1999, 19, 173–180. [Google Scholar] [CrossRef]
- Meurer, J.; Schmid, L.M.; Stoppel, R.; Leister, D.; Brachmann, A.; Manavski, N. PALE CRESS Binds to Plastid Rnas and Facilitates the Biogenesis of the 50S Ribosomal Subunit. Plant J. 2017, 92, 400–413. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Lu, S.; Zhuang, M.; Zhang, Y.; Lv, H.; Ji, J.; Hou, X.; Fang, Z.; Wang, Y.; Yang, L. Genome-Wide Identification and Expression Analysis of the Brassica Oleracea L. Chitin-Binding Genes and Response to Pathogens Infections. Planta 2021, 253, 80. [Google Scholar] [CrossRef] [PubMed]
- Hanfrey, C.; Fife, M.; Buchanan-Wollaston, V. Leaf Senescence in Brassica napus: Expression of Genes Encoding Pathogenesis-Related Proteins. Plant Mol. Biol. 1996, 30, 597–609. [Google Scholar] [CrossRef]
- Kwon, S.I.; Cho, H.J.; Jung, J.H.; Yoshimoto, K.; Shirasu, K.; Park, O.K. The Rab GTPase RabG3b Functions in Autophagy and Contributes to Tracheary Element Differentiation in Arabidopsis. Plant J. 2010, 64, 151–164. [Google Scholar] [CrossRef]
- Kwon, S.I.; Cho, H.J.; Lee, J.S.; Jin, H.; Shin, S.J.; Kwon, M.; Noh, E.W.; Park, O.K. Overexpression of Constitutively Active Arabidopsis Rabg3b Promotes Xylem Development in Transgenic Poplars. Plant Cell Environ. 2011, 34, 2212–2224. [Google Scholar] [CrossRef] [PubMed]
- Colebrook, E.H.; Thomas, S.G.; Phillips, A.L.; Hedden, P. The Role of Gibberellin Signalling in Plant Responses to Abiotic Stress. J. Exp. Biol. 2014, 217, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Achard, P.; Gusti, A.; Cheminant, S.; Alioua, M.; Dhondt, S.; Coppens, F.; Beemster, G.T.; Genschik, P. Gibberellin Signaling Controls Cell Proliferation Rate in Arabidopsis. Curr. Biol. 2009, 19, 1188–1193. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, Y.; Hu, Y.; Liu, H.; He, M.; Yang, Z.; Kong, F.; Liu, X.; Hou, X. DELLA and EDS1 Form a Feedback Regulatory Module to Fine-Tune Plant Growth-Defense Tradeoff in Arabidopsis. Mol. Plant. 2019, 12, 1485–1498. [Google Scholar] [CrossRef]
- Dill, A.; Sun, T.P. Synergistic Derepression of Gibberellin Signaling by Removing RGA and GAI Function in Arabidopsis thaliana. Genetics 2001, 159, 777–785. [Google Scholar] [CrossRef]
- King, K.E.; Moritz, T.; Harberd, N.P. Gibberellins are not Required for Normal Stem Growth in Arabidopsis thaliana in the Absence of GAI and RGA. Genetics 2001, 159, 767–776. [Google Scholar] [CrossRef]
- Chen, L.; Xiang, S.; Chen, Y.; Li, D.; Yu, D. Arabidopsis WRKY45 Interacts with the DELLA Protein RGL1 to Positively Regulate Age-Triggered Leaf Senescence. Mol. Plant. 2017, 10, 1174–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, M.D.; Barro-Trastoy, D.; Fuster-Almunia, C.; Tornero, P.; Alonso, J.M.; Perez-Amador, M.A. Gibberellin-Mediated RGA-LIKE1 Degradation Regulates Embryo Sac Development in Arabidopsis. J. Exp. Bot. 2020, 71, 7059–7072. [Google Scholar] [CrossRef]
- Lee, S.C.; Cheng, H.; King, K.E.; Wang, W.F.; He, Y.W.; Hussain, A.; Lo, J.; Harberd, N.P.; Peng, J.R. Gibberellin Regulates Arabidopsis Seed Germination via RGL2, a GAI/RGA-like Gene whose Expression is up-Regulated Following Imbibition. Gene Dev. 2002, 16, 646–658. [Google Scholar] [CrossRef] [Green Version]
- Ravindran, P.; Verma, V.; Stamm, P.; Kumar, P.P. A novel RGL2–DOF6 Complex Contributes to Primary Seed Dormancy in Arabidopsis thaliana by Regulating a GATA Transcription Factor. Mol. Plant. 2017, 10, 1307–1320. [Google Scholar] [CrossRef] [Green Version]
- Wild, M.; Achard, P. The DELLA protein RGL3 Positively Contributes to Jasmonate/Ethylene Defense Responses. Plant Signal. Behav. 2014, 8, e23891. [Google Scholar] [CrossRef] [PubMed]
- Gray, J.; Caparrós-Ruiz, D.; Grotewold, E. Grass Phenylpropanoids: Regulate before Using! Plant Sci. 2012, 184, 112–120. [Google Scholar] [CrossRef]
- Lanot, A.; Hodge, D.; Lim, E.K.; Vaistij, F.E.; Bowles, D.J. Redirection of Flux Through the Phenylpropanoid Pathway by Increased Glucosylation of Soluble Intermediates. Planta 2008, 228, 609–616. [Google Scholar] [CrossRef]
- Zhong, R.; Lee, C.; Ye, Z.H. Evolutionary Conservation of the Transcriptional Network Regulating Secondary Cell Wall Biosynthesis. Trends Plant Sci. 2010, 15, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.C.; Ko, J.H.; Kim, J.Y.; Kim, J.; Bae, H.J.; Han, K.H. MYB46 Directly Regulates the Gene Expression of Secondary Wall-Associated Cellulose Synthases in Arabidopsis. Plant J. 2013, 73, 26–36. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A Fast Spliced Aligner with Low Memory Requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript Assembly and Quantification by RNA-Seq Reveals Unannotated Transcripts and Isoform Switching During Cell Differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq--a Python Framework to Work with High-Throughput Sequencing Data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [Green Version]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for Linking Genomes to Life and the Environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Salinity (mmol L−1) | Plant Height at Six-Leaf Stage (cm) | Aboveground Fresh Weight at Early Flowering Stage (g) | Underground Fresh Weight at Early Flowering Stage (g) | Aboveground Dry Weight at Early Flowering Stage (g) | Underground Dry Weight at Early Flowering Stage (g) | Specific Leaf Weight (SLW) at Early Flowering Stage (µg/cm2) |
|---|---|---|---|---|---|---|---|
| CH336 | 0 | 32.7 ± 1.4 b | 244.39 ± 14.27 a | 16.23 ± 0.99 b | 29.47 ± 1.94 ab | 2.51 ± 0.26 b | 4.89 ± 0.37 a |
| 25 | 43.7 ± 2.0 a | 281.75 ± 15.70 a | 19.37 ± 1.08 ab | 33.64 ± 1.56 a | 3.46 ± 0.30 ab | 4.94 ± 0.27 a | |
| 50 | 42.0 ± 1.2 a | 286.37 ± 12.88 a | 20.90 ± 1.40 a | 35.30 ± 2.05 a | 3.73 ± 0.25 a | 5.41 ± 0.10 a | |
| 75 | 35.1 ± 1.4 b | 235.81 ± 12.69 ab | 14.94 ± 0.70 bc | 24.27 ± 1.08 b | 2.53 ± 0.14 b | 5.13 ± 0.91 a | |
| 120 | 34.4 ± 0.8 b | 185.27 ± 8.88 b | 14.54 ± 0.98 bc | 23.22 ± 1.75 bc | 2.83 ± 0.27 ab | 5.32 ± 0.47 a | |
| 180 | 24.0 ± 1.6 c | 133.53 ± 5.10 c | 10.75 ± 0.87 c | 16.33 ± 1.29 c | 1.82 ± 0.30 b | 5.16 ± 0.32 a | |
| Brutor | 0 | 44.9 ± 1.2 bc | 646.58 ± 32.05 ab | 46.21 ± 2.86 b | 69.02 ± 3.44 ab | 8.92 ± 0.77 b | 3.76 ± 0.53 a |
| 25 | 60.2 ± 2.6 a | 700.47 ± 45.55 a | 59.51 ± 4.63 a | 74.96 ± 4.49 ab | 12.44 ± 1.24 ab | 3.99 ± 0.40 a | |
| 50 | 47.9 ± 1.2 b | 691.86 ± 44.52 a | 66.16 ± 3.34 a | 82.90 ± 5.34 a | 14.70 ± 0.82 a | 4.14 ± 0.63 a | |
| 75 | 46.6 ± 1.5 bc | 612.30 ± 31.61 ab | 56.14 ± 2.41 ab | 71.09 ± 3.77 ab | 13.11 ± 0.65 ab | 4.49 ± 0.80 a | |
| 120 | 39.6 ± 1.4 c | 545.23 ± 21.52 b | 43.67 ± 2.82 b | 64.09 ± 4.92 b | 10.15 ± 0.79 b | 4.46 ± 0.12 a | |
| 180 | 38.3 ± 0.7 c | 436.96 ± 13.67 b | 28.13 ± 0.95 c | 44.56 ± 1.54 c | 5.17 ± 0.35 c | 3.90 ± 0.36 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, B.; Bian, X.; Tu, M.; Yu, T.; Jiang, L.; Lu, Y.; Chen, X. Moderate Salinity Stress Increases the Seedling Biomass in Oilseed Rape (Brassica napus L.). Plants 2023, 12, 1650. https://doi.org/10.3390/plants12081650
Chen B, Bian X, Tu M, Yu T, Jiang L, Lu Y, Chen X. Moderate Salinity Stress Increases the Seedling Biomass in Oilseed Rape (Brassica napus L.). Plants. 2023; 12(8):1650. https://doi.org/10.3390/plants12081650
Chicago/Turabian StyleChen, Beini, Xiaobo Bian, Mengxin Tu, Tao Yu, Lixi Jiang, Yunhai Lu, and Xiaoyang Chen. 2023. "Moderate Salinity Stress Increases the Seedling Biomass in Oilseed Rape (Brassica napus L.)" Plants 12, no. 8: 1650. https://doi.org/10.3390/plants12081650