
Identification of Single-Nucleotide Polymorphisms (SNPs) Associated with Heat Tolerance at the Reproductive Stage in Synthetic Hexaploid Wheats Using GWAS
 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
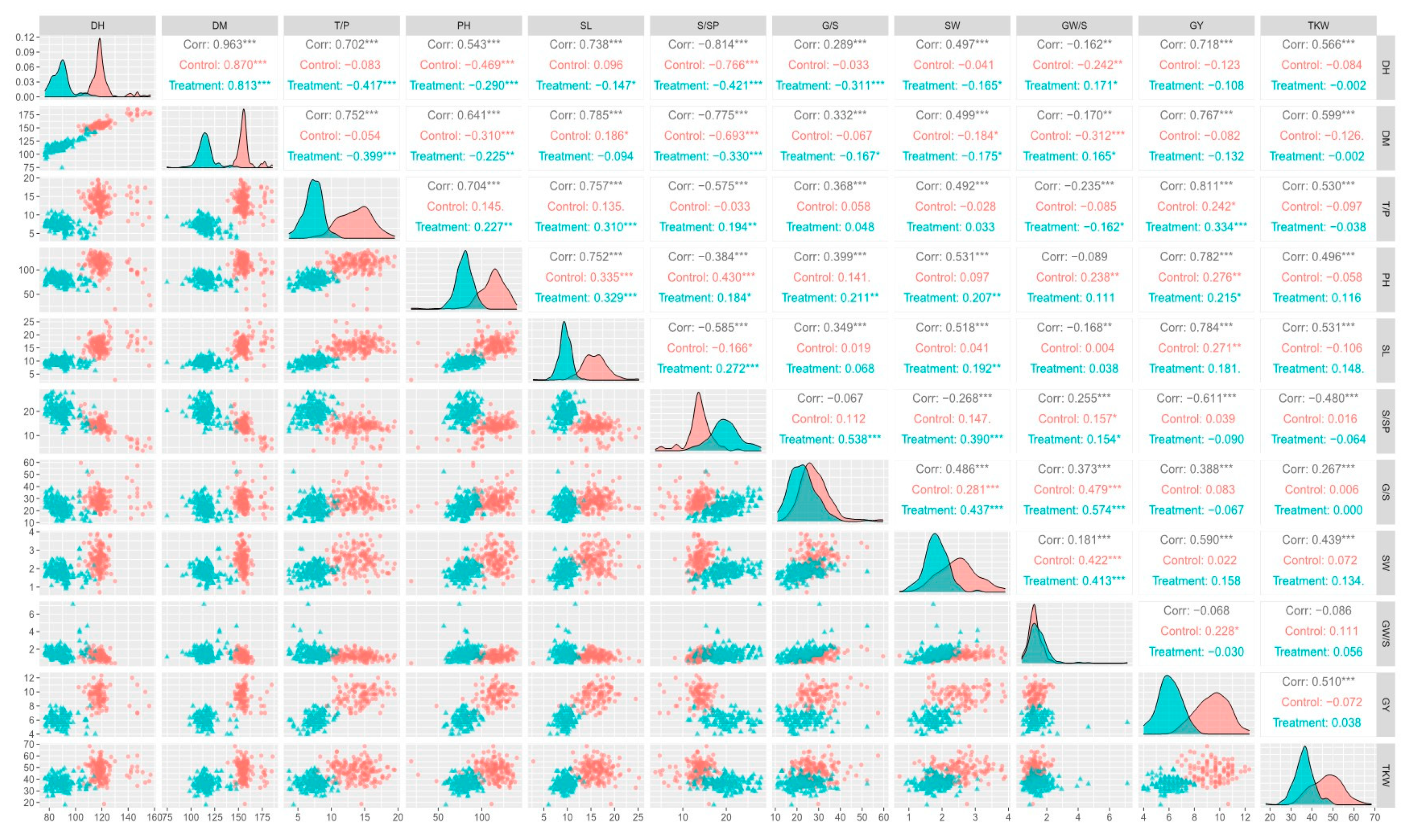
2.1. Phenotypic Data Analysis
2.2. Marker Statistics
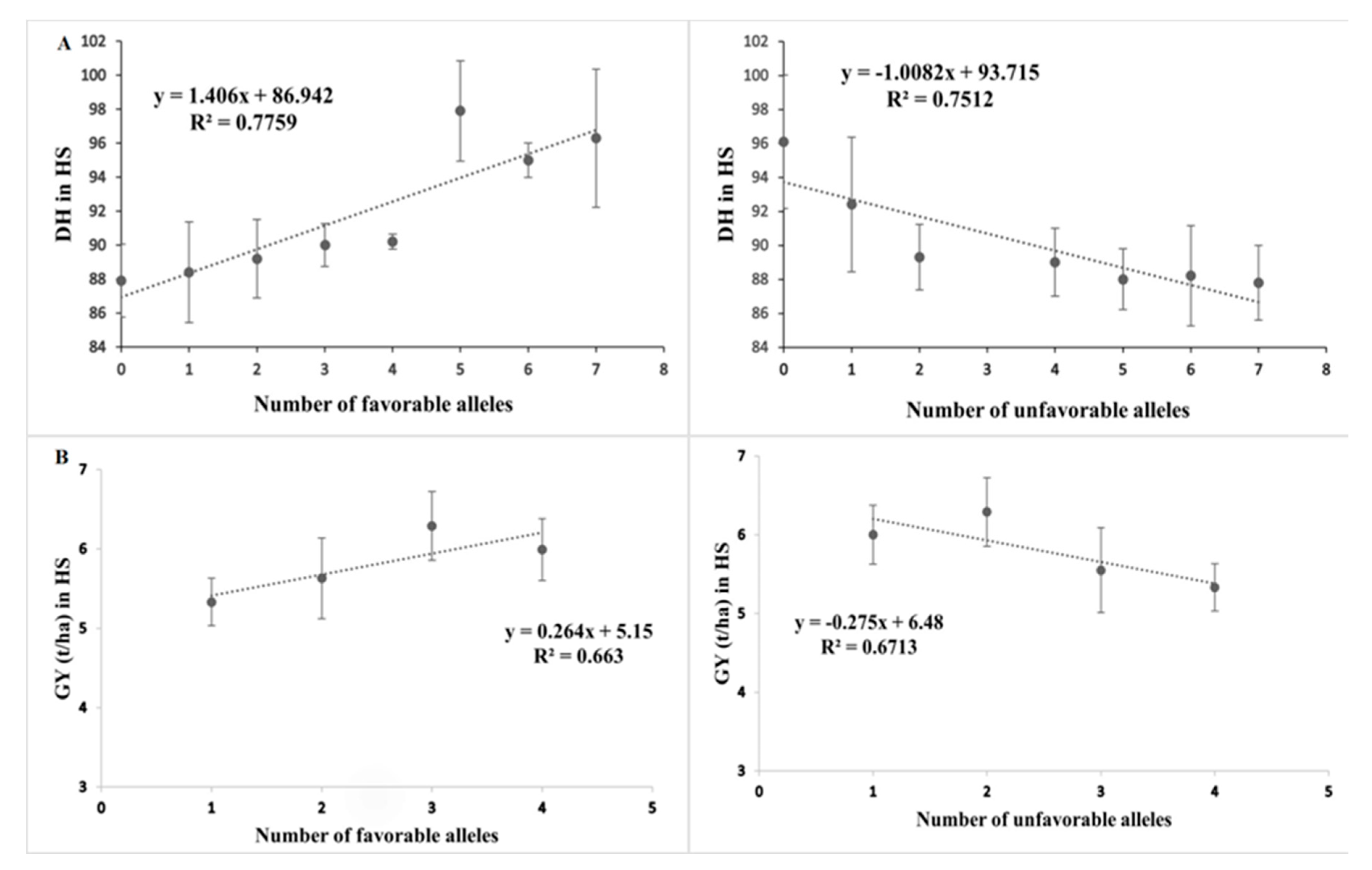
2.3. QTNs Associated with Yield Traits under Heat Stress
2.4. Co-Localization of QTNs and Heat Shock Proteins
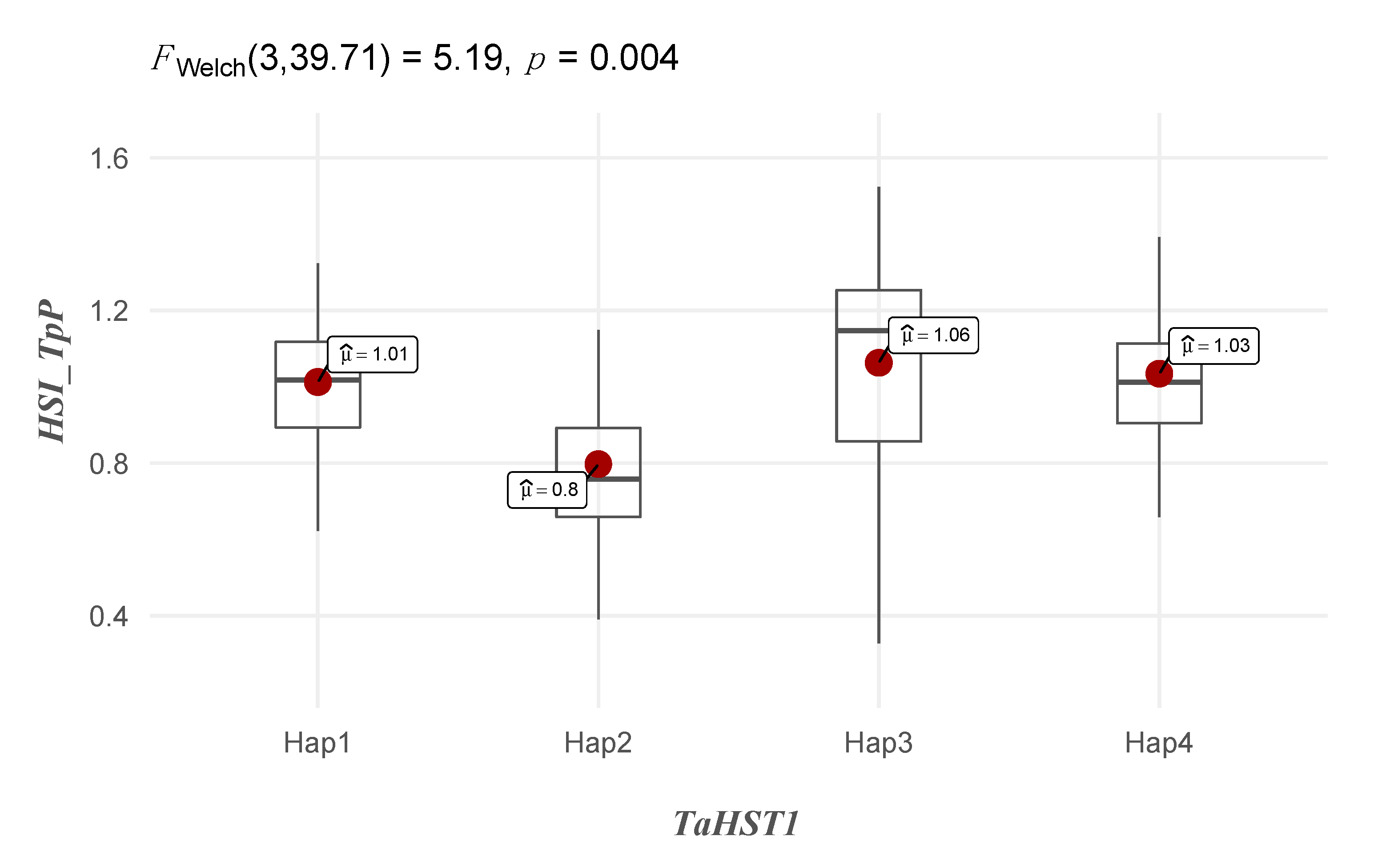
2.5. Characterization of the TaHST1 Locus in the Diversity Panel
3. Discussion
3.1. Phenotypic Variation for Agronomic Traits in SHWs under Heat Stress
3.2. Effects of Heat Stress in Synthetic Hexaploid Wheat
3.3. GWASs for the Identification of Loci Associated with Heat Stress Tolerance
3.4. Co-Localization of QTNs and HeatShock Proteins
3.5. Characterization of the TaHST1 Locus in the Diversity Panel
4. Materials and Methods
4.1. Plant Material
4.2. Field Evaluation
4.3. Genotyping for the TaHST1Locus
4.4. DNA Extraction and Genotyping
4.5. Statistical Analysis
4.5.1. Analysis of Phenotypic Data
4.5.2. Genotypic Data Analysis and GWASs
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Faostat, F. 2017. Available online: https://www.fao.org/faostat/en/#data (accessed on 18 January 2018).
- Thistlthwaite, R.J.; Tan, D.K.Y.; Bokshi, A.I.; Ullah, S.; Trethowan, R.M. A phenotyping strategy for evaluating the high temperature tolerance of wheat. Field Crops Res. 2020, 255, 107905. [Google Scholar] [CrossRef]
- Asseng, S.; Foster, I.; Turner, N.C. The impact of temperature variability on wheat yields. Glob. Chang. Biol. 2011, 17, 997–1012. [Google Scholar] [CrossRef]
- Cossani, C.M.; Reynolds, M.P. Physiological Traits for Improving Heat Tolerance in Wheat. Plant Physiol. 2012, 160, 1710–1718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Rasheed, A.; Hickey, L.; He, Z. Fast-Forwarding Genetic Gain. Trends Plant Sci. 2018, 23, 184–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Truong, H.A.; Lee, W.J.; Kishii, M.; Hong, S.-W.; Kang, C.-S.; Lee, B.C.; Lee, H. Assessment of synthetic hexaploid wheats in response to heat stress and leaf rust infection for the improvement of wheat production. Crop Pasture Sci. 2019, 70, 837–848. [Google Scholar] [CrossRef]
- Dreisigack, S.; Crossa, J.; Pérez-Rodríguez, P.; Montesinos-López, O.A.; Rosyara, U.; Juliana, P.; Mondal, S.; Crespo-Herrera, L.; Govindan, V.; Singh, R.P.; et al. Implementation of genomic selection in the CIMMYT global wheat program, findings from the past 10 years. Crop Breed. Genet. Genom. 2021, 3, e210005. [Google Scholar]
- Trethowan, R.M.; VanGinkel, M. Synthetic Wheat—An Emerging Genetic Resource; WheatSci. Trade. Ed.B. Carver; Wiley-Blackwell: Ames, IA, USA, 2009; pp. 369–386. [Google Scholar]
- International Wheat Genome Sequencing Consortium (IWGSC); Appels, R.; Eversole, K.; Stein, N.; Feuillet, C.; Keller, B.; Rogers, J.; Pozniak, C.J.; Choulet, F.; Distelfeld, A.; et al. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar]
- Zhang, R.; Liu, G.; Xu, H.; Lou, H.; Zhai, S.; Chen, A.; Hao, S.; Xing, J.; Liu, J.; You, M.; et al. Heat Stress Tolerance 2 confers basal heat stress tolerance in allohexaploid wheat (Triticum aestivum L.). J. Exp. Bot. 2022, 73, 6600–6614. [Google Scholar] [CrossRef]
- Su, P.; Jiang, C.; Qin, H.; Hu, R.; Feng, J.; Chang, J.; Yang, G.; He, G. Identification of potential genes responsiblefor thermo-tolerance in wheat under high temperature stress. Genes 2019, 10, 174. [Google Scholar] [CrossRef] [Green Version]
- Talukder, S.K.; Babar, A.; Vijayalakshmi, K.; Poland, J.; Prasad, P.V.V.; Bowden, R.; Fritz, A. Mapping QTL for the traits associated with heat tolerance in wheat (Triticum aestivum L.). BMC Genet. 2014, 15, 97. [Google Scholar] [CrossRef] [Green Version]
- Bennett, D.; Reynolds, M.; Mullan, D.; Izanloo, A.; Kuchel, H.; Langridge, P.; Schnurbusch, T. Detection of two major grain yield QTL in bread wheat (Triticum aestivum L.) unde rheat, drought and high yield potential environments. Theor. Appl. Genet. 2012, 125, 1473–1485. [Google Scholar] [CrossRef] [PubMed]
- Mason, R.E.; Hays, D.B.; Mondal, S.; Ibrahim, A.M.H.; Basnet, B.R. QTL for yield, yield components and canopy temperature depression in wheat under late sown field conditions. Euphytica 2013, 194, 243–259. [Google Scholar] [CrossRef]
- Maulana, F.; Ayalew, H.; Anderson, J.D.; Kumssa, T.T.; Huang, W.; Ma, X.-F. Genome-Wide Association Mapping of Seedling Heat Tolerance in Winter Wheat. Front. Plant Sci. 2018, 9, 1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, H.G.M.D.; Naeem, M.; Zeng, Y.; Rashid, M.A.R.; Ullah, A.; Saeed, A.; Qadeer, A. Genome-wide association mapping for high temperature tolerance in wheat through 90k SNP array using physiological and yield traits. PLoS ONE 2022, 17, e0262569. [Google Scholar] [CrossRef]
- Lopes, M.S.; Dreisigacker, S.; Peña, R.J.; Sukumaran, S.; Reynolds, M.P. Genetic characterization of the wheat association mapping initiative (WAMI) panel for dissection of complex traits in spring wheat. Theor. Appl. Genet. 2015, 128, 453–464. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, J.; Mao, W.; Guan, P.; Wang, Y.; Chen, Y.; Liu, W.; Guo, W.; Yao, Y.; Hu, Z.; et al. Association mapping identifies loci and candidate genes for grain-related traits in spring wheat in response to heat stress. Plant Sci. 2023, 331, 111676. [Google Scholar] [CrossRef]
- Ullah, S.; Randhawa, I.A.S.; Trethowan, R. Genome-wide association study of multiple traits linked to heat tolerance in emmer-derived hexaploid wheat genotypes. Mol. Breed. 2022, 41, 29. [Google Scholar] [CrossRef]
- Zhai, H.; Jiang, C.; Zhao, Y.; Yang, S.; Li, Y.; Yan, K.; Wu, S.; Luo, B.; Du, Y.; Jin, H.; et al. Wheat heat tolerance is impaired by heightened deletions in the distal end of 4AL chromosomal arm. Plant Biotechnol. J. 2021, 19, 1038–1051. [Google Scholar] [CrossRef]
- Tanin, M.J.; Saini, D.K.; Sandhu, K.S.; Pal, N.; Gudi, S.; Chaudhary, J.; Sharma, A. Consensus genomic regions associated with multiple abiotic stress tolerance in wheat and implications for wheat breeding. Sci. Rep. 2022, 12, 13680. [Google Scholar] [CrossRef]
- Aziz, A.; Mahmood, T.; Mahmood, Z.; Shazadi, K.; Mujeeb-Kazi, A.; Rasheed, A. Genotypic variation and Genotype × Environment Interaction for Yield-Related Traits in Synthetic Hexaploid Wheats under a Range of Optimal and Heat-Stressed Environments. Crop Sci. 2018, 58, 295–303. [Google Scholar] [CrossRef]
- Sharma, P.; Sareen, S.; Saini, M.; Verma, A.; Tyagi, B.S.; Sharma, I. Assessing genetic variation for heat tolerance in synthetic wheat lines using phenotypic data and molecular markers. Aust. J. Crop Sci. 2014, 8, 515–522. [Google Scholar]
- Sehgal, S.K.; Kaur, S.; Gupta, S.; Sharma, A.; Kaur, R.; Bains, N.S. A direct hybridization approach to gene transfer from Aegilops tauschii Coss. To Triticum aestivum L. Plant Breed. 2011, 130, 98–100. [Google Scholar] [CrossRef]
- Cossani, C.M.; Reynolds, M.P. Heat Stress Adaptation in Elite Lines Derived from Synthetic Hexaploid Wheat. Crops Sci. 2015, 55, 2719–2735. [Google Scholar] [CrossRef]
- Mohammadi, R.; Armion, M.; Sadeghzadeh, D.; Amri, A.; Nachit, M. Analysis of Genotype-by-Environment Interaction for Agronomic Traits of Durum Wheat in Iran. Plant Prod. Sci. 2011, 14, 15–21. [Google Scholar] [CrossRef]
- Degewione, A.; Alamerew, S. Genetic Diversity in Bread Wheat (Triticum aestivum L.) Genotypes. Pak. J. Biol. Sci. 2013, 16, 1330–1335. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and anti oxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 26, 217037. [Google Scholar] [CrossRef] [Green Version]
- Singh, J.P.; Shambhoo, P.; Singh, K.N.; Randhir, S. Screening of heat tolerant wheat varieties by membrane thermo stability index in relation to yield and yield attributing traits. Int. J. Plant Sci. Muzz. 2007, 2, 159–165. [Google Scholar]
- Rahman, M.A.; Chikushi, J.; Yoshida, S.; Karim, A. Growth and yield components of wheat genotypes exposed to high temperature stress under control environment. Bangladesh J. Agric. Res. 2009, 34, 360–372. [Google Scholar] [CrossRef] [Green Version]
- Kazan, K.; Lyons, R. The link between flowering time and stress tolerance. J. Exp. Bot. 2015, 67, 47–60. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Mondal, S.; Cheng, D.; Wang, C.; Liu, A.; Song, J.; Li, H.; Zhao, Z.; Liu, J. Evaluation of agronomic and physiological traits associated with high temperature stress tolerance in the winter wheat cultivars. Acta Physiol. Plant. 2015, 37, 90. [Google Scholar] [CrossRef]
- Wardlaw, I.; Wrigley, C. Heat Tolerance in Temperate Cereals: An Overview. Funct. Plant Biol. 1994, 21, 695–703. [Google Scholar] [CrossRef]
- Reynolds, M.; Balota, M.; Delgado, M.; Amani, I.; Fischer, R. Physiological and Morphological Traits Associated with Spring Wheat Yield Under Hot, Irrigated Conditions. Funct. Plant Biol. 1994, 21, 717–730. [Google Scholar] [CrossRef]
- Tewolde, H.; Fernandez, C.J.; Erickson, C.A. Wheat Cultivars Adapted to Post-Heading High Temperature Stress. J. Agron. Crop. Sci. 2006, 192, 111–120. [Google Scholar] [CrossRef]
- Asseng, S.; Ewert, F.; Martre, P.; Rotter, R.P.; Lobell, D.B.; Cammarano, D.; Kimball, B.A.; Ottman, M.J.; Wall, G.W.; White, J.W.; et al. Rising temperatures reduce global wheat production. Nat. Clim. Chang. 2015, 5, 143–147. [Google Scholar] [CrossRef]
- Vijayalakshmi, K.; Fritz, A.K.; Paulsen, G.M.; Bai, G.; Pandravada, S.; Gill, B.S. Modeling and mapping QTL for senescence related traits in winter wheat under high temperature. Mol. Breed. 2010, 26, 163–175. [Google Scholar] [CrossRef]
- Sangwan, S.; Munjal, R.; Ram, K.; Kumar, N. QTL mapping for morphological and physiological traits in RILs of spring wheat population of WH1021 X WH711. J. Environ. Biol. 2019, 40, 674–682. [Google Scholar] [CrossRef]
- Telfer, P.; Edwards, J.; Norman, A.; Bennett, D.; Smith, A.; Able, J.A.; Kuchel, H. Genetic analysis of wheat (Triticum aestivum) adaptation to heat stress. Theor. Appl. Genet. 2021, 134, 1387–1407. [Google Scholar] [CrossRef]
- Li, L.; Mao, X.; Wang, J.; Chang, X.; Reynolds, M.P.; Jing, R. Genetic dissection of drought and heat responsive agronomic traits in wheat. Plant Cell Environ. 2019, 42, 2540–2553. [Google Scholar] [CrossRef] [Green Version]
- Hassan, F.S.C.; Solouki, M.; Fakheri, B.A.; Nezhad, N.M.; Masoudi, B. Mapping QTLs for physiological and biochemical traits related to grain yield under control and terminal heat stress conditions in bread wheat (Triticum aestivum L.). Physiol. Mol. Biol. Plants 2018, 24, 1231–1243. [Google Scholar] [CrossRef]
- Ogbonnaya, F.C.; Rasheed, A.; Okechukwu, E.C.; Jighly, A.; Makdis, F.; Wuletaw, T.; Hagras, A.; Uguru, M.I.; Agbo, C.U. Genome-wide association study for agronomic and physiological traits in spring wheat evaluated in a range of heat prone environments. Theor. Appl. Genet. 2017, 130, 1819–1835. [Google Scholar] [CrossRef]
- Khan, M.I.; Kainat, Z.; Maqbool, S.; Mehwish, A.; Ahmad, S.; Suleman, H.M.; Mahmood, Z.; Ali, M.; Aziz, A.; Rasheed, A.; et al. Genome-wide association for heat tolerance at seedling stage in historical spring wheat cultivars. Front. Plant Sci. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, A.; Xia, X.; Ogbonnaya, F.; Mahmood, T.; Zhang, Z.; Mujeeb-Kazi, A.; He, Z. Genome-wide association for grain morphology in synthetic hexaploid wheats using digital imaging analysis. BMC Plant Biol. 2014, 14, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jighly, A.; Alagu, M.; Makdis, F.; Singh, M.; Singh, S.; Emebiri, L.C.; Ogbonnaya, F.C. Genomic regions conferring resistance to multiple fungal pathogens in synthetic hexaploid wheat. Mol. Breed. 2016, 36, 127. [Google Scholar] [CrossRef]
- Zegeye, H.; Rasheed, A.; Makdis, F.; Badebo, A.; Ogbonnaya, F.C. Genome-Wide Association Mapping for Seedling and Adult Plant Resistance to Stripe Rust in Synthetic Hexaploid Wheat. PLoS ONE 2014, 9, e105593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emebiri, L.C.; Oliver, J.R.; Mrva, K.; Mares, D. Association mapping of late maturity α-amylase (LMA) activity in a collection of synthetic hexaploid wheat. Mol. Breed. 2010, 26, 39–49. [Google Scholar] [CrossRef]
- Acevedo, E.; Silva, P.; Silva, H. Wheat growth and physiology. Bread Wheat Improv. Prod. 2002, 30, 39–70. [Google Scholar]
- Dreisigacker, S.; Tiwari, R.; Sheoran, S. Laboratory Manual: ICAR-CIMMYT Molecular Breeding Course in Wheat; ICAR/BMZ/CIMMYT: Haryana, India, 2013; p. 36. [Google Scholar]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative usage of fixed and random effect models for powerful and efficient genome-wide association studies. PLoS Genet. 2016, 12, e1005767. [Google Scholar] [CrossRef]
- Nyquist, W.E.; Baker, R. Estimation of heritability and prediction of selection response in plant populations. Crit. Rev. Plant Sci. 1991, 10, 235–322. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Control | HS a | HIS b | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Mean | Range | SD | Mean | Range | SD | Mean | Range | SD | |
| DH | 119.09 | 102–157 | 12.7 | 89.3 | 77–115 | 7.42 | 1 | 0.31–1.4 | 0.1 |
| DM | 156.5 | 139–186 | 7.7 | 115.5 | 75–148 | 9.11 | 1 | 0.55–1.97 | 0.1 |
| TpP | 13.67 | 7.3–20 | 2.4 | 7.28 | 4–11 | 1.4 | 0.97 | −1.81 | 0.3 |
| PH (cm) | 111.8 | 19–140 | 16.9 | 79.9 | 56–103 | 8.9 | 0.89 | −8–1.8 | 0.9 |
| SL (cm) | 15.8 | 3–25 | 2.8 | 9.6 | 6–14 | 1.2 | 0.93 | −5.7–1.8 | 0.6 |
| SpSP | 13.7 | 4–23 | 3.1 | 19.7 | 11–28 | 3.6 | 1.15 | −2.2–9.2 | 1.1 |
| GpS | 28.3 | 14–60 | 7.4 | 22.8 | 11–52 | 6.4 | 0.87 | −8–5 | 1.3 |
| SW (g) | 2.1 | 0.7–3.9 | 0.96 | 1.8 | 0.9–3 | 0.3 | 1.69 | −11–4.8 | 2.0 |
| GWpS (g) | 1.2 | 0.4–2.4 | 0.4 | 1.4 | 0.4–7 | 0.7 | 0.83 | −2–10 | 2.2 |
| GY (t/ha) | 9.4 | 7–12 | 1.2 | 6.03 | 4–8 | 0.9 | 0.59 | 0.4–0.8 | 0.09 |
| TKW (g) | 46.9 | 19–68 | 8.2 | 36.3 | 18–49 | 4.7 | 0.94 | −2.9–3.2 | 0.7 |
| Traits | Condition | SNP | CHR | POS | Pos(Mb) | MAF | Effect | p-Value | Previous QTL/QTN |
|---|---|---|---|---|---|---|---|---|---|
| DH | HS | AX-109842766 | 1A | 4.78 × 108 | 477.74 | 0.20 | 3.16 | 7.23 × 10−5 | |
| HS | AX-111599726 | 3D | 6.02 × 108 | 602.27 | 0.19 | 3.67 | 2.36 × 10−5 | ||
| HSI | AX-179559172 | 4B | 6.50 × 108 | 649.6 | 0.17 | 0.09 | 8.18 × 10−5 | ||
| HS | AX-94503623 | 5B | 6.19 × 108 | 619.14 | 0.24 | 3.46 | 5.61 × 10−5 | ||
| HS | AX-86180088 | 6A | 2.71 × 108 | 270.61 | 0.16 | 3.50 | 7.96 × 10−5 | ||
| HS | AX-95660535 | 6A | 3.15 × 108 | 315.41 | 0.17 | 3.60 | 4.87 × 10−5 | ||
| HS | AX-94744160 | 6A | 3.81 × 108 | 381.03 | 0.16 | 3.50 | 7.96 × 10−5 | ||
| HS | AX-109950287 | 6A | 3.90 × 108 | 390.08 | 0.16 | 3.70 | 3.85 × 10−5 | ||
| Control | AX-94444292 | 6B | 9.78 × 107 | 97.82 | 0.05 | −11.18 | 6.24 × 10−5 | ||
| HSI | AX-89339930 | 6B | 4.62 × 108 | 462.15 | 0.20 | 0.08 | 1.50 × 10−5 | ||
| HSI | AX-95169568 | 6B | 7.04 × 108 | 703.94 | 0.24 | 0.08 | 2.91 × 10−5 | ||
| HSI | AX-86173990 | 6D | 4.58 × 108 | 458.36 | 0.19 | 0.08 | 5.48 × 10−5 | ||
| HSI | AX-110389623 | 6D | 4.63 × 108 | 462.7 | 0.25 | 0.08 | 9.35 × 10−5 | ||
| Control | AX-95659681 | 7A | 7.21 × 108 | 721.22 | 0.39 | 7.79 | 8.04 × 10−5 | ||
| HS | AX-94980563 | 7B | 4.38 × 108 | 437.93 | 0.13 | 4.08 | 1.97 × 10−5 | ||
| HSI | AX-94750097 | 7D | 6.24 × 108 | 623.54 | 0.23 | 0.10 | 7.49 × 10−5 | ||
| DM | HSI | AX-94754683 | 2B | 6.71 × 106 | 6.71 | 0.05 | 0.13 | 9.39 × 10−5 | |
| Control | AX-94584932 | 2B | 3.22 × 108 | 321.96 | 0.31 | 2.55 | 4.77 × 10−4 | ||
| Control | AX-95660688 | 3B | 1.59 × 106 | 1.59 | 0.37 | 2.37 | 8.98 × 10−4 | ||
| Control | AX-109430132 | 3B | 6.29 × 108 | 628.8 | 0.14 | 3.16 | 8.93 × 10−4 | ||
| HS | AX-94786898 | 3D | 5.88 × 108 | 588.33 | 0.07 | −6.77 | 5.83 × 10−5 | ||
| Control | AX-94457757 | 4B | 6.53 × 108 | 652.98 | 0.10 | 5.59 | 5.61 × 10−4 | ||
| Control | AX-110458863 | 4D | 4.28 × 107 | 42.81 | 0.11 | 3.63 | 6.95 × 10−4 | ||
| Control | AX-94605765 | 6A | 8.50 × 107 | 84.98 | 0.31 | 2.67 | 2.94 × 10−4 | ||
| Control | AX-86180088 | 6A | 2.71 × 108 | 270.61 | 0.16 | 2.96 | 6.43 × 10−4 | ||
| HS | AX-110984345 | 6A | 3.85 × 108 | 384.73 | 0.15 | 4.75 | 6.89 × 10−5 | ||
| Control | AX-109950287 | 6A | 3.90 × 108 | 390.08 | 0.16 | 3.11 | 4.03 × 10−4 | ||
| HS | AX-109950287 | 6A | 3.90 × 108 | 390.08 | 0.16 | 4.59 | 8.56 × 10−5 | ||
| Control | AX-109886100 | 6B | 1.41 × 108 | 140.62 | 0.32 | 2.48 | 5.44 × 10−4 | ||
| Control | AX-109397401 | 6D | 8.03 × 105 | 0.8 | 0.05 | 5.04 | 6.60 × 10−4 | ||
| Control | AX-94980563 | 7B | 4.38 × 108 | 437.93 | 0.13 | 3.27 | 4.85 × 10−4 | ||
| HS | AX-94980563 | 7B | 4.38 × 108 | 437.93 | 0.13 | 5.25 | 2.36 × 10−5 | ||
| Control | AX-95658771 | 7B | 6.09 × 108 | 608.72 | 0.27 | 3.48 | 4.71 × 10−4 | ||
| Control | AX-94479139 | 7D | 5.07 × 108 | 507.25 | 0.15 | 5.26 | 3.18 × 10−4 | ||
| GpS | HS | AX-95150128 | 1D | 7.91 × 107 | 79.14 | 0.11 | −3.19 | 7.32 × 10−4 | |
| HS | AX-110335177 | 1D | 3.95 × 108 | 394.73 | 0.09 | 3.49 | 6.74 × 10−4 | ||
| HS | AX-110911350 | 2A | 4.42 × 107 | 44.18 | 0.12 | 2.85 | 7.46 × 10−4 | ||
| HS | AX-110046675 | 2A | 4.47 × 107 | 44.75 | 0.13 | 2.85 | 7.46 × 10−4 | ||
| HS | AX-110533769 | 2B | 7.17 × 108 | 717.22 | 0.21 | 2.50 | 6.62 × 10−4 | ||
| HS | AX-179562568 | 2D | 5.99 × 108 | 598.71 | 0.10 | 2.97 | 9.43 × 10−4 | ||
| HS | AX-94942177 | 3A | 4.75 × 107 | 47.55 | 0.43 | 2.77 | 2.66 × 10−4 | ||
| Control | AX-95244621 | 3A | 7.32 × 108 | 732.24 | 0.41 | 2.45 | 6.90 × 10−4 | ||
| HS | AX-109270732 | 3B | 6.37 × 108 | 637.08 | 0.09 | 4.10 | 1.04 × 10−4 | ||
| Control | AX-89477157 | 3B | 8.23 × 108 | 822.97 | 0.33 | 2.83 | 5.42 × 10−4 | ||
| Control | AX-94710887 | 3D | 6.14 × 108 | 613.74 | 0.34 | 2.83 | 5.42 × 10−4 | ||
| Control | AX-109290547 | 4A | 7.13 × 108 | 712.92 | 0.13 | 3.23 | 7.51 × 10−4 | ||
| Control | AX-110396807 | 4B | 4.49 × 107 | 44.94 | 0.47 | −2.40 | 4.71 × 10−4 | ||
| Control | AX-95094806 | 4D | 3.12 × 108 | 311.96 | 0.30 | 2.88 | 6.76 × 10−4 | ||
| Control | AX-95253860 | 4D | 5.00 × 108 | 499.62 | 0.38 | 2.25 | 9.17 × 10−4 | ||
| HSI | AX-94948850 | 5B | 4.26 × 108 | 426.09 | 0.05 | −1.44 | 7.86 × 10−5 | ||
| HS | AX-110559492 | 5B | 5.91 × 108 | 591.14 | 0.46 | 2.07 | 6.79 × 10−4 | MQTL5B.2 | |
| HSI | AX-110136588 | 5B | 6.50 × 108 | 650.08 | 0.11 | 0.65 | 9.19 × 10−5 | ||
| Control | AX-111160156 | 6A | 1.66 × 107 | 16.57 | 0.22 | 2.72 | 5.36 × 10−4 | ||
| Control | AX-179558749 | 6A | 1.01 × 108 | 101.3 | 0.10 | −3.69 | 6.05 × 10−4 | ||
| Control | AX-94729591 | 6B | 2.82 × 107 | 28.16 | 0.24 | 2.61 | 9.31 × 10−4 | ||
| Control | AX-112289907 | 6B | 4.11 × 108 | 411.1 | 0.38 | 2.72 | 4.17 × 10−4 | ||
| HS | AX-95653657 | 7B | 5.96 × 107 | 59.61 | 0.18 | −3.00 | 7.08 × 10−4 | ||
| Control | AX-94624899 | 7B | 7.09 × 108 | 709.38 | 0.37 | −2.53 | 4.15 × 10−4 | ||
| GWpS | HSI | AX-110932792 | 1A | 5.66 × 108 | 566.01 | 0.12 | 1.40 | 1.81 × 10−5 | |
| HSI | AX-95654468 | 1B | 1.43 × 108 | 142.52 | 0.06 | 2.70 | 1.43 × 10−5 | ||
| HS | AX-95633859 | 1D | 3.57 × 108 | 357.46 | 0.07 | 0.51 | 8.39 × 10−5 | ||
| Control | AX-94725049 | 2A | 3.45 × 106 | 3.45 | 0.08 | −0.27 | 3.26 × 10−5 | ||
| Control | AX-109388159 | 2B | 6.18 × 106 | 6.18 | 0.16 | −0.20 | 8.21 × 10−5 | ||
| HS | AX-94648480 | 2B | 1.88 × 108 | 188.02 | 0.10 | 0.48 | 7.50 × 10−5 | ||
| HS | AX-94997695 | 2B | 1.98 × 108 | 198.07 | 0.10 | 0.48 | 7.50 × 10−5 | ||
| HS | AX-95202875 | 3A | 5.72 × 108 | 571.79 | 0.06 | 0.66 | 3.93 × 10−5 | ||
| HSI | AX-108804725 | 3A | 7.13 × 108 | 713.21 | 0.09 | 1.49 | 9.30 × 10−5 | ||
| HS | AX-109911103 | 5A | 6.12 × 108 | 611.65 | 0.08 | 0.51 | 6.49 × 10−5 | ||
| HS | AX-94886530 | 5B | 1.87 × 108 | 186.84 | 0.05 | 0.61 | 6.92 × 10−5 | ||
| HS | AX-111441228 | 5D | 5.49 × 108 | 548.64 | 0.08 | 0.48 | 8.07 × 10−5 | ||
| HS | AX-94846296 | 6A | 7.40 × 107 | 74.03 | 0.05 | 0.58 | 8.93 × 10−5 | ||
| HS | AX-94746774 | 6A | 9.64 × 107 | 96.44 | 0.05 | 0.58 | 8.93 × 10−5 | ||
| HS | AX-110087041 | 6B | 1.32 × 108 | 132.12 | 0.05 | 0.58 | 8.93 × 10−5 | ||
| HS | AX-95685124 | 6B | 1.52 × 108 | 152.09 | 0.05 | 0.58 | 8.93 × 10−5 | ||
| HS | AX-179476704 | 6D | 6.21 × 107 | 62.11 | 0.08 | 0.51 | 6.93 × 10−5 | ||
| GY | HS | AX-94580758 | 1B | 6.27 × 108 | 626.6 | 0.50 | 0.34 | 3.32 × 10−4 | MQTL1B.4 |
| HS | AX-111214234 | 1D | 4.56 × 108 | 455.8 | 0.29 | −0.39 | 2.25 × 10−4 | MQTL1D.5 | |
| HSI | AX-94869969 | 2B | 3.87 × 108 | 387.13 | 0.21 | 0.06 | 6.36 × 10−5 | ||
| HSI | AX-95230131 | 2B | 6.38 × 108 | 638.26 | 0.19 | 0.05 | 8.41 × 10−5 | MQTL2B.3, MQTL2B.6, MQTL2B.7 | |
| HSI | AX-94411868 | 2D | 5.38 × 108 | 537.51 | 0.23 | 0.05 | 4.16 × 10−5 | ||
| HS | AX-111599726 | 3D | 6.02 × 108 | 602.27 | 0.19 | −0.45 | 3.26 × 10−4 | ||
| Control | AX-94752113 | 4A | 1.38 × 107 | 13.76 | 0.38 | 0.60 | 9.72 × 10−5 | ||
| Control | AX-94525682 | 4B | 5.64 × 108 | 564.22 | 0.39 | 0.59 | 8.24 × 10−5 | ||
| HSI | AX-108840130 | 4B | 6.22 × 108 | 621.71 | 0.15 | 0.05 | 3.17 × 10−5 | ||
| Control | AX-94988232 | 4D | 4.56 × 108 | 455.75 | 0.40 | 0.59 | 8.24 × 10−5 | ||
| HSI | AX-94882874 | 4D | 4.87 × 108 | 487.05 | 0.17 | 0.05 | 4.94 × 10−5 | ||
| HSI | AX-94423826 | 5B | 5.89 × 108 | 588.83 | 0.10 | −0.06 | 2.89 × 10−5 | ||
| HSI | AX-111052660 | 5B | 6.11 × 108 | 610.88 | 0.30 | 0.04 | 6.44 × 10−5 | ||
| HSI | AX-95651957 | 6D | 2.41 × 106 | 2.41 | 0.50 | 0.04 | 5.17 × 10−5 | MQTL6D.1 | |
| HS | AX-111558280 | 6D | 4.64 × 108 | 463.79 | 0.45 | 0.35 | 4.26 × 10−4 | ||
| HS | AX-111603870 | 7D | 4.64 × 106 | 4.64 | 0.39 | −0.33 | 6.79 × 10−4 | ||
| PH | Control | AX-111780688 | 1B | 5.30 × 108 | 529.99 | 0.17 | −8.14 | 7.10 × 10−5 | MQTL1B.5 |
| Control | AX-95235626 | 2A | 6.28 × 108 | 627.51 | 0.11 | −10.21 | 3.84 × 10−5 | ||
| HS | AX-89556873 | 2B | 7.32 × 108 | 732.05 | 0.11 | −5.49 | 7.58 × 10−5 | MQTL2B.2 | |
| HS | AX-94512274 | 3A | 5.61 × 107 | 56.09 | 0.20 | 4.36 | 3.45 × 10−5 | ||
| HS | AX-95237322 | 3D | 4.50 × 107 | 45.02 | 0.17 | 4.67 | 1.88 × 10−5 | ||
| Control | AX-95094658 | 5B | 3.98 × 108 | 398.08 | 0.08 | −11.72 | 3.16 × 10−5 | ||
| Control | AX-89381684 | 5B | 4.79 × 108 | 478.76 | 0.09 | −11.58 | 1.35 × 10−5 | MQTL5B.2 | |
| HSI | AX-86179657 | 5B | 6.70 × 108 | 670.08 | 0.13 | −0.61 | 4.43 × 10−5 | ||
| HSI | AX-108844431 | 5D | 3.24 × 107 | 32.44 | 0.10 | −0.59 | 5.70 × 10−5 | ||
| HS | AX-110726038 | 6D | 5.73 × 105 | 0.57 | 0.08 | 6.11 | 8.46 × 10−5 | ||
| HSI | AX-109431581 | 6D | 2.34 × 108 | 23.44 | 0.13 | −0.55 | 6.46 × 10−5 | ||
| HSI | AX-86177729 | 6D | 4.62 × 108 | 461.71 | 0.13 | −0.57 | 3.57 × 10−5 | ||
| HSI | AX-110911240 | 7D | 5.50 × 108 | 549.75 | 0.12 | −0.54 | 4.38 × 10−5 | ||
| SL | Control | AX-179559739 | 2D | 6.36 × 107 | 63.57 | 0.50 | 1.12 | 7.83 × 10−5 | MQTL2D.1 |
| Control | AX-109809781 | 2D | 6.06 × 108 | 606.12 | 0.10 | 1.79 | 4.89 × 10−5 | ||
| Control | AX-94522134 | 2D | 6.29 × 108 | 629 | 0.26 | 1.39 | 1.34 × 10−5 | ||
| HSI | AX-94959965 | 3A | 7.28 × 108 | 727.99 | 0.19 | −0.47 | 1.72 × 10−5 | ||
| HSI | AX-111016736 | 3B | 7.74 × 108 | 773.76 | 0.08 | −0.51 | 8.92 × 10−5 | ||
| HS | AX-179557838 | 3B | 8.22 × 108 | 822.19 | 0.44 | 0.47 | 6.78 × 10−5 | ||
| HSI | AX-94752943 | 4A | 6.04 × 108 | 603.68 | 0.06 | −0.49 | 7.20 × 10−5 | ||
| SpPS | HS | AX-179388661 | 1A | 5.94 × 108 | 593.55 | 0.33 | −1.39 | 7.02 × 10−4 | |
| Control | AX-86174804 | 1D | 7.87 × 106 | 7.87 | 0.11 | 1.35 | 9.64 × 10−4 | ||
| HS | AX-109545057 | 2B | 5.39 × 108 | 539.33 | 0.19 | −2.19 | 8.81 × 10−4 | ||
| HS | AX-94415873 | 2D | 5.51 × 108 | 550.65 | 0.24 | −1.30 | 5.48 × 10−4 | ||
| HSI | AX-94669552 | 5B | 4.42 × 107 | 44.23 | 0.12 | 0.71 | 6.21 × 10−5 | ||
| HSI | AX-95629141 | 5B | 5.66 × 107 | 56.56 | 0.09 | 0.79 | 6.39 × 10−5 | ||
| SW | Control | AX-179388053 | 2B | 1.76 × 108 | 176.29 | 0.45 | −0.42 | 6.52 × 10−5 | |
| HSI | AX-109809781 | 2D | 6.06 × 108 | 606.12 | 0.10 | −1.48 | 2.30 × 10−5 | ||
| HSI | AX-109312408 | 3D | 6.08 × 108 | 608.25 | 0.09 | −1.41 | 7.84 × 10−5 | ||
| HSI | AX-94700952 | 4B | 6.12 × 108 | 611.51 | 0.14 | −1.23 | 4.88 × 10−5 | ||
| HS | AX-110516900 | 4B | 6.70 × 108 | 669.71 | 0.07 | −0.24 | 1.32 × 10−5 | ||
| HS | AX-109482775 | 4B | 6.70 × 108 | 670.43 | 0.07 | −0.24 | 1.32 × 10−5 | ||
| HS | AX-108778179 | 6B | 5.91 × 108 | 591.21 | 0.14 | −0.17 | 4.79 × 10−5 | ||
| HSI | AX-111907482 | 6D | 4.69 × 108 | 469.1 | 0.05 | −2.12 | 2.75 × 10−5 | ||
| TKW | Control | AX-110402508 | 2A | 4.47 × 107 | 44.73 | 0.11 | −4.28 | 9.55 × 10−4 | |
| Control | AX-108895787 | 2A | 4.51 × 107 | 45.07 | 0.12 | −4.37 | 5.74 × 10−4 | ||
| Control | AX-108944653 | 2D | 6.00 × 108 | 599.89 | 0.21 | −3.36 | 6.55 × 10−4 | ||
| Control | AX-109948996 | 2D | 6.10 × 108 | 609.64 | 0.18 | −4.42 | 3.01 × 10−4 | ||
| Control | AX-109551920 | 3A | 7.28 × 108 | 727.99 | 0.08 | −9.31 | 1.60 × 10−4 | ||
| Control | AX-110589570 | 3B | 6.25 × 108 | 625.42 | 0.15 | −4.74 | 2.41 × 10−4 | ||
| HSI | AX-109270732 | 3B | 6.37 × 108 | 637.08 | 0.09 | −0.47 | 8.26 × 10−5 | ||
| HS | AX-94460648 | 6D | 4.62 × 108 | 462.36 | 0.49 | −1.84 | 6.99 × 10−5 | ||
| TpP | Control | AX-95654322 | 1A | 5.80 × 108 | 580.5 | 0.10 | 2.24 | 8.94 × 10−5 | |
| HSI | AX-109016685 | 2A | 1.91 × 107 | 19.11 | 0.20 | 0.13 | 4.13 × 10−4 | ||
| HSI | AX-95093243 | 2A | 7.72 × 108 | 771.51 | 0.08 | 0.14 | 9.39 × 10−4 | ||
| HS | AX-94760904 | 2B | 7.31 × 108 | 731.35 | 0.33 | −0.60 | 3.21 × 10−5 | ||
| HSI | AX-179477221 | 6A | 2.31 × 108 | 230.7 | 0.32 | 0.11 | 3.02 × 10−4 | ||
| HSI | AX-179475799 | 6A | 3.59 × 108 | 359.08 | 0.08 | 0.23 | 6.18 × 10−4 | ||
| HSI | AX-95016643 | 6A | 3.85 × 108 | 384.68 | 0.06 | 0.24 | 8.94 × 10−4 | ||
| HSI | AX-95119810 | 6A | 3.87 × 108 | 387.07 | 0.16 | 0.11 | 6.18 × 10−4 | ||
| HSI | AX-94949800 | 7B | 4.72 × 108 | 472.33 | 0.08 | 0.17 | 5.75 × 10−4 | ||
| HSI | AX-94665491 | 7D | 1.51 × 108 | 151.32 | 0.33 | 0.12 | 2.70 × 10−4 |
| Condition | Traits | SNP | CHR | Pos (Mb) | Effect | p-Value |
|---|---|---|---|---|---|---|
| HS | GpS | AX-110911350 | 2A | 44.18 | 2.85 | 7.46 × 10−4 |
| Control | TKW | AX-110402508 | 2A | 44.73 | −4.28 | 9.55 × 10−4 |
| HS | GpS | AX-110046675 | 2A | 44.75 | 2.85 | 7.46 × 10−4 |
| Control | TKW | AX-108895787 | 2A | 45.07 | −4.37 | 5.74 × 10−4 |
| Control | GWpS | AX-109388159 | 2B | 6.18 | −0.20 | 8.21 × 10−5 |
| HSI | DM | AX-94754683 | 2B | 6.71 | 0.13 | 9.39 × 10−5 |
| HS | TpP | AX-94760904 | 2B | 731.35 | −0.60 | 3.21 × 10−5 |
| HS | PH | AX-89556873 | 2B | 732.05 | −5.49 | 7.58 × 10−5 |
| HS | GpS | AX-179562568 | 2D | 598.71 | 2.97 | 9.43 × 10−4 |
| Control | TKW | AX-108944653 | 2D | 599.89 | −3.36 | 6.55 × 10−4 |
| HSI | SL | AX-94959965 | 3A | 727.99 | −0.47 | 1.72 × 10−5 |
| Control | TKW | AX-109551920 | 3A | 727.99 | −9.31 | 1.60 × 10−4 |
| Control | TKW | AX-110589570 | 3B | 625.42 | −4.74 | 2.41 × 10−4 |
| Control | DM | AX-109430132 | 3B | 628.80 | 3.16 | 8.93 × 10−4 |
| HS | GpS | AX-109270732 | 3B | 637.08 | 4.10 | 1.04 × 10−4 |
| HSI | TKW | AX-109270732 | 3B | 637.08 | −0.47 | 8.26 × 10−5 |
| HS | SL | AX-179557838 | 3B | 822.19 | 0.47 | 6.78 × 10−5 |
| Control | GpS | AX-89477157 | 3B | 822.97 | 2.83 | 5.42 × 10−4 |
| HS | DH | AX-111599726 | 3D | 602.27 | 3.67 | 2.36 × 10−5 |
| HS | GY | AX-111599726 | 3D | 602.27 | −0.45 | 3.26 × 10−4 |
| HIS | GY | AX-94423826 | 5B | 588.83 | −0.06 | 2.89 × 10−5 |
| HS | GpS | AX-110559492 | 5B | 591.14 | 2.07 | 6.79 × 10−4 |
| HS | DH | AX-86180088 | 6A | 270.61 | 3.50 | 7.96 × 10−5 |
| Control | DM | AX-86180088 | 6A | 270.61 | 2.96 | 6.43 × 10−4 |
| HS | DH | AX-94744160 | 6A | 381.03 | 3.50 | 7.96 × 10−5 |
| HSI | TpP | AX-95016643 | 6A | 384.68 | 0.24 | 8.94 × 10−4 |
| HS | DM | AX-110984345 | 6A | 384.73 | 4.75 | 6.89 × 10−5 |
| HS | DH | AX-109950287 | 6A | 390.08 | 3.70 | 3.85 × 10−5 |
| Control | DM | AX-109950287 | 6A | 390.08 | 3.11 | 4.03 × 10−4 |
| HSI | DH | AX-86173990 | 6D | 458.36 | 0.08 | 5.48137 × 10−5 |
| HSI | PH | AX-86177729 | 6D | 461.71 | −0.57 | 3.57 × 10−5 |
| HS | TKW | AX-94460648 | 6D | 462.36 | −1.84 | 6.99 × 10−5 |
| HSI | DH | AX-110389623 | 6D | 462.70 | 0.08 | 9.35026 × 10−5 |
| HS | GY | AX-111558280 | 6D | 463.79 | 0.35 | 4.26 × 10−4 |
| HS | DH | AX-94980563 | 7B | 437.93 | 4.08 | 1.97 × 10−5 |
| Control | DM | AX-94980563 | 7B | 437.93 | 3.27 | 4.85 × 10−4 |
| Traits | SNP | CHR | Pos(Mb) | Hsp | Gene ID | POS(Mb)2 |
|---|---|---|---|---|---|---|
| GpS | AX-110533769 | 2B | 717.22 | TaHSP40.54 | TraesCS2B01G526300 | 720.58 |
| TpP | AX-94760904 | 2B | 731.35 | TaHSP70.24 | TraesCS2B01G535000 | 731.25 |
| PH | AX-94512274 | 3A | 56.09 | TaHSP40.63 | TraesCS3A01G083700 | 53.83 |
| PH | AX-95237322 | 3D | 45.02 | TaHSP40.95 | TraesCS3D01G083700 | 42.49 |
| SW | AX-109482775 | 4B | 670.43 | TaHSP70.52 | TraesCS4B01G397600 | 671.74 |
| GWpS | AX-109911103 | 5A | 611.65 | TaHSP40.150 | TraesCS5A01G426100 | 611.34 |
| GWpS | AX-94886530 | 5B | 186.84 | TaHSP40.155 | TraesCS5B01G114500 | 185.72 |
| TKW | AX-94460648 | 6D | 462.36 | TaHSP60.69 | TraesCS6D01G383500 | 462.46 |
| Haplotype | Xhau1 | Xhau2 | Xhau3 | Xhau4 | Xhau5 | Deleted Sites | No. of Lines | Frequency % |
|---|---|---|---|---|---|---|---|---|
| Hap1 | + | + | + | 127 | + | 0 | 33 | 25 |
| Hap2 | - | - | 127 | + | 3 | 21 | 16 | |
| Hap3 | - | + | + | 127 | + | 1 | 20 | 15 |
| Hap4 | - | + | + | 127 | - | 2 | 13 | 10 |
| Hap5 | - | - | - | 127 | - | 4 | 13 | 10 |
| Hap6 | + | + | + | 127 | - | 1 | 10 | 8 |
| Hap7 | - | - | + | 127 | + | 2 | 6 | 5 |
| Hap8 | - | - | - | 195 | - | 4 | 5 | 4 |
| Hap9 | - | + | - | 127 | + | 2 | 3 | 2 |
| Hap10 | + | + | - | 127 | + | 1 | 2 | 2 |
| Hap11 | - | - | - | 195 | + | 3 | 2 | 2 |
| Hap12 | - | - | - | - | - | 5 | 1 | 1 |
| Hap13 | + | + | - | 127 | - | 2 | 1 | 1 |
| Hap14 | + | - | + | 127 | + | 1 | 1 | 1 |
| Hap15 | - | - | + | 195 | + | 2 | 1 | 1 |
| Total | 132 | 100 |
| Primer | Sequence | Tm °C | Product Size (bp) |
|---|---|---|---|
| Xhau1-F | GGGAGTGTTTGTGTGAGGATTTG | 65 | 825 |
| Xhau1-R | GCACTACTACCAAACCACGTGTA | ||
| Xhau2-F | GGGAGCCAATTCGTGTGACT | 56 | 426 |
| Xhau2-R | CAAGCGCTATACAACTGTGCT | ||
| Xhau3-F | GCCCGTGAATCATACTTGAGCG | 58 | 788 |
| Xhau3-R | TGAGGAGATAATTGTACGCCGA | ||
| Xhau4-F | TCGGTTGGTTTGTTTATACTTGC | 65 | 127/195 |
| Xhau4-R | CCACGCTTGCACAATCTATTCT | ||
| Xhau5-F | TGCCTACCAAAGTGAGACCTG | 60 | 259 |
| Xhau5-R | ACCTACCTCTACCTCAACCCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mehvish, A.; Aziz, A.; Bukhari, B.; Qayyum, H.; Mahmood, Z.; Baber, M.; Sajjad, M.; Pang, X.; Wang, F. Identification of Single-Nucleotide Polymorphisms (SNPs) Associated with Heat Tolerance at the Reproductive Stage in Synthetic Hexaploid Wheats Using GWAS. Plants 2023, 12, 1610. https://doi.org/10.3390/plants12081610
Mehvish A, Aziz A, Bukhari B, Qayyum H, Mahmood Z, Baber M, Sajjad M, Pang X, Wang F. Identification of Single-Nucleotide Polymorphisms (SNPs) Associated with Heat Tolerance at the Reproductive Stage in Synthetic Hexaploid Wheats Using GWAS. Plants. 2023; 12(8):1610. https://doi.org/10.3390/plants12081610
Chicago/Turabian StyleMehvish, Ambreen, Abdul Aziz, Birra Bukhari, Humaira Qayyum, Zahid Mahmood, Muhammad Baber, Muhammad Sajjad, Xuequn Pang, and Fenglan Wang. 2023. "Identification of Single-Nucleotide Polymorphisms (SNPs) Associated with Heat Tolerance at the Reproductive Stage in Synthetic Hexaploid Wheats Using GWAS" Plants 12, no. 8: 1610. https://doi.org/10.3390/plants12081610