
Pollinator Proboscis Length Plays a Key Role in Floral Integration of Honeysuckle Flowers (Lonicera spp.)


Abstract
:1. Introduction
2. Results
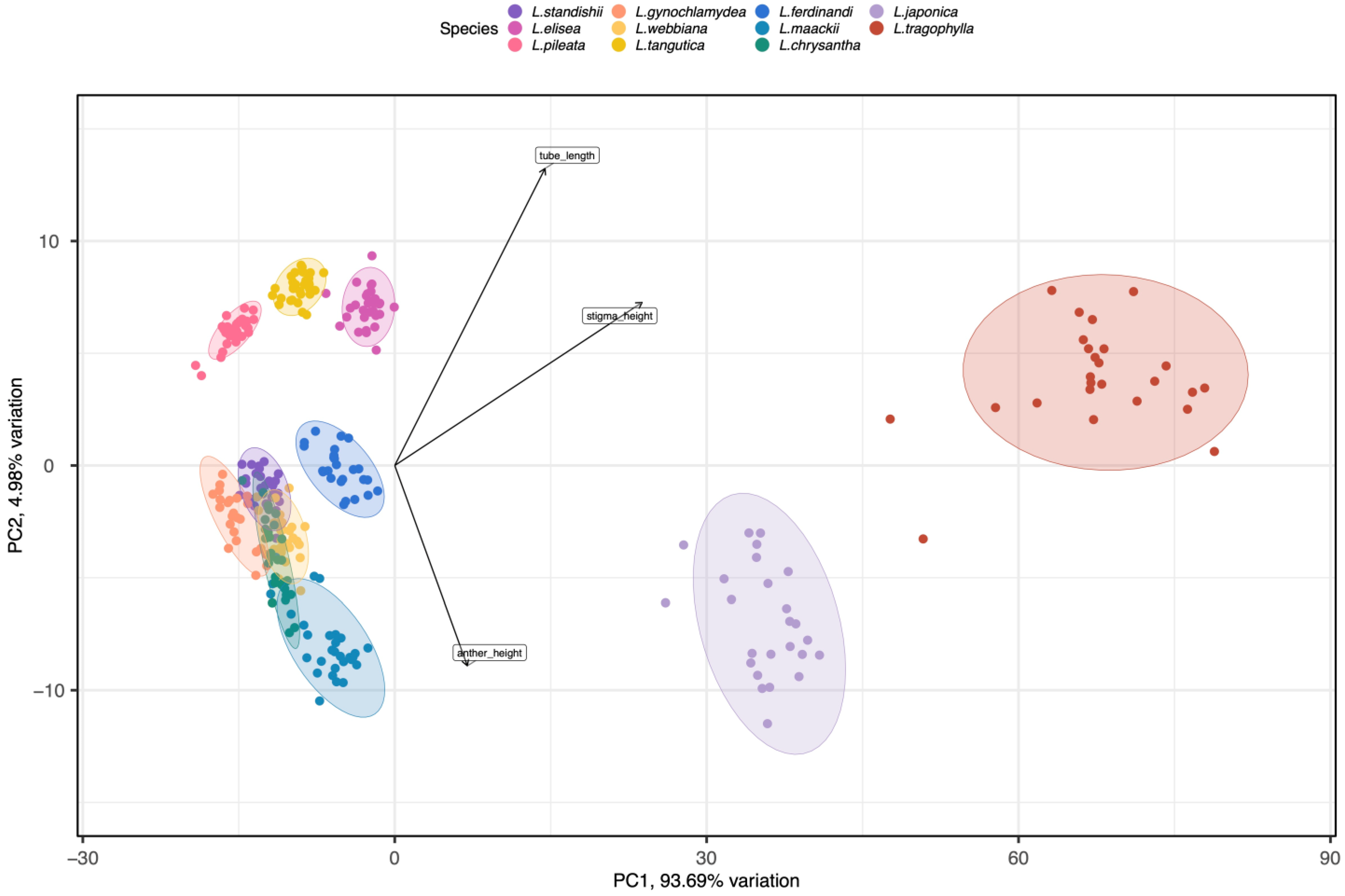
2.1. The Divergence of Floral Traits among 11 Lonicera Species
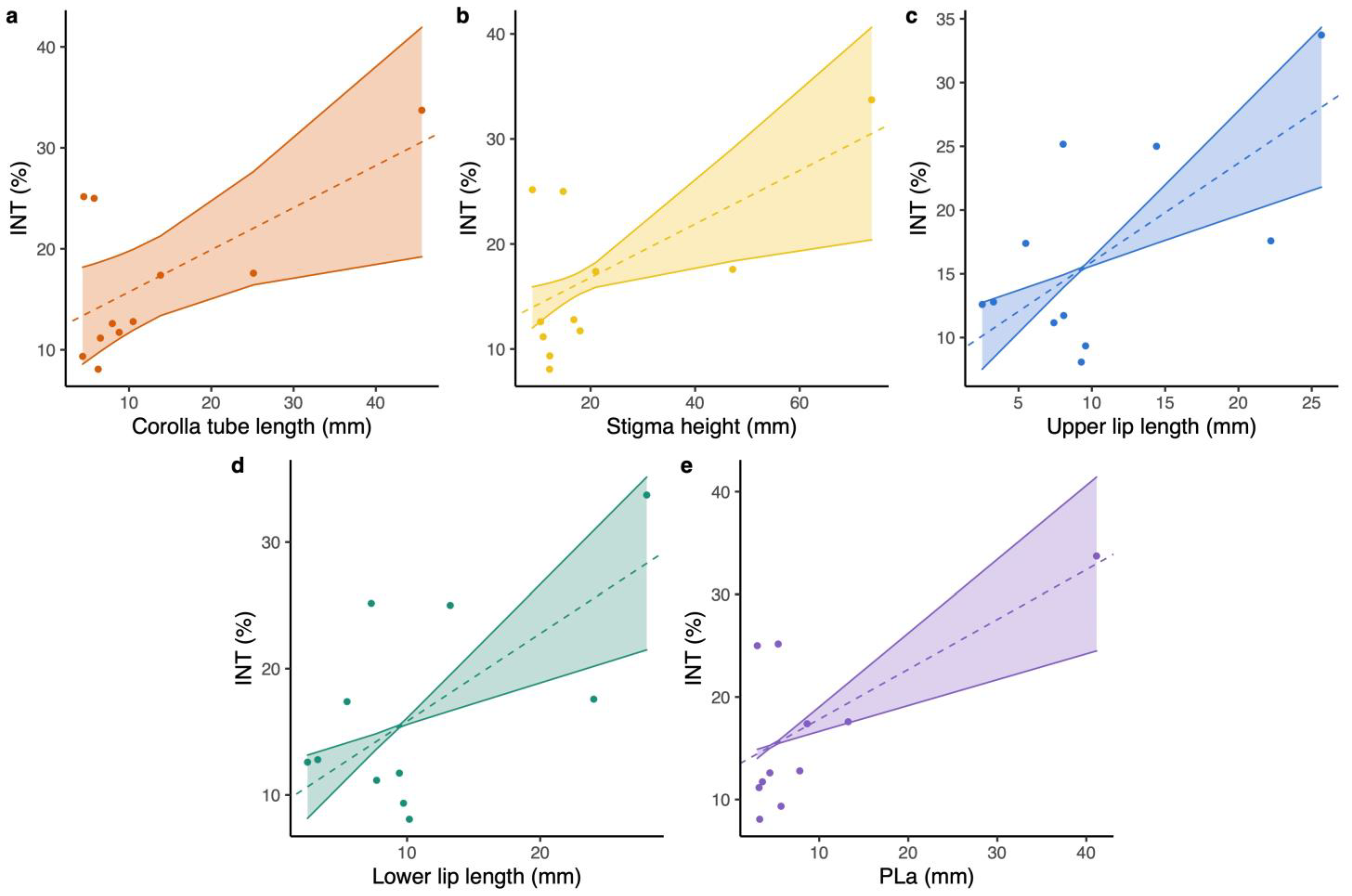
2.2. The Relationship between Floral Integration and Floral Traits and Pollinators’ Proboscis Length
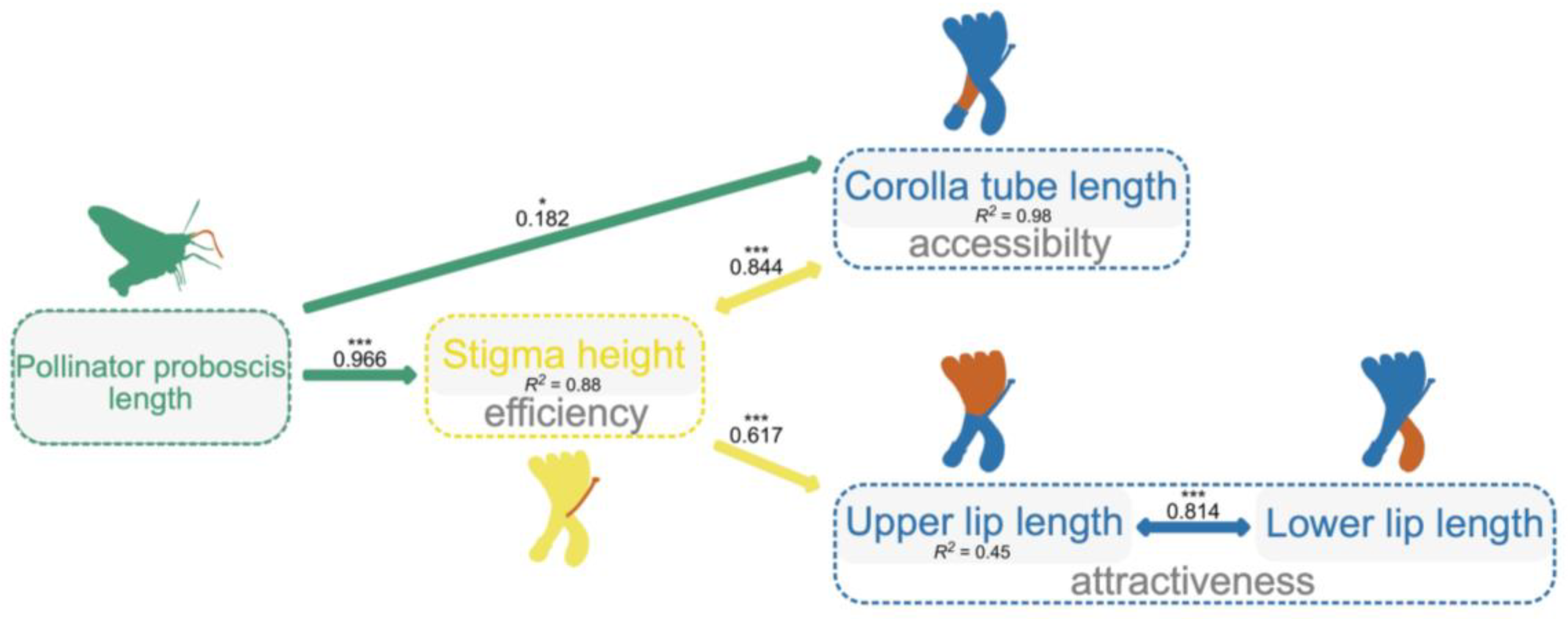
2.3. The Potential Pathway by Which Pollinators Integrate Floral Traits
3. Discussion
3.1. Influence of Pollinator Proboscis Length on Floral Integration
3.2. The Potential Pathway by Which Pollinators Integrate Floral Traits
4. Materials and Methods
4.1. Study Species and Sites
4.2. Measurements of Floral Traits and Estimation of Floral Integration
4.3. Pollinator Observation and Measurement of Pollinators’ Proboscis Length
4.4. Statistical Analyses
4.4.1. The Divergence of Floral Traits in 11 Lonicera Species
4.4.2. The Relationship of Floral Integration with Pollinator Proboscis Length and Floral Traits
4.4.3. The Potential Pathway by Which Pollinators Integrate Floral Traits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Conner, J.K.; Sterling, A. Testing hypotheses of functional relationships: A comparative survey of correlation patterns among floral traits in 5 insect-pollinated plants. Am. J. Bot. 1995, 82, 1399–1406. [Google Scholar] [CrossRef]
- Pigliucci, M. Phenotypic integration: Studying the ecology and evolution of complex phenotypes. Ecol. Lett. 2003, 6, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Ordano, M.; Fornoni, J.; Boege, K.; Domínguez, C.A. The adaptive value of phenotypic floral integration. New Phytol. 2008, 179, 1183–1192. [Google Scholar] [CrossRef]
- Gómez, J.M.; Torices, R.; Lorite, J.; Klingenberg, C.P.; Perfectti, F. The role of pollinators in the evolution of corolla shape variation, disparity and integration in a highly diversified plant family with a conserved floral bauplan. Ann. Bot. 2016, 117, 889–904. [Google Scholar] [CrossRef] [Green Version]
- Armbruster, W.S.; Pelabon, C.; Bolstad, G.H.; Hansen, T.F. Integrated phenotypes: Understanding trait covariation in plants and animals. Proc. R. Soc. B Biol. Sci. 2014, 369, 20130245. [Google Scholar] [CrossRef] [Green Version]
- Klingenberg, C.P. Studying morphological integration and modularity at multiple levels: Concepts and analysis. Phil. Trans. Biol. Sci. 2014, 369, 20130249. [Google Scholar] [CrossRef]
- Klingenberg, C.P. Morphological integration and developmental modularity. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 115–132. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.D. Pleiotropy and the evolution of floral integration. New Phytol. 2016, 209, 80–85. [Google Scholar] [CrossRef]
- Bissell, E.K.; Diggle, P.K. Modular genetic architecture of floral morphology in Nicotiana: Quantitative genetic and comparative phenotypic approaches to floral integration. J. Evol. Biol. 2010, 23, 1744–1758. [Google Scholar] [CrossRef]
- Reich, D.; Berger, A.; von Balthazar, M.; Chartier, M.; Sherafati, M.; Schönenberger, J.; Manafzade, S.; Staedler, Y.M. Modularity and evolution of flower shape: The role of function, development, and spandrels in Erica. New Phytol. 2020, 226, 267–280. [Google Scholar] [CrossRef] [Green Version]
- Lázaro, A.; Santamaría, L. Flower-visitor selection on floral integration in three contrasting populations of Lonicera implexa. Am. J. Bot. 2016, 103, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Rosas-Guerrero, V.; Quesada, M.; Armbruster, W.S.; Pérez-Barrales, R.; Smith, S.D. Influence of pollination specialization and breeding system on floral integration and phenotypic variation in Ipomoea. Evolution 2011, 65, 350–364. [Google Scholar] [CrossRef]
- Anderson, I.A.; Busch, J.W. Relaxed pollinator-mediated selection weakens floral integration in self-compatible taxa of Leaven worthia (Brassicaceae). Am. J. Bot. 2006, 93, 860–867. [Google Scholar] [CrossRef]
- Pérez-Barrales, R.; Arroyo, J.; Armbruster, W.S. Differences in pollinator faunas may generate geographic differences in floral morphology and integration in Narcissus papyraceus (Amaryllidaceae). Oikos 2007, 116, 1904–1918. [Google Scholar] [CrossRef]
- Ellis, A.G.; Brockington, S.F.; de Jager, M.L.; Mellers, G.; Walker, R.H.; Glover, B.J. Floral trait variation and integration as a function of sexual deception in Gorteria diffusa. Proc. R. Soc. B Biol. Sci. 2014, 369, 20130563. [Google Scholar] [CrossRef] [Green Version]
- Itagaki, T.; Misaki, A.; Sakai, S. Selection for floral integration and trait variation in zygomorphic flowers of Aconitum japonicum ssp. subcuneatum (Ranunculaceae). Plant Ecol. 2020, 221, 347–359. [Google Scholar] [CrossRef]
- Galen, C. Rates of floral evolution: Adaptation to bumblebee pollination in an alpine wildflower, Polemonium viscosum. Evolution 1996, 50, 120–125. [Google Scholar] [CrossRef]
- Whittall, J.B.; Hodges, S.A. Pollinator shifts drive increasingly long nectar spurs in columbine flowers. Nature 2007, 447, 706–709. [Google Scholar] [CrossRef]
- Anderson, B.; Ros, P.; Wiese, T.J.; Ellis, A.G. Intraspecific divergence and convergence of floral tube length in specialized pollination interactions. Proc. R. Soc. B Biol. Sci. 2014, 281, 20141420. [Google Scholar] [CrossRef]
- Johnson, S.D.; Moré, M.; Amorim, F.W.; Haber, W.A.; Frankie, G.W.; Stanley, D.A.; Cocucci, A.A.; Raguso, R.A. The long and the short of it: A global analysis of hawkmoth pollination niches and interaction networks. Funct. Ecol. 2017, 31, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Caruso, C.M.; Eisen, K.E.; Martin, R.A.; Sletvold, N. A meta-analysis of the agents of selection on floral traits. Evolution 2019, 73, 4–14. [Google Scholar] [CrossRef] [Green Version]
- Lázaro, A.; Totland, Ø. The influence of floral symmetry, dependence on pollinators and pollination generalization on flower size variation. Ann. Bot. 2014, 114, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Rehder, A. Synopsis of the genus Lonicera. Mo. Bot. Gard. Annu. Rep. 1903, 14, 27–232. [Google Scholar] [CrossRef]
- Van Steenis, C.G. Preliminary revision of the genus Lonicera in Malaysia. J. Arnold Arbor. 1946, 27, 442–452. [Google Scholar] [CrossRef]
- Yang, Q.; Landrein, S.; Osborne, J.; Borosova, R. Flora of China; Science Press: Beijing, China, 2011; Volume 19. [Google Scholar]
- Theis, N.; Donoghue, M.J.; Li, J. Phylogenetics of the Caprifolieae and Lonicera (Dipsacales) based on nuclear and chloroplast DNA sequences. Syst. Bot. 2008, 33, 776–783. [Google Scholar] [CrossRef]
- Bieniasz, M.; Dziedzic, E.; Kusibab, T. Evaluation of morphological traits of flowers and crossing possibility of haskap (Lonicera L.) cultivars depending on their origin. J. Berry Res. 2022, 12, 115–129. [Google Scholar] [CrossRef]
- Deng, H.; Xiang, G.J.; Guo, Y.H.; Yang, C.F. Study on the breeding system and floral color change of four Lonicera species in the Qinling mountains. Plant Sci. J. 2017, 35, 1–12. [Google Scholar]
- Pickens, A.L. Steps in the development of the bird-flower. Condor 1936, 38, 150–154. [Google Scholar] [CrossRef]
- Zhu, X.G. Pollination Biology of Four Species of Lonicera in Qinling Mountain. Master’s Thesis, Wuhan University, Wuhan, China, 2014. [Google Scholar]
- Gómez, J.M.; Perfectti, F.; Klingenberg, C.P. The role of pollinator diversity in the evolution of corolla-shape integration in a pollination-generalist plant clade. Phil. Trans. Biol. Sci. 2014, 369, 20130257. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Barrales, R.; Simón-Porcar, V.I.; Santos-Gally, R.; Arroyo, J. Phenotypic integration in style dimorphic daffodils (Narcissus, Amaryllidaceae) with different pollinators. Phil. Trans. Biol. Sci. 2014, 369, 20130258. [Google Scholar] [CrossRef] [Green Version]
- Lázaro, A.; Seguí, J.; Santamaría, L. Continuous variation in herkogamy enhances the reproductive response of Lonicera implexa to spatial variation in pollinator assemblages. AoB Plants 2020, 12, plz078. [Google Scholar] [CrossRef]
- Lázaro, A.; Vignolo, C.; Santamaría, L. Long corollas as nectar barriers in Lonicera implexa: Interactions between corolla tube length and nectar volume. Evol. Ecol. 2015, 29, 419–435. [Google Scholar] [CrossRef]
- Berg, R.L. The ecological significance of correlation pleiades. Evolution 1960, 14, 171–180. [Google Scholar] [CrossRef]
- Heywood, J.S.; Michalski, J.S.; McCann, B.K.; Russo, A.D.; Andres, K.J.; Hall, A.R.; Middleton, T.C. Genetic and environmental integration of the hawkmoth pollination syndrome in Ruellia humilis (Acanthaceae). Ann. Bot. 2017, 119, 1143–1155. [Google Scholar] [CrossRef] [Green Version]
- Beattie, A.J. Pollination mechanisms in Viola. New Phytol. 1971, 70, 343–360. [Google Scholar] [CrossRef]
- Sharma, B.; Yant, L.; Hodges, S.A.; Kramer, E.M. Understanding the development and evolution of novel floral form in Aquilegia. Curr. Opin. Plant Biol. 2014, 17, 22–27. [Google Scholar] [CrossRef]
- Toji, T.; Hirota, S.K.; Ishimoto, N.; Suyama, Y.; Itino, T. Intraspecific independent evolution of floral spur length in response to local flower visitor size in Japanese Aquilegia in different mountain regions. Ecol. Evol. 2022, 12, e8668. [Google Scholar] [CrossRef]
- Janzen, D.H. On ecological fitting. Oikos 1985, 45, 308–310. [Google Scholar] [CrossRef] [Green Version]
- Newman, E.; Manning, J.; Anderson, B. Matching floral and pollinator traits through guild convergence and pollinator ecotype formation. Ann. Bot. 2014, 113, 373–384. [Google Scholar] [CrossRef] [Green Version]
- Armbruster, W.S.; Wege, J.A. Detecting canalization and intra-floral modularity in triggerplant (Stylidium) flowers: Correlations are only part of the story. Am. J. Bot. 2019, 123, 355–372. [Google Scholar] [CrossRef] [Green Version]
- Benitez-Vieyra, S.; Fornoni, J.; Pérez-Alquicira, J.; Boege, K.; Dominguez, C.A. The evolution of signal-reward correlations in bee- and hummingbird-pollinated species of Salvia. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132934. [Google Scholar] [CrossRef] [Green Version]
- Wagner, G.P. On the eigenvalue distribution of genetic and phenotypic dispersion matrices: Evidence for a nonrandom organization of quantitative character variation. J. Math. Biol. 1984, 21, 77–95. [Google Scholar] [CrossRef]
- Cheverud, J.M.; Wagner, G.P.; Dow, M.M. Methods for the comparative analysis of variation patterns. Syst. Zool. 1989, 38, 201–213. [Google Scholar] [CrossRef]
- Torices, R.; Muñoz-Pajares, A.J. PHENIX: An R package to estimate a size-controlled phenotypic integration index. Appl. Plant Sci. 2015, 3, 1400104. [Google Scholar] [CrossRef]
- Miyake, T.; Yahara, T. Why does the flower of Lonicera japonica open at dusk? Botany 1998, 76, 1806–1811. [Google Scholar] [CrossRef]
- Gegear, R.J.; Burns, R.; Swoboda-Bhattarai, K.A. “Hummingbird” floral traits interact synergistically to discourage visitation by bumble bee foragers. Ecology 2017, 98, 489–499. [Google Scholar] [CrossRef] [Green Version]
- Vlašánková, A.; Padyšáková, E.; Bartoš, M.; Mengual, X.; Janečková, P.; Janeček, Š. The nectar spur is not only a simple specialization for long-proboscid pollinators. New Phytol. 2017, 215, 1574–1581. [Google Scholar] [CrossRef] [Green Version]
- Kulbaba, M.W.; Worley, A.C. Selection on Polemonium brandegeei (Polemoniaceae) flowers under hummingbird pollination: In opposition, parallel, or independent of selection by hawkmoths? Evolution 2013, 67, 2194–2206. [Google Scholar] [CrossRef]
- Blighe, K.; Lun, A. PCAtools: Everything principal Components Analysis. R Package Version 2.2.0. 2020. Available online: http://cran.r-project.org/web/packages/PCAtools/ (accessed on 1 May 2021).
- Hothorn, T.; Bretz, F.; Westfall, P.; Heiberger, R.M.; Schuetzenmeister, A.; Scheibe, S. Multcomp: Simultaneous Inference in General Parametric Models. R Package Version 1.3. 2014. Available online: http://cran.r-project.org/web/packages/multcomp/ (accessed on 2 May 2020).
- Garland, T.; Dickerman, A.W.; Janis, C.M.; Jones, J.A. Phylogenetic analysis of covariance by computer simulation. Syst. Biol. 1993, 42, 265–292. [Google Scholar] [CrossRef] [Green Version]
- Martins, E.P.; Hansen, T.F. Phylogenies and the comparative method: A general approach to incorporating phylogenetic information into the analysis of interspecific data. Am. Nat. 1997, 149, 646–667. [Google Scholar] [CrossRef] [Green Version]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Xiang, G.J.; Guo, Y.H.; Yang, C.F. Diversification of floral orientation in Lonicera is associated with pollinator shift and flowering phenology. J. Syst. Evol. 2021, 59, 557–566. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1. 2019. Available online: https://CRAN.R-project.org/package=nlme (accessed on 2 May 2020).
- Lefcheck, J.S. piecewiseSEM: Piecewise structural equation modelling in R for ecology, evolution, and systematics. Methods Ecol. Evol. 2016, 7, 573–579. [Google Scholar] [CrossRef]
- Wei, N.; Kaczorowski, R.L.; Arceo-Gómez, G.; O’Neill, E.M.; Hayes, R.A.; Ashman, T.L. Pollinators contribute to the maintenance of flowering plant diversity. Nature 2021, 597, 688–692. [Google Scholar] [CrossRef]
- Shipley, B. Confirmatory path analysis in a generalized multilevel context. Ecology 2009, 90, 363–368. [Google Scholar] [CrossRef]
- Shipley, B. The AIC model selection method applied to path analytic models compared using a d-separation test. Ecology 2013, 94, 560–564. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.r-project.org/ (accessed on 1 January 2019).




{kind=link}
{kind=link}
{kind=link}
| Species | Corolla Tube Length (mm, N = 30) | Throat Diameter (mm, N = 30) | Anther Height (mm, N = 30) | Stigma Height (mm, N = 30) | Upper Corolla Lip Length (mm, N = 30) | Upper Corolla Lip Width (mm, N = 30) | Lower Corolla Lip Length (mm, N = 30) | Lower Corolla Lip Width (mm, N = 30) | Phenotypic Integration Index (%) | PLa (mm) |
|---|---|---|---|---|---|---|---|---|---|---|
| L. chrysantha | 4.36 ± 0.53 h | 3.14 ± 0.044 bc | 7.75 ± 0.237 bc | 12.2 ± 0.115 f | 9.57 ± 0.097 d | 3.12 ± 0.043 cd | 9.73 ± 0.115 d | 3.05 ± 0.04 ef | 9.346 | 5.711 |
| L. elisae | 13.83 ± 0.15 c | 1.86 ± 0.046 g | 6.98 ± 0.217 c | 20.98 ± 0.267 c | 5.49 ± 0.091 f | 4.66 ± 0.073 b | 5.49 ± 0.091 f | 4.66 ± 0.073 a | 17.387 | 8.64 |
| L. ferdinandi | 8.8 ± 0.47 e | 2.94 ± 0.018 cd | 8.77 ± 0.220 b | 17.99 ± 0.278 d | 8.08 ± 0.069 e | 2.63 ± 0.030 e | 9.42 ± 0.082 d | 2.51 ± 0.028 g | 11.727 | 3.619 |
| L. gynochlamydea | 4.5 ± 0.29 gh | 2.32 ± 0.02 ef | 7.07 ± 0.312 c | 8.87 ± 0.157 g | 8.04 ± 0.027 e | 2.69 ± 0.117 de | 7.31 ± 0.032 e | 2.79 ± 0.11 fg | 25.164 | 4.457 |
| L. japonica | 25.12 ± 0.65 b | 3.51 ± 0.057 b | 22.12 ± 0.245 a | 47.18 ± 0.462 b | 22.21 ± 0.112 b | 4.27 ± 0.049 b | 24.02 ± 0.132 b | 3.91 ± 0.046 bc | 17.583 | 13.253 |
| L. maackii | 5.76 ± 0.48 fg | 2.59 ± 0.031 de | 7.98 ± 0.2284 bc | 14.76 ± 0.347 e | 14.41 ± 0.083 c | 3.44 ± 0.037 c | 13.24 ± 0.099 c | 3.49 ± 0.034 cd | 25.002 | 3.045 |
| L. pileata | 7.97 ± 0.25 e | 2.25 ± 0.033 eg | 5.44 ± 0.23 de | 10.45 ± 0.259 fg | 2.52 ± 0.131 g | 2.55 ± 0.0548 e | 2.52 ± 0.157 g | 2.55 ± 0.051 g | 12.597 | 5.392 |
| L. standishii | 6.52 ± 0.1 f | 2.1 ± 0.058 fg | 7.74 ± 0.299 bc | 10.93 ± 0.159 f | 7.41 ± 0.15 e | 2.57 ± 0.069 e | 7.71 ± 0.175 e | 2.66 ± 0.066 fg | 11.16 | 3.239 |
| L. tangutica | 10.5 ± 0.64 d | 4.46 ± 0.055 a | 4.51 ± 0.212 e | 16.8 ± 0.077 d | 3.29 ± 0.055 g | 3.26 ± 0.024 c | 3.29 ± 0.066 g | 3.26 ± 0.023 de | 12.792 | 7.817 |
| L. tragophylla | 45.59 ± 0.71 a | 4.34 ± 0.069 a | 21.64 ± 0.253 a | 73.76 ± 1.218 a | 25.66 ± 0.126 a | 5.48 ± 0.055 a | 28 ± 0.149 a | 5.1 ± 0.052 a | 33.724 | 41.15 |
| L. webbiana | 6.25 ± 0.31 f | 3.33 ± 0.073 bc | 6.84 ± 0.204 cd | 12.15 ± 0.18 f | 9.28 ± 0.041 d | 3.51 ± 0.179 c | 10.17 ± 0.049 d | 4.21 ± 0.168 b | 8.077 | 3.311 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, G.-J.; Lázaro, A.; Dai, X.-K.; Xia, J.; Yang, C.-F. Pollinator Proboscis Length Plays a Key Role in Floral Integration of Honeysuckle Flowers (Lonicera spp.). Plants 2023, 12, 1629. https://doi.org/10.3390/plants12081629
Xiang G-J, Lázaro A, Dai X-K, Xia J, Yang C-F. Pollinator Proboscis Length Plays a Key Role in Floral Integration of Honeysuckle Flowers (Lonicera spp.). Plants. 2023; 12(8):1629. https://doi.org/10.3390/plants12081629
Chicago/Turabian StyleXiang, Gan-Ju, Amparo Lázaro, Xiao-Kang Dai, Jing Xia, and Chun-Feng Yang. 2023. "Pollinator Proboscis Length Plays a Key Role in Floral Integration of Honeysuckle Flowers (Lonicera spp.)" Plants 12, no. 8: 1629. https://doi.org/10.3390/plants12081629