
The Effect of Glycine Betaine on Nitrogen and Polyamine Metabolisms, Expression of Glycoside-Related Biosynthetic Enzymes, and K/Na Balance of Stevia under Salt Stress


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparing Samples and Applying Treatments
2.2. Growth Parameters and Photosynthetic Pigments
2.3. Free Polyamines
2.4. Methionine and Arginine
2.5. Hydrogen Peroxide and Malondialdehyde
2.6. Relative Water Content and Membrane Stability Index
2.7. Activity of Polyamine Catabolizing Enzymes
2.8. Activity of Antioxidant Enzymes
2.9. Nitrogen and Nitrate Content
2.10. Activity of Nitrogen Metabolism Enzymes
2.11. Sodium and Potassium Concentration
2.12. Diterpene Glycosides
2.13. Gene Expression
2.14. Statistical Analyses
3. Results
3.1. Growth and Photosynthesis Traits
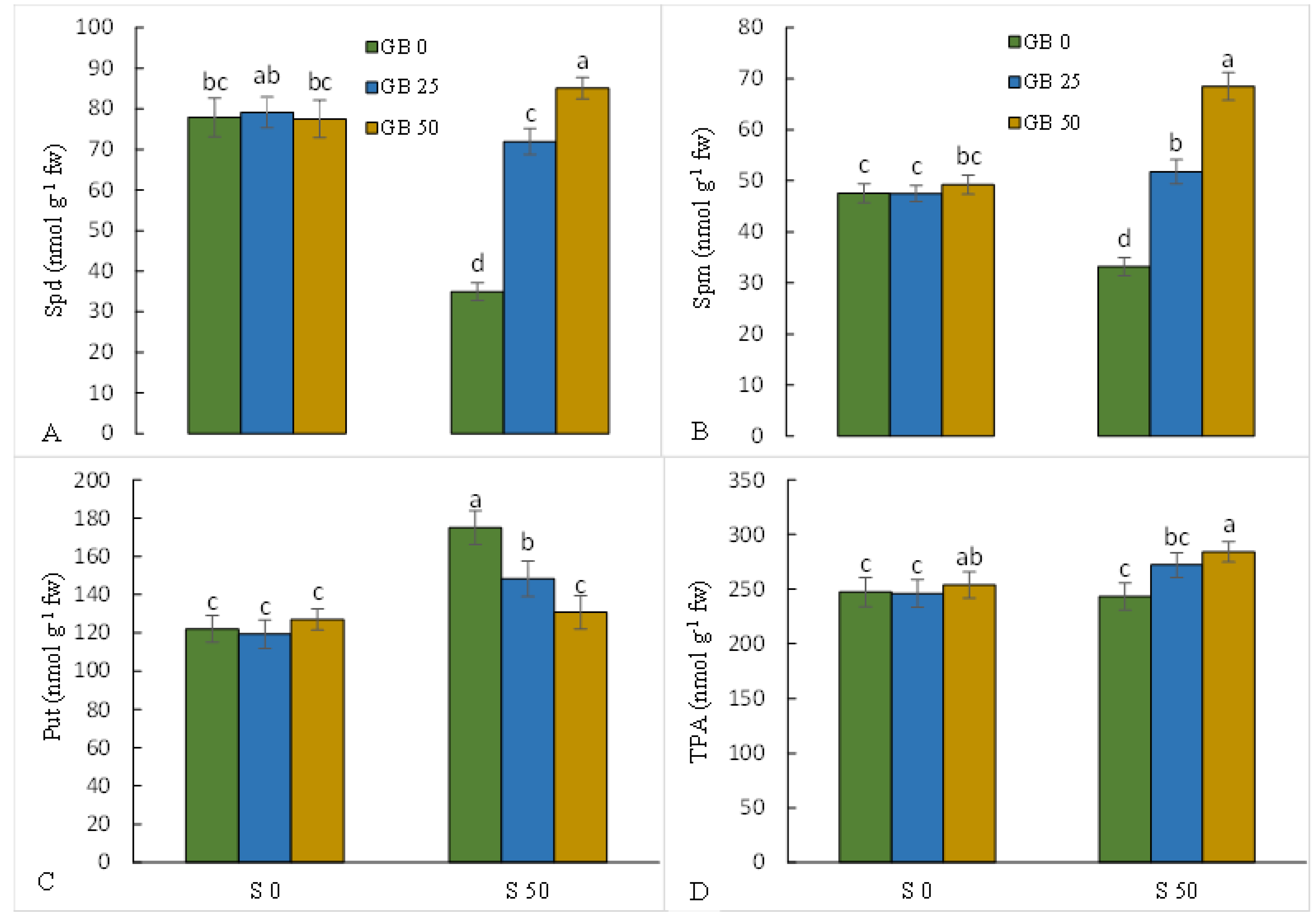
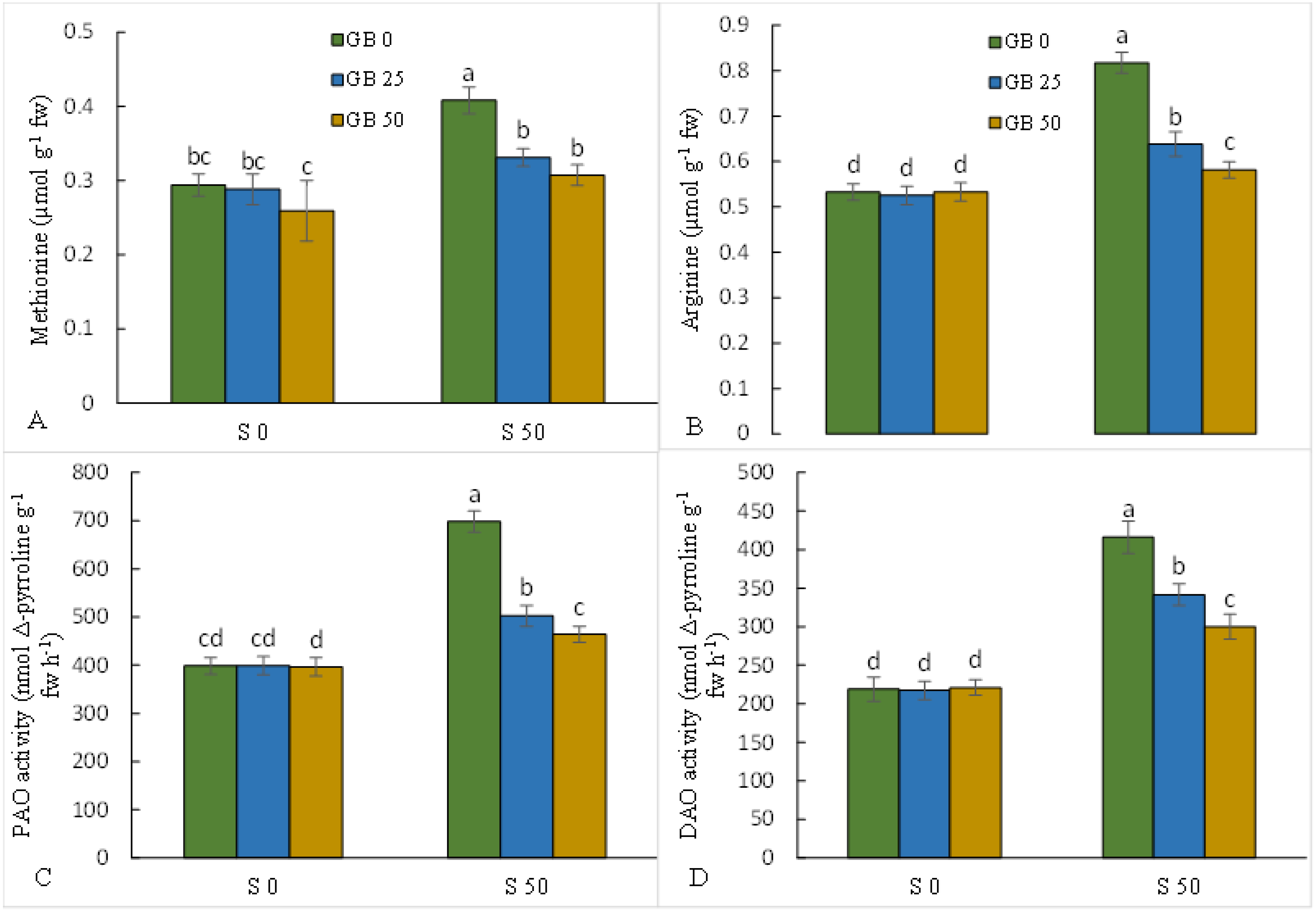
3.2. Metabolism of Polyamines
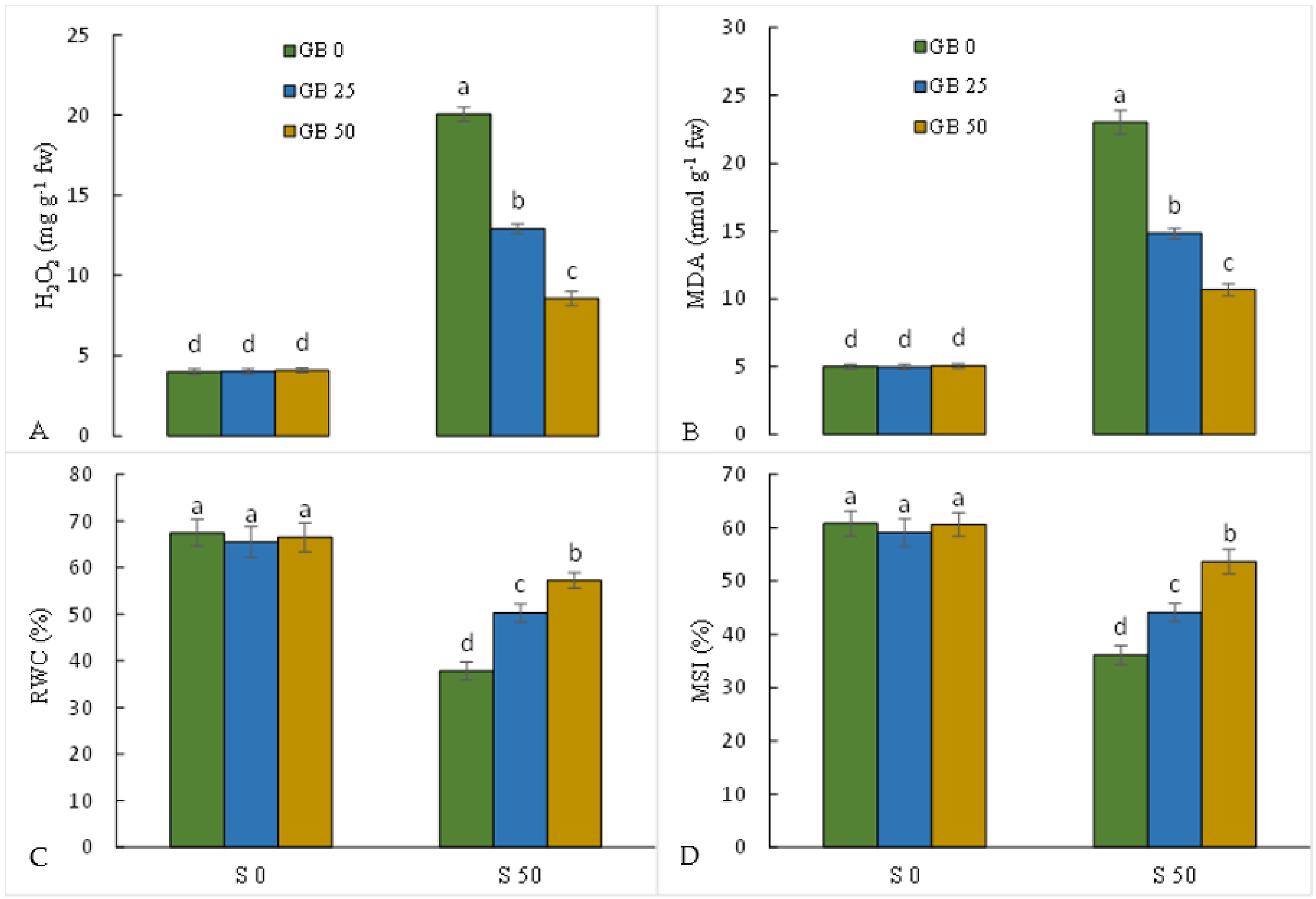
3.3. Oxidative Stress
3.4. K/Na Homeostasis
3.5. Nitrogen Metabolism
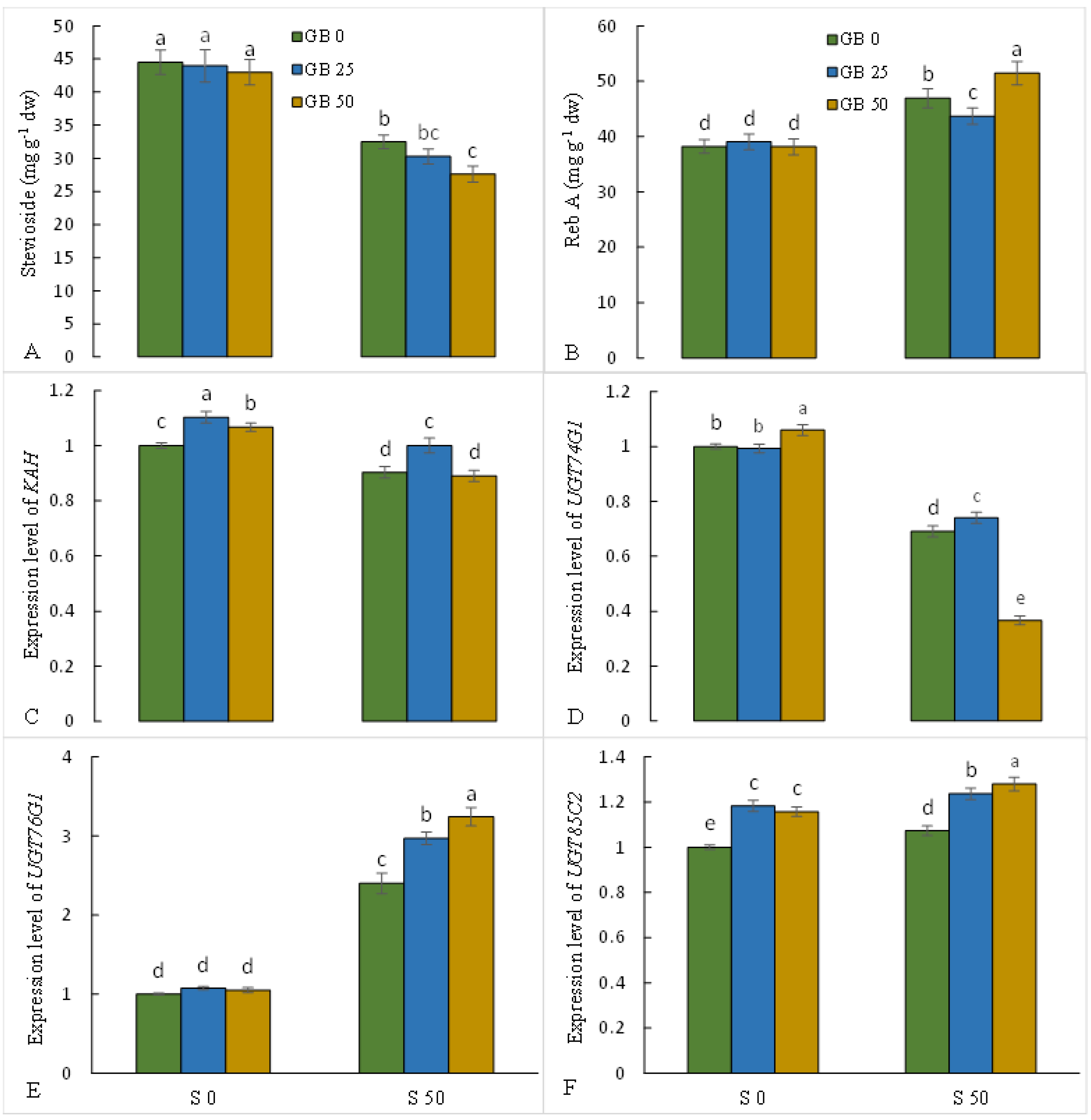
3.6. Diterpene Glycosides
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tester, M.; Langridge, P. Breeding technologies to increase crop production in a changing world. Science 2010, 327, 818–822. [Google Scholar] [CrossRef]
- Ghorbani, A.; Razavi, S.M.; Ghasemi Omran, V.; Pirdeshti, H. Effects of endophyte fungi symbiosis on some physiological parameters of tomato plants under 10 day long salinity stress. J. Plant Proc. Funct. 2019, 7, 193–208. [Google Scholar]
- Ke, Q.; Ye, J.; Wang, B.; Ren, J.; Yin, L.; Deng, X.; Wang, S. Melatonin mitigates salt stress in wheat seedlings by modulating polyamine metabolism. Front. Plant Sci. 2018, 9, 914. [Google Scholar] [CrossRef] [Green Version]
- Hao, S.; Wang, Y.; Yan, Y.; Liu, Y.; Wang, J.; Chen, S. A review on plant responses to salt stress and their mechanisms of salt resistance. Horticulturae 2021, 7, 132. [Google Scholar] [CrossRef]
- Ghorbani, A.; Pishkar, L.; Saravi, K.V.; Chen, M.X. Melatonin-mediated endogenous nitric oxide coordinately boosts stability through proline and nitrogen metabolism, antioxidant capacity, and Na+/K+ transporters in tomato under NaCl stress. Front. Plant Sci. 2023, 14, 1135943. [Google Scholar]
- Ghorbani, A.; Ghasemi Omran, V.O.; Razavi, S.M.; Pirdashti, H.; Ranjbar, M. Piriformospora indica confers salinity tolerance on tomato (Lycopersicon esculentum Mill.) through amelioration of nutrient accumulation, K+/Na+ homeostasis and water status. Plant Cell Rep. 2019, 38, 1151–1163. [Google Scholar] [CrossRef]
- Xu, Z.; Pehlivan, N.; Ghorbani, A.; Wu, C. Effects of Azorhizobium caulinodans and Piriformospora indica co-inoculation on growth and fruit quality of tomato (Solanum lycopersicum L.) under salt stress. Horticulturae 2022, 8, 302. [Google Scholar] [CrossRef]
- Ghorbani, A.; Tafteh, M.; Roudbari, N.; Pishkar, L.; Zhang, W.; Wu, C. Piriformospora indica augments arsenic tolerance in rice (Oryza sativa) by immobilizing arsenic in roots and improving iron translocation to shoots. Ecotoxicol. Environ. Saf. 2020, 209, 111793. [Google Scholar] [CrossRef]
- Ramezani, M.; Enayati, M.; Ramezani, M.; Ghorbani, A. A study of different strategical views into heavy metal (oid) removal in the environment. Arab. J. Geosci. 2021, 14, 2225. [Google Scholar] [CrossRef]
- Abd Elhamid, E.M.; Sadak, M.S.; Ezzo, M.I.; Abdalla, A.M. Impact of glycine betaine on drought tolerance of Moringa oleifera plant grown under sandy soil. Asian J. Plant Sci. 2021, 20, 578–589. [Google Scholar] [CrossRef]
- Bai, M.; Zeng, W.; Chen, F.; Ji, X.; Zhuang, Z.; Jin, B.; Wang, J.; Jia, L.; Peng, Y. Transcriptome expression profiles reveal response mechanisms to drought and drought-stress mitigation mechanisms by exogenous glycine betaine in maize. Biotechnol. Lett. 2022, 44, 367–386. [Google Scholar] [CrossRef]
- Zhu, M.; Li, Q.; Zhang, Y.; Zhang, M.; Li, Z. Glycine betaine increases salt tolerance in maize (Zea mays L.) by regulating Na+ homeostasis. Front. Plant Sci. 2022, 13, 978304. [Google Scholar] [CrossRef] [PubMed]
- Mäkelä, P.; Peltonen-Sainio, P.; Jokinen, K.; Pehu, E.; Setälä, H.; Hinkkanen, R.; Somersalo, S. Uptake and translocation of foliar-applied glycinebetaine in crop plants. Plant Sci. 1996, 121, 221–230. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Hamani, A.K.M.; Li, S.; Chen, J.; Amin, A.S.; Wang, G.; Xiaojun, S.; Zain, M.; Gao, Y. Linking exogenous foliar application of glycine betaine and stomatal characteristics with salinity stress tolerance in cotton (Gossypium hirsutum L.) seedlings. BMC Plant Biol. 2021, 21, 146. [Google Scholar] [CrossRef] [PubMed]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef] [Green Version]
- Quan, R.; Shang, M.; Zhang, H.; Zhao, Y.; Zhang, J. Engineering of enhanced glycine betaine synthesis improves drought tolerance in maize. Plant Biotechnol. J. 2004, 2, 477–486. [Google Scholar] [CrossRef]
- Annunziata, M.G.; Ciarmiello, L.F.; Woodrow, P.; Dell’Aversana, E.; Carillo, P. Spatial and temporal profile of glycine betaine accumulation in plants under abiotic stresses. Front. Plant Sci. 2019, 10, 230. [Google Scholar] [CrossRef] [Green Version]
- Carillo, P.; Mastrolonardo, G.; Nacca, F.; Parisi, D.; Verlotta, A.; Fuggi, A. Nitrogen metabolism in durum wheat under salinity: Accumulation of proline and glycine betaine. Funct. Plant Biol. 2008, 35, 412–426. [Google Scholar] [CrossRef]
- Gantait, S.; Banerjee, J. Geographical distribution, botanical description and self-incompatibility mechanism of genus Stevia. Sugar. Tech. 2017, 20, 1–10. [Google Scholar] [CrossRef]
- Ceunen, S.; Geuns, J.M.C. Steviol glycosides: Chemical diversity, metabolism, and function. J. Nat. Prod. 2013, 76, 1201–1228. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi-Omran, V.O.; Ghorbani, A.; Sajjadi-Otaghsara, S.A. Melatonin alleviates NaCl-induced damage by regulating ionic homeostasis, antioxidant system, redox homeostasis, and expression of steviol glycosides-related biosynthetic genes in in vitro cultured Stevia rebaudiana Bertoni. In Vitro Cell Dev. Biol. Plant 2021, 57, 319–331. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassay with tobacco tissues. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Naka, Y.; Watanabe, K.; Sagor, G.H.M.; Niitsu, M.; Pillai, M.A.; Kusano, T.; Takahashi, Y. Quantitative analysis of plant polyamines including thermospermine during growth and salinity stress. Plant Physiol. Biochem. 2010, 48, 527–533. [Google Scholar] [CrossRef]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Loreto, F.; Velikova, V. Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products, and reduces lipid peroxidation of cellular membranes. Plant Physiol. 2001, 127, 1781–1787. [Google Scholar] [CrossRef]
- Hodges, D.M.; De Long, J.M.; Forney, C.F.; Prange, R.K. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Turner, N.C.; Kramer, P.J. (Eds.) Adaptation of Plant to Water and High Temperature Stress; Wiley Interscience Pub.: New York, NY, USA, 1980; pp. 207–230. [Google Scholar]
- Asthir, B.; Duffus, C.M.; Smith, R.C.; Spoor, W. Diamine oxidase is involved in H2O2 production in the chalazal cells during barley grain filling. J. Exp. Bot. 2002, 53, 677–682. [Google Scholar] [CrossRef] [Green Version]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Cakmak, I.; Marschner, H. Magnesium-deficiency and high light-enhance activities of superoxide-dismutase, ascorbate peroxidase, and glutathione-reductase in bean-leaves. Plant Physiol. 1992, 98, 1222–1227. [Google Scholar] [CrossRef] [Green Version]
- Foyer, C.H.; Halliwell, B. The presence of glutathione and glutathione reductase in chloroplasts: A proposed role in ascorbic acid metabolism. Planta 1976, 133, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.J.; Leul, M. Uniconazole-induced tolerance of rape plants to heat stress in relation to changes in hormonal levels, enzyme activities and lipid peroxidation. Plant Growth Regul. 1999, 27, 99–104. [Google Scholar] [CrossRef]
- Singh, J.P. A rapid method for determination of nitrate in soil and plant extracts. Plant Soil 1988, 110, 137–139. [Google Scholar] [CrossRef]
- Debouba, M.; Gouia, H.; Valadier, M.H.; Ghorbel, M.H.; Suzuki, A. Salinity-induced tissue-specific diurnal changes in nitrogen assimilatory enzymes in tomato seedlings grown under high or low nitrate medium. Plant Physiol. Biochem. 2006, 44, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Agbaria, H.; Heuer, B.; Zieslin, N. Rootstock-imposed alterations in nitrate reductase and glutamine synthetase activities in leaves of rose plants. Biol. Plant. 1998, 41, 85–91. [Google Scholar] [CrossRef]
- Groat, R.G.; Vance, C.P. Root nodule enzymes of ammonia assimilation in alfalfa (Medicago sativa L.): Developmental patterns and response to applied nitrogen. Plant Physiol. 1981, 67, 1198–1203. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.T.; Wu, W.; Cai, Q.R.; Xu, Y.W.; Wei, C. Comparison on main agronomic traits and glucoside content in different Stevia rebaudiana new lines. Sugars China 2011, 3, 26–29. [Google Scholar]
- Ghorbani, A.; Zarinkamar, F.; Fallah, A. The effect of cold stress on the morphologic and physiologic characters of tow rice varieties in seedling stage. J. Crop. Breed. 2009, 1, 50–66. [Google Scholar]
- Lucho, S.R.; do Amaral, M.N.; Auler, P.A.; Bianchi, V.J.; Ferrer, M.Á.; Calderón, A.A.; Braga, E.J.B. Salt stress-induced changes in in vitro cultured Stevia rebaudiana bertoni: Effect on metabolite contents, antioxidant capacity and expression of steviol glycosides-related biosynthetic genes. J. Plant Growth Regul. 2019, 38, 1341–1353. [Google Scholar] [CrossRef]
- Shahverdi, M.A.; Omidi, H.; Tabatabaei, S.J. Stevia (Stevia rebaudiana Bertoni) responses to NaCl stress: Growth, photosynthetic pigments, diterpene glycosides and ion content in root and shoot. J. Saudi. Soc. Agric. Sci. 2019, 18, 335–360. [Google Scholar] [CrossRef]
- Debnath, M.; Ashwath, N.; Hill, C.B.; Callahane, D.L.; Dias, D.A.; Jayasinghe, D.S.; Midmore, D.J.; Roessner, U. Comparative metabolic and ionomic profiling of two cultivars of Stevia rebaudiana Bert. (Bertoni) grown under salinity stress. Plant Physiol. Biochem. 2018, 129, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, A.; Razavi, S.M.; Ghasemi Omran, V.O.; Pirdashti, H. Piriformospora indica inoculation alleviates the adverse effect of NaCl stress on growth, gas exchange and chlorophyll fluorescence in tomato (Solanum lycopersicum L.). Plant Biol. 2018, 20, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, A.; Razavi, S.M.; Ghasemi Omran, V.O.; Pirdashti, H. Piriformospora indica alleviates salinity by boosting redox poise and antioxidative potential of tomato. Russ. J. Plant Physiol. 2018, 65, 898–907. [Google Scholar]
- Kao, W.Y.; Tsai, T.T.; Shih, C.N. Photosynthetic gas exchange and chlorophyll a fluorescence of three wild soybean species in response to NaCl treatments. Photosynthetica 2003, 41, 415–419. [Google Scholar] [CrossRef]
- Raza, S.H.; Athar, H.R.; Ashraf, M.; Hameed, A. Glycinebetaine-induced modulation of antioxidant enzymes activities and ion accumulation in two wheat cultivars differing in salt tolerance. Environ. Exp. Bot. 2007, 60, 368–376. [Google Scholar] [CrossRef]
- Sakr, M.T.; El-Sarkassy, N.M.; Fuller, M.P. Osmoregulators proline and glycine betaine counteract salinity stress in canola. Agron. Sustain. Dev. 2012, 32, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Hasanuzzaman, M.; Nahar, K.; Fujita, M. Exogenous proline and glycine betaine mediated upregulation of antioxidant defense and glyoxalase systems provides better protection against salt-induced oxi- dative stress in two rice (Oryza sativa L.) varieties. BioMed Res. Int. 2014, 2014, 757219. [Google Scholar] [CrossRef] [Green Version]
- Sofy, M.R.; Elhawat, N.; Alshaal, T. Glycine betaine counters salinity stress by maintaining high K+/Na+ ratio and antioxidant defense via limiting Na+ uptake in common bean (Phaseolus vulgaris L.). Ecotoxicol. Environ. Saf. 2020, 200, 110732. [Google Scholar] [CrossRef]
- Shetty, K.; Shetty, G.A.; Nakazaki, Y.; Yoshioka, K.; Asano, Y.; Oosawa, K. Stimulation of benzyladenine induced in vitro shoot organogensis in Cucumus melo L. by proline, salicylic acid and aspirin. Plant Sci. 1992, 84, 193–199. [Google Scholar] [CrossRef]
- Mishra, S.N.; Sharma, I. Putrescine as a growth inducer and as a source of nitrogen for mustard seedlings under sodium chloride salinity. Indian J. Expt. Physiol. 1994, 32, 916–918. [Google Scholar]
- Hu, L.; Hu, T.; Zhang, X.; Pang, H.; Fu, J. Exogenous glycine betaine ameliorates the adverse effect of salt stress on perennial ryegrass. J. Amer. Soc. Hort. Sci. 2012, 137, 38–46. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, T.; Ashraf, M.; Shahbaz, M. Does exogenous application of glycine betaine as a pre-sowing seed treatment improve growth and regulate some key physiological attributes in wheat plants grown under water deficit conditions? Pakistan J. Bot. 2009, 41, 1291–1302. [Google Scholar]
- Wei, D.D.; Zhang, W.; Wang, C.C.; Mengm, Q.W.; Li, G.; Chen, T.H.H.; Yang, X.H. Genetic engineering of the biosynthesis of glycine betaine leads to alleviate salt-induced potassium efflux and enhances salt tolerance in tomato plants. Plant Sci. 2017, 257, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Abogadallah, G.M. Antioxidative defense under salt stress. Plant Signal. Behav. 2010, 5, 369–374. [Google Scholar] [CrossRef] [Green Version]
- Allakhverdiev, S.I.; Kreslavski, V.D.; Klimov, V.V.; Los, D.A.; Carpentier, R.; Mohanty, P. Heat stress: An overview of molecular responses in photosynthesis. Photosynth. Res. 2008, 98, 541–550. [Google Scholar] [CrossRef]
- Sakamoto, A.; Murata, N. The role of glycine betaine in the protection of plants from stress: Clues from transgenic plants. Plant Cell Environ. 2002, 25, 163–171. [Google Scholar] [CrossRef]
- Figueroa-Soto, C.G.; Valenzuela-Soto, E.M. Glycine betaine rather than acting only as an osmolyte also plays a role as regulator in cellular metabolism. Biochimie 2018, 147, 89–97. [Google Scholar] [CrossRef]
- Desoky, E.M.; Ibrahim, S.A.; Merwad, A.M. Mitigation of salinity stress effects on growth, physio-chemical parameters and yield of snapbean (Phaseolus vulgaris L.) by exogenous application of glycine betaine. Int. Lett. Nat. Sci. 2019, 76, 60–71. [Google Scholar] [CrossRef]
- Gerami, M.; Ghorbani, A.; Karimi, S. Role of salicylic acid pretreatment in alleviating cadmium-induced toxicity in Salvia officinalis L. Iran J. Plant Biol. 2018, 10, 81–95. [Google Scholar]
- Ghorbani, A.; Pishkar, L.; Roodbari, N.; Ali Tavakoli, S.; Moein Jahromi, E.; Chu, W. Nitrate reductase is needed for methyl jasmonate-mediated arsenic toxicity tolerance of rice by modulating the antioxidant defense system, glyoxalase system and arsenic sequestration mechanism. J. Plant Growth. Regul. 2022, 42, 1107. [Google Scholar] [CrossRef]
- Talaat, N.B. Polyamine and nitrogen metabolism regulation by melatonin and salicylic acid combined treatment as a repressor for salt toxicity in wheat (Triticum aestivum L.) plants. Plant Growth Regul. 2021, 95, 315–329. [Google Scholar] [CrossRef]
- Hussain, S.J.; Khan, N.A.; Anjum, N.A.; Masood, A.; Khan, M.I.R. Mechanistic elucidation of salicylic acid and sulphur-induced defence systems, nitrogen metabolism, photosynthetic, and growth potential of Mungbean (Vigna radiata) Under Salt Stress. J. Plant Growth Regul. 2021, 40, 1000–1016. [Google Scholar] [CrossRef]
- Ghorbani, A.; Pishkar, L.; Roodbari, N.; Pehlivan, N.; Wu, C. Nitric oxide could allay arsenic phytotoxicity in tomato (Solanum lycopersicum L.) by modulating photosynthetic pigments, phytochelatin metabolism, molecular redox status and arsenic sequestration. Plant Physiol. Biochem. 2021, 167, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Abbas, Z.; Seleiman, M.F.; Rizwan, M.; YavaŞ, İ.; Alhammad, B.A.; Shami, A.; Hasanuzzaman, M.; Kalderis, D. Glycine betaine accumulation, significance and interests for heavy metal tolerance in plants. Plants 2020, 9, 896. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, A.; Zarinkamar, F.; Fallah, A. Effect of cold stress on the anatomy and morphology of the tolerant and sensitive cultivars of rice during germination. J. Cell Tissue 2011, 2, 235–244. [Google Scholar]
- ElSayed, A.I.; Mohamed, A.H.; Rafudeen, M.S.; Omar, A.A.; Awad, M.F.; Mansour, E. Polyamines mitigate the destructive impacts of salinity stress by enhancing photosynthetic capacity, antioxidant defense system and upregulation of calvin cycle-related genes in rapeseed (Brassica napus L.). Saudi J. Biol. Sci. 2022, 29, 3675–3686. [Google Scholar] [CrossRef]
- Handa, A.K.; Fatima, T.; Mattoo, A.K. Polyamines: Bio-molecules with diverse functions in plant and human health and disease. Front. Chem. 2018, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Alcázar, R.; Bueno, M.; Tiburcio, A.F. Polyamines: Small Amines with Large Effects on Plant Abiotic Stress Tolerance. Cells 2020, 9, 2373. [Google Scholar] [CrossRef]
- Liu, J.; Liu, H.P.; Liu, Y.L. Effect of exogenous glycine betaine on polyamines and ions content of barley seedlings under salt stress. Acta Agron. Sin. 2004, 30, 1119–1123. [Google Scholar]
- Zeng, J.; Cheng, A.; Lim, D.; Yi, B.; Wu, W. Effects of salt stress on the growth, physiological responses, and glycoside contents of Stevia rebaudiana Bertoni. J. Agric. Food Chem. 2013, 61, 5720–5726. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments (mM) | N Node | N Root | TDW | Chl a | Chl b | Car |
|---|---|---|---|---|---|---|
| mg/gFW | ||||||
| Control | 8.10 ± 0.26 a | 7.55 ± 0.20 b | 0.251 ± 0.022 a | 1.91 ± 0.15 a | 1.10 ± 0.08 a | 0.348 ± 0.023 a |
| GB 25 | 8.39 ± 0.17 a | 8.08 ± 0.24 a | 0.275 ± 0.021 a | 2.03 ± 0.16 a | 1.08 ± 0.07 a | 0.354 ± 0.015 a |
| GB 50 | 8.24 ± 0.13 a | 7.45 ± 0.21 bc | 0.251 ± 0.016 a | 2.05 ± 0.10 a | 1.04 ± 0.10 ab | 0.342 ± 0.024 a |
| NaCl 50 | 3.66 ± 0.18 d | 4.88 ± 0.22 e | 0.121 ± 0.009 d | 1.02 ± 0.07 d | 0.64 ± 0.03 c | 0.207 ± 0.018 d |
| NaCl 50 + GB 25 | 5.29 ± 0.15 c | 6.73 ± 0.23 d | 0.179 ± 0.011 c | 1.47 ± 0.08 c | 0.92 ± 0.04 b | 0.263 ± 0.016 c |
| NaCl 50 + GB 50 | 6.24 ± 0.20 b | 7.10 ± 0.23 cd | 0.209 ± 0.009 b | 1.69 ± 0.09 b | 0.99 ± 0.04 ab | 0.299 ± 0.016 b |
| Treatments (mM) | K | Na | K/Na | N (mg/gDW) | NO3 (µmol/gfw) |
|---|---|---|---|---|---|
| mg/gDW | |||||
| Control | 4.81 ± 0.30 ab | 0.72 ± 0.04 d | 6.70 ± 0.16 a | 33.07 ± 0.60 a | 28.17 ± 0.72 a |
| GB 25 | 4.81 ± 0.30 ab | 0.73 ± 0.03 d | 6.56 ± 0.12 a | 32.26 ± 0.55 a | 26.96 ± 1.20 a |
| GB 50 | 4.90 ± 0.27 a | 0.72 ± 0.04 d | 6.82 ± 0.05 a | 32.44 ± 0.79 a | 27.39 ± 0.79 a |
| NaCl 50 | 2.99 ± 0.15 d | 2.46 ± 0.10 a | 1.22 ± 0.02 d | 17.87 ± 0.60 d | 13.70 ± 0.44 d |
| NaCl 50 + GB 25 | 3.96 ± 0.16 c | 1.30 ± 0.06 b | 3.06 ± 0.26 c | 25.33 ± 0.43 c | 20.91 ± 0.63 c |
| NaCl 50 + GB 50 | 4.39 ± 0.14 b | 1.02 ± 0.10 c | 4.36 ± 0.53 b | 28.25 ± 0.43 b | 22.96 ± 0.64 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghorbani, A.; Ghasemi-Omran, V.O.; Chen, M. The Effect of Glycine Betaine on Nitrogen and Polyamine Metabolisms, Expression of Glycoside-Related Biosynthetic Enzymes, and K/Na Balance of Stevia under Salt Stress. Plants 2023, 12, 1628. https://doi.org/10.3390/plants12081628
Ghorbani A, Ghasemi-Omran VO, Chen M. The Effect of Glycine Betaine on Nitrogen and Polyamine Metabolisms, Expression of Glycoside-Related Biosynthetic Enzymes, and K/Na Balance of Stevia under Salt Stress. Plants. 2023; 12(8):1628. https://doi.org/10.3390/plants12081628
Chicago/Turabian StyleGhorbani, Abazar, Vali Ollah Ghasemi-Omran, and Moxian Chen. 2023. "The Effect of Glycine Betaine on Nitrogen and Polyamine Metabolisms, Expression of Glycoside-Related Biosynthetic Enzymes, and K/Na Balance of Stevia under Salt Stress" Plants 12, no. 8: 1628. https://doi.org/10.3390/plants12081628