
The Mechanism of Resistance of EUROPEAN Plum to Plum pox virus Mediated by Hypersensitive Response Is Linked to VIRAL NIa and Its Protease Activity
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
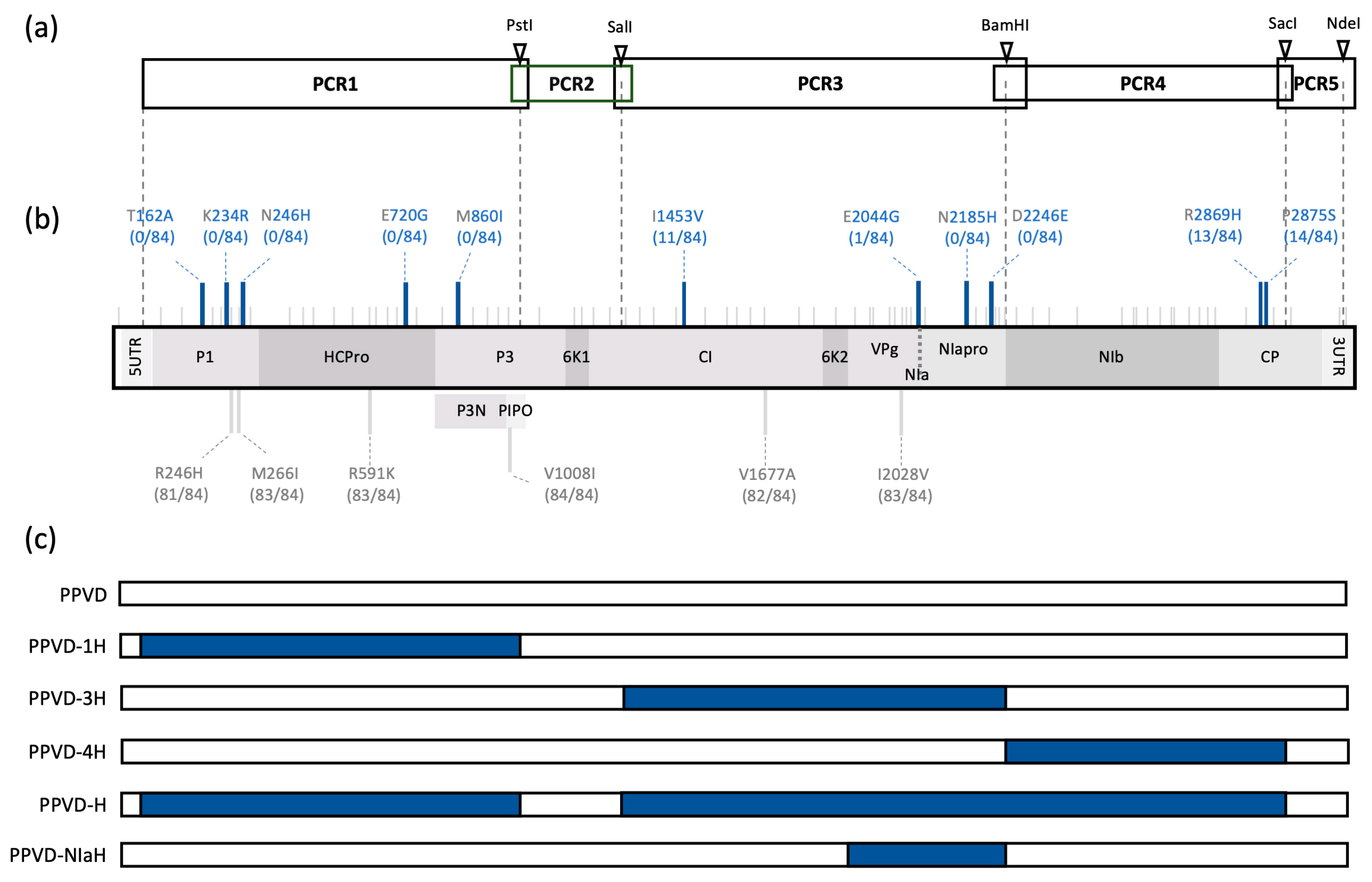
2.1. Genome Sequence Analysis of the HR-Escaping PPV Isolate
2.2. PPVD-H Escapes the Hypersensitive Response of Resistant Plum Causing Typical PPV Symptomatology
2.3. The NIa Region of PPVD-H Is Sufficient to Avoid the Hypersensitive Response of Resistant Plum
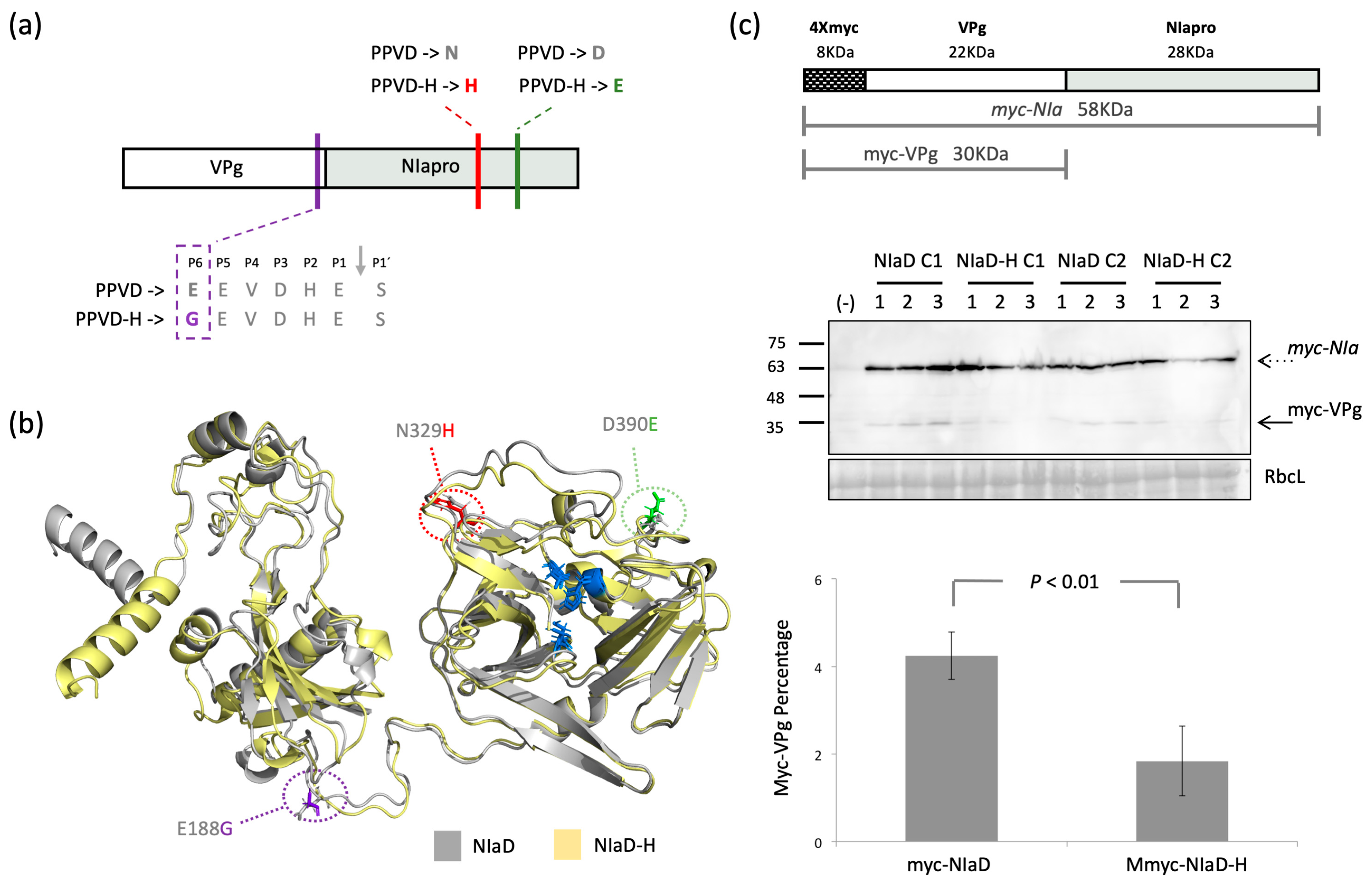
2.4. All Three Amino Acid Changes Found in PPVD-H NIa Are Necessary to Escape the Hypersensitive Response of Resistant Plum
2.5. Cleavage between Vpg and NIapro Is Modulated by the Amino Acid Changes Found in NIa
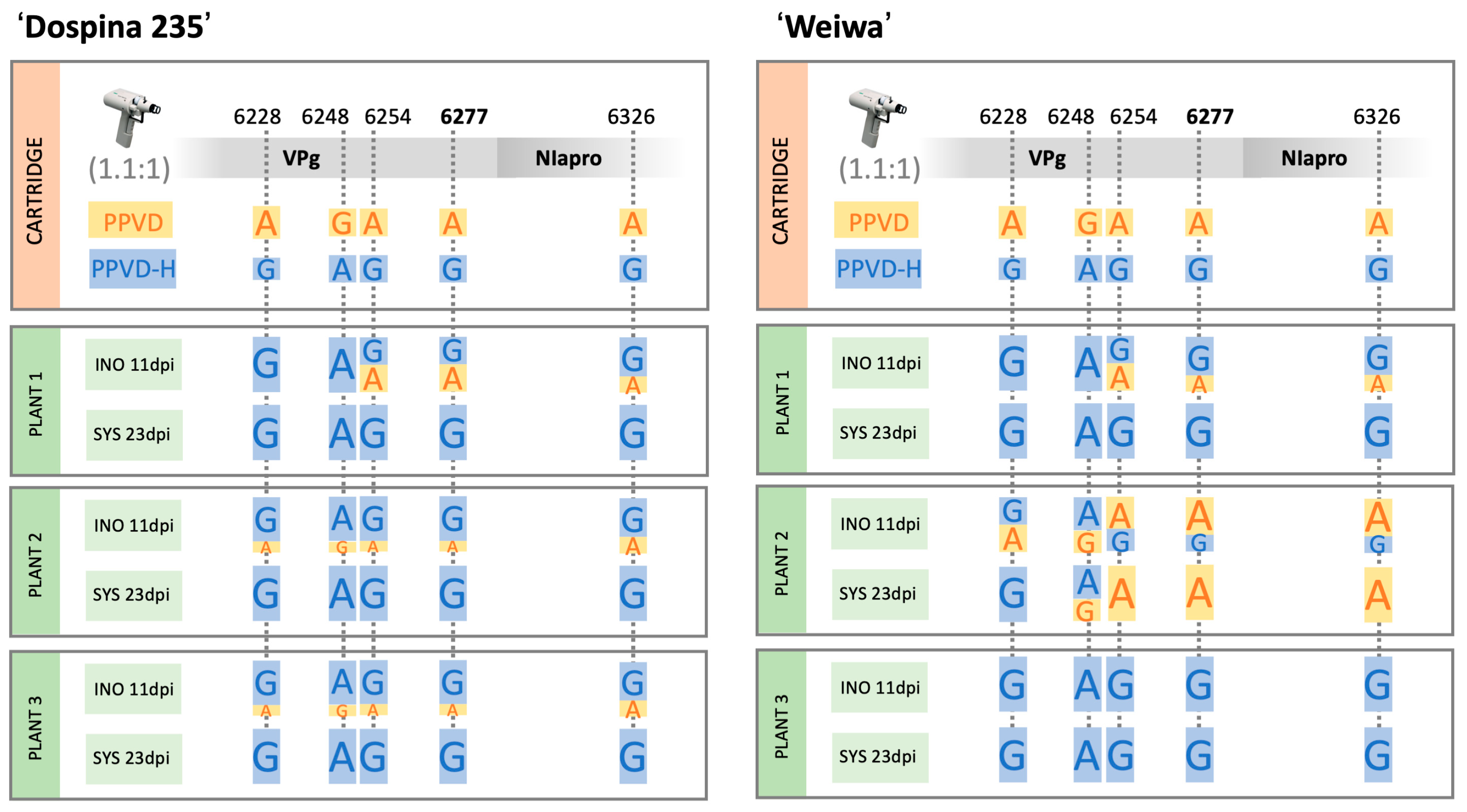
2.6. PPVD-H Outcompetes PPVD in Resistant Plants
3. Discussion
4. Materials and Methods
4.1. Cloning of PPVD-H and Its Variants
4.2. Plasmids Used for Transient Expression and Agroinfiltration
4.3. Biolistic Inoculation and RT-PCR Analysis
4.4. Western Blot Analysis
4.5. In Silico Study of PPV-D Alignment and NIa Structure
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cirilli, M.; Geuna, F.; Babini, A.R.; Bozhkova, V.; Catalano, L.; Cavagna, B.; Dallot, S.; Decroocq, V.; Dondini, L.; Foschi, S.; et al. Fighting sharka in peach: Current limitations and future perspectives. Front. Plant Sci. 2016, 7, 1290. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Moreno, M.J.; Hernández, J.A.; Díaz-Vivancos, P. Sharka: How do plants respond to Plum pox virus infection? J. Exp. Bot. 2015, 66, 25–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šubr, Z.; Glasa, M. Unfolding the secrets of plum pox virus: From epidemiology to genomics. Acta Virol. 2013, 57, 217–228. [Google Scholar] [CrossRef] [Green Version]
- Garcia, J.A.; Glasa, M.; Cambra, M.; Candresse, T. Plum pox virus and sharka: A model potyvirus and a major disease. Mol. Plant Pathol. 2014, 15, 226–241. [Google Scholar] [CrossRef]
- Atanasoff. Plum pox. A new virus disease. Yearb. Univ. Sofia Fac. Agric. 1932, 11, 49–69. [Google Scholar]
- Cambra, M.; Capote, N.; Myrta, A.; Llácer, G. Plum pox virus and the estimated costs associated with sharka disease. EPPO Bull. 2006, 36, 202–204. [Google Scholar] [CrossRef]
- Milosevic, T.; Milosevic, N.; Mladenovic, J.; Jevremovic, D. Impact of Sharka disease on tree growth, productivity and fruit quality of apricot (Prunus armeniaca L.). Sci. Hortic. 2019, 244, 270–276. [Google Scholar] [CrossRef]
- Rodamilans, B.; Valli, A.; García, J.A. Molecular Plant-Plum Pox Virus Interactions. Mol. Plant-Microbe Interact. 2020, 33, 6–17. [Google Scholar] [CrossRef]
- Valli, A.; García, J.A.; López-Moya, J.J. Potyviruses (Potyviridae); Academic Press: Oxford, UK, 2021; Volume 3. [Google Scholar]
- Chung, B.Y.; Miller, W.A.; Atkins, J.F.; Firth, A.E. An overlapping essential gene in the Potyviridae. Proc. Natl. Acad. Sci. USA 2008, 105, 5897–5902. [Google Scholar] [CrossRef] [Green Version]
- Hagiwara-Komoda, Y.; Choi, S.H.; Sato, M.; Atsumi, G.; Abe, J.; Fukuda, J.; Honjo, M.N.; Nagano, A.J.; Komoda, K.; Nakahara, K.S.; et al. Truncated yet functional viral protein produced via RNA polymerase slippage implies underestimated coding capacity of RNA viruses. Sci. Rep. 2016, 6, 21411. [Google Scholar] [CrossRef]
- Olspert, A.; Chung, B.Y.; Atkins, J.F.; Carr, J.P.; Firth, A.E. Transcriptional slippage in the positive-sense RNA virus family Potyviridae. EMBO Rep. 2015, 16, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Rodamilans, B.; Valli, A.; Mingot, A.; San León, D.; Baulcombe, D.; López-Moya, J.J.; García, J.A. RNA polymerase slippage as a mechanism for the production of frameshift gene products in plant viruses of the Potyviridae family. J. Virol. 2015, 89, 6965–6967. [Google Scholar] [CrossRef] [Green Version]
- Rodamilans, B.; Shan, H.; Pasin, F.; Garcia, J.A. Plant Viral Proteases: Beyond the Role of Peptide Cutters. Front. Plant Sci. 2018, 9, 666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodamilans, B.; Valli, A.; García, J.A. Mechanistic divergence between P1 proteases of the family Potyviridae. J. Gen. Virol. 2013, 94, 1407–1414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valli, A.A.; Gallo, A.; Rodamilans, B.; López-Moya, J.J.; García, J.A. The HCPro from the Potyviridae family: An enviable multitasking Helper Component that every virus would like to have. Mol. Plant Pathol. 2018, 19, 744–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pasin, F.; Daros, J.A.; Tzanetakis, I.E. Proteome expansion in the Potyviridae evolutionary radiation. FEMS Microbiol. Rev. 2022, 46, fuac011. [Google Scholar] [CrossRef]
- Riechmann, J.L.; Cervera, M.T.; Garcia, J.A. Processing of the plum pox virus polyprotein at the P3-6K1 junction is not required for virus viability. J. Gen. Virol. 1995, 76 Pt 4, 951–956. [Google Scholar] [CrossRef]
- Calvo, M.; Malinowski, T.; Garcia, J.A. Single amino acid changes in the 6K1-CI region can promote the alternative adaptation of Prunus- and Nicotiana-propagated Plum pox virus C isolates to either host. Mol. Plant Microbe Interact. 2014, 27, 136–149. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.N.; Tang, R.Q.; Zhang, Y.; Peng, J.; Xian, O.; Zhang, Z.H.; Zhang, S.B.; Zhang, D.Y.; Liu, H.; Luo, X.W.; et al. The NIa-Protease Protein Encoded by the Pepper Mottle Virus Is a Pathogenicity Determinant and Releases DNA Methylation of Nicotiana benthamiana. Front. Microbiol. 2020, 11, 102. [Google Scholar] [CrossRef] [Green Version]
- Faurez, F.; Baldwin, T.; Tribodet, M.; Jacquot, E. Identification of new Potato virus Y (PVY) molecular determinants for the induction of vein necrosis in tobacco. Mol. Plant Pathol. 2012, 13, 948–959. [Google Scholar] [CrossRef]
- Mestre, P.; Brigneti, G.; Baulcombe, D.C. An Ry-mediated resistance response in potato requires the intact active site of the NIa proteinase from potato virus Y. Plant J. 2000, 23, 653–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mestre, P.; Brigneti, G.; Durrant, M.C.; Baulcombe, D.C. Potato virus Y NIa protease activity is not sufficient for elicitation of Ry-mediated disease resistance in potato. Plant J. 2003, 36, 755–761. [Google Scholar] [CrossRef] [PubMed]
- Cambra, M.; Vidal, E. Sharka, a vector-borne disease caused by Plum pox virus: Vector species, transmission mechanism, epidemiology and mitigation strategies to reduce its natural spread. Acta Hortic. 2017, 1163, 57–68. [Google Scholar] [CrossRef]
- Gildow, F.; Damsteegt, V.; Stone, A.; Schneider, W.; Luster, D.; Levy, L. Plum pox in north america: Identification of aphid vectors and a potential role for fruit in virus spread. Phytopathology 2004, 94, 868–874. [Google Scholar] [CrossRef] [Green Version]
- Labonne, G.; Ybon, M.; Quiot, J.B.; Avinert, L.; Llacer, G. Aphids as potential vectors of Plum pox virus: Comparison of methods of testing and epidemiological consequences. Acta Hortic. 1995, 386, 207–218. [Google Scholar] [CrossRef]
- Sihelská, N.; Glasa, M.; Šubr, W. Host preference of the major strains of Plum pox virus—Opinions based on regional and world-wide sequence data. J. Integr. Agric. 2017, 16, 510–515. [Google Scholar] [CrossRef]
- Llácer, G.; Cambra, M. Hosts and symptoms of Plum pox virus: Fruiting Prunus species. EPPO Bull. 2006, 36, 219–221. [Google Scholar] [CrossRef]
- Lichtenegger, L.; Neumüller, M.; Treutter, D.; Hartmann, W. The inheritance of the hypersensitivity resistance of European plum (Prunus domestica L.) against the Plum pox virus. Julius-Kühn-Arch. 2010, 427, 327–329. [Google Scholar]
- Neumüller, M.; Hartmann, W. The phenotypically quantitative nature of hypersensitivity of European plum (Prunus domestica L.) against the Plum pox virus and its description using the hypersensitivity index. Hortic. Sci. 2008, 35, 50–64. [Google Scholar] [CrossRef] [Green Version]
- Neumüller, M.; Hartmann, W.; Petruschke, M.; Treutter, D. The hypersensitivity resistance of european plum to the Plum pox virus and its potential impact on the epidemiology of the virus. Julius-Kuhn-Arch. 2010, 427, 147–150. [Google Scholar]
- Neumüller, M.; Hartmann, W.; Stösser, R. The hypersensitivity of European plum against Plum pox virus (PPV) as a promising mechanism of resistance. Phytopathol. Pol. 2005, 36, 77–83. [Google Scholar]
- Rodamilans, B.; San León, D.; Mühlberger, L.; Candresse, T.; Neumüller, M.; Oliveros, J.C.; García, J.A. Transcriptomic analysis of Prunus domestica undergoing hypersensitive response to Plum pox virus infection. PLoS ONE 2014, 9, e100477. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fernández, M.R.; Mouriño, M.; Rivera, J.; Rodríguez, F.; Plana-Durán, J.; García, J.A. Protection of rabbits against rabbit hemorrhagic disease virus by immunization with the VP60 protein expressed in plants with a potyvirus-based vector. Virology 2001, 280, 283–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plum Pox Virus Database. Sharco: Sharka Containment in View of Eu Expansion. Available online: http://w3.pierroton.inra.fr:806h (accessed on 27 June 2014).
- Mann, K.S.; Sanfaçon, H. Expanding Repertoire of Plant Positive-Strand RNA Virus Proteases. Viruses 2019, 11, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shan, H.; Pasin, F.; Tzanetakis, I.E.; Simón-Mateo, C.; García, J.A.; Rodamilans, B. Truncation of a P1 leader proteinase facilitates potyvirus replication in a non-permissive host. Mol. Plant Pathol. 2018, 19, 1504–1510. [Google Scholar] [CrossRef] [Green Version]
- Pasin, F.; Simón-Mateo, C.; García, J.A. The hypervariable amino-terminus of P1 protease modulates potyviral replication and host defense responses. PLoS Pathog. 2014, 10, e1003985. [Google Scholar] [CrossRef]
- García, J.A.; Martín, M.T.; Cervera, M.T.; Riechmann, J.L. Proteolytic processing of the plum pox potyvirus polyprotein by the NIa protease at a novel cleavage site. Virology 1992, 188, 697–703. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Jones, R.A.C.; Naidu, R.A. Global Dimensions of Plant Virus Diseases: Current Status and Future Perspectives. Annu. Rev. Virol. 2019, 6, 387–409. [Google Scholar] [CrossRef]
- Revers, F.; Nicaise, V. Plant resistance to infection by viruses. eLS 2014, 1–10. [Google Scholar] [CrossRef]
- Phan, J.; Zdanov, A.; Evdokimov, A.G.; Tropea, J.E.; Peters, H.K., 3rd; Kapust, R.B.; Li, M.; Wlodawer, A.; Waugh, D.S. Structural basis for the substrate specificity of tobacco etch virus protease. J. Biol. Chem. 2002, 277, 50564–50572. [Google Scholar] [CrossRef] [Green Version]
- Xiao, H.; Lord, E.; Sanfaçon, H. Proteolytic processing of plant proteins by potyvirus NIa proteases. J. Virol. 2022, 96, e0144421. [Google Scholar] [CrossRef]
- Jagdeo, J.M.; Dufour, A.; Klein, T.; Solis, N.; Kleifeld, O.; Kizhakkedathu, J.; Luo, H.; Overall, C.M.; Jan, E. N-Terminomics TAILS Identifies Host Cell Substrates of Poliovirus and Coxsackievirus B3 3C Proteinases That Modulate Virus Infection. J. Virol. 2018, 92, e02211-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ade, J.; DeYoung, B.J.; Golstein, C.; Innes, R.W. Indirect activation of a plant nucleotide binding site-leucine-rich repeat protein by a bacterial protease. Proc. Natl. Acad. Sci. USA 2007, 104, 2531–2536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, F.; Golstein, C.; Ade, J.; Stoutemyer, M.; Dixon, J.E.; Innes, R.W. Cleavage of Arabidopsis PBS1 by a bacterial type III effector. Science 2003, 301, 1230–1233. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Nakamura, S.; Kawamukai, M.; Koizumi, N.; Nakagawa, T. Development of a series of gateway binary vectors possessing a tunicamycin resistance gene as a marker for the transformation of Arabidopsis thaliana. Biosci. Biotechnol. Biochem. 2011, 75, 804–807. [Google Scholar] [CrossRef]
- Valli, A.; Martín-Hernández, A.M.; López-Moya, J.J.; García, J.A. RNA silencing suppression by a second copy of the P1 serine protease of Cucumber vein yellowing ipomovirus, a member of the family Potyviridae that lacks the cysteine protease HCPro. J. Virol. 2006, 80, 10055–10063. [Google Scholar] [CrossRef] [Green Version]
- Pasin, F.; Bedoya, L.C.; Bernabé-Orts, J.M.; Gallo, A.; Simón-Mateo, C.; Orzaez, D.; García, J.A. Multiple T-DNA Delivery to Plants Using Novel Mini Binary Vectors with Compatible Replication Origins. ACS Synth. Biol. 2017, 6, 1962–1968. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodamilans, B.; Hadersdorfer, J.; Berki, Z.; García, B.; Neumüller, M.; García, J.A. The Mechanism of Resistance of EUROPEAN Plum to Plum pox virus Mediated by Hypersensitive Response Is Linked to VIRAL NIa and Its Protease Activity. Plants 2023, 12, 1609. https://doi.org/10.3390/plants12081609
Rodamilans B, Hadersdorfer J, Berki Z, García B, Neumüller M, García JA. The Mechanism of Resistance of EUROPEAN Plum to Plum pox virus Mediated by Hypersensitive Response Is Linked to VIRAL NIa and Its Protease Activity. Plants. 2023; 12(8):1609. https://doi.org/10.3390/plants12081609
Chicago/Turabian StyleRodamilans, Bernardo, Johannes Hadersdorfer, Zita Berki, Beatriz García, Michael Neumüller, and Juan Antonio García. 2023. "The Mechanism of Resistance of EUROPEAN Plum to Plum pox virus Mediated by Hypersensitive Response Is Linked to VIRAL NIa and Its Protease Activity" Plants 12, no. 8: 1609. https://doi.org/10.3390/plants12081609