

Influence of Temperature on Seed Germination of Five Wild-Growing Tulipa Species of Greece Associated with Their Ecological Profiles: Implications for Conservation and Cultivation

 , , , , , ,
, , , , , , 
Abstract
:1. Introduction
2. Results
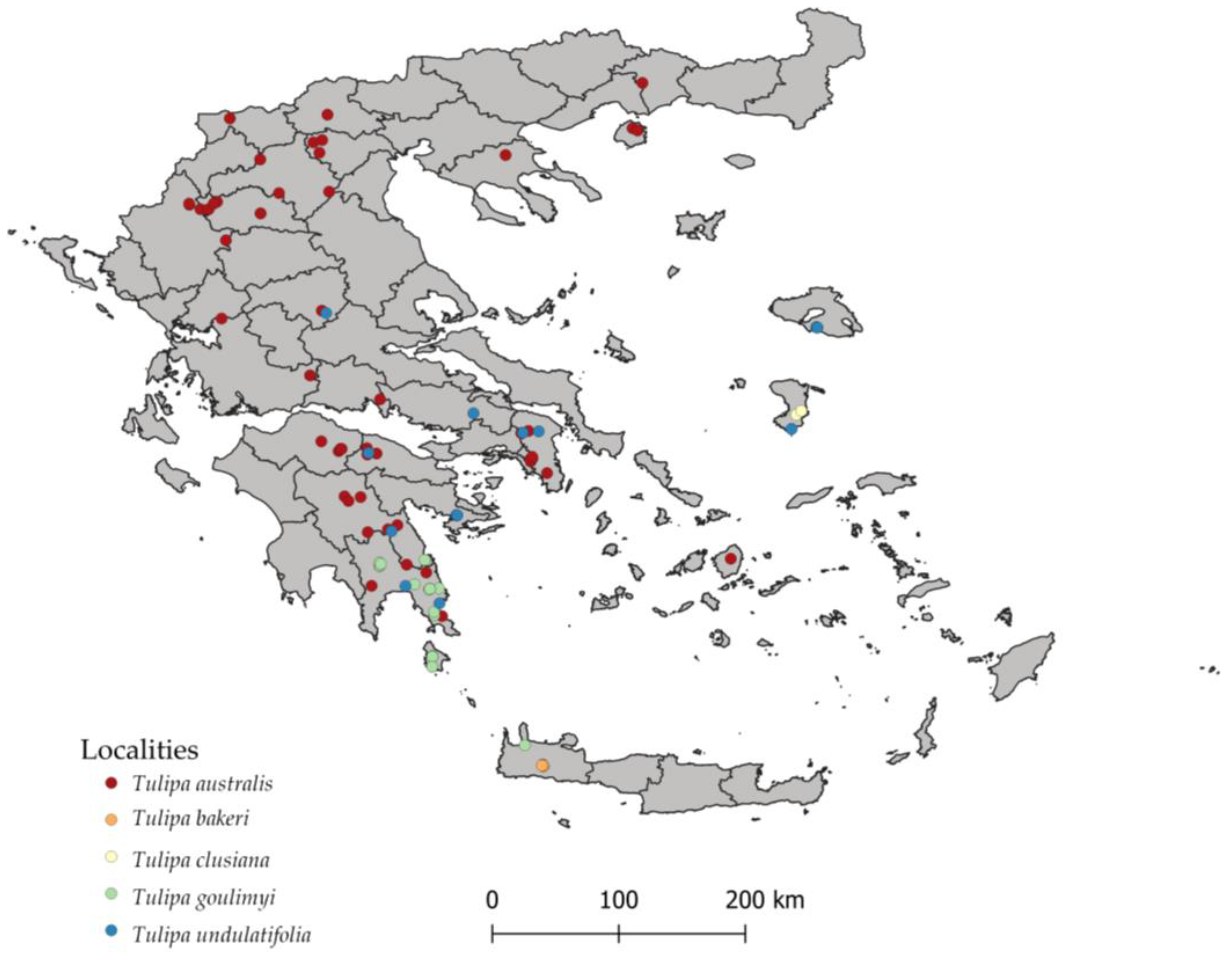
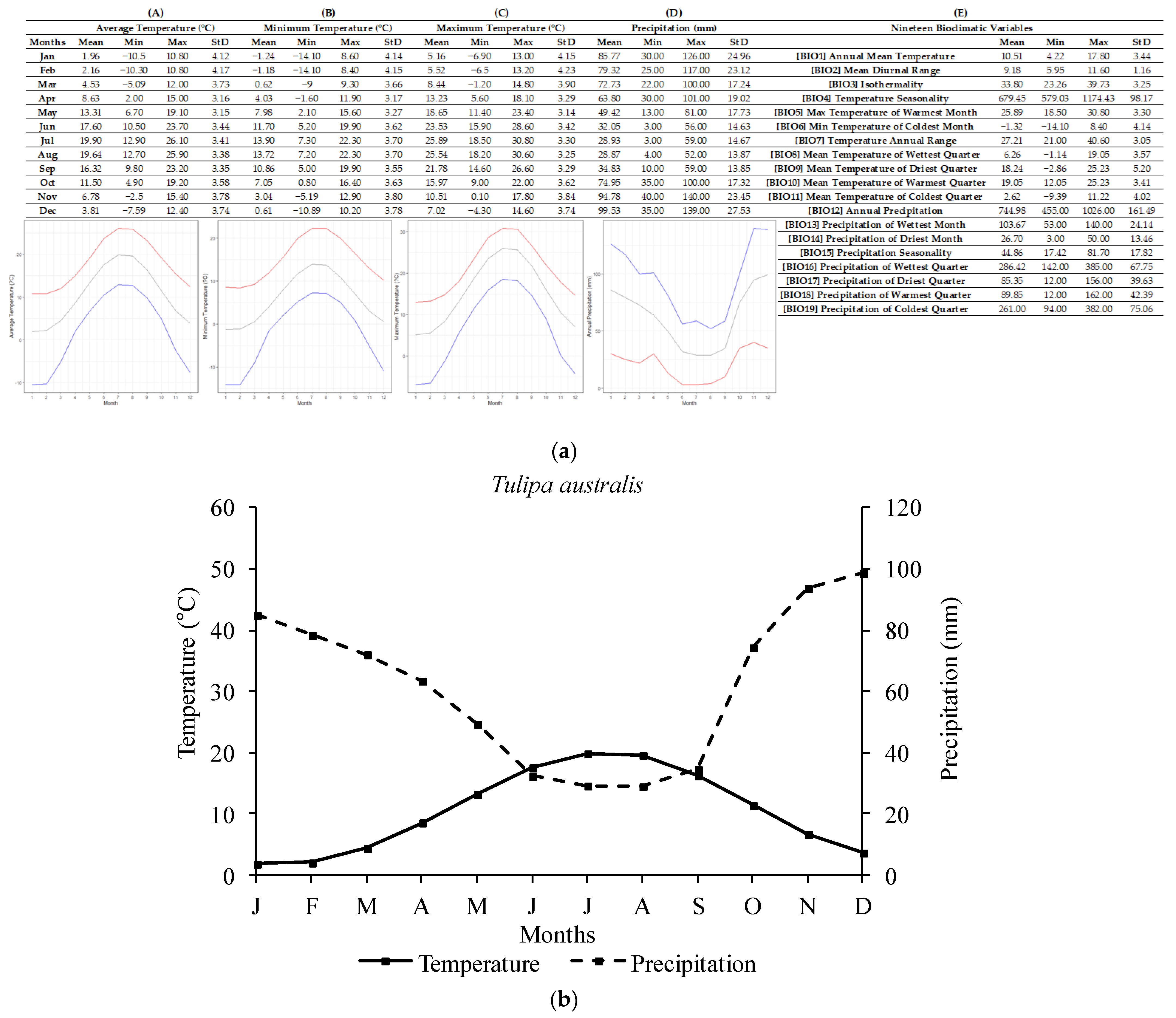
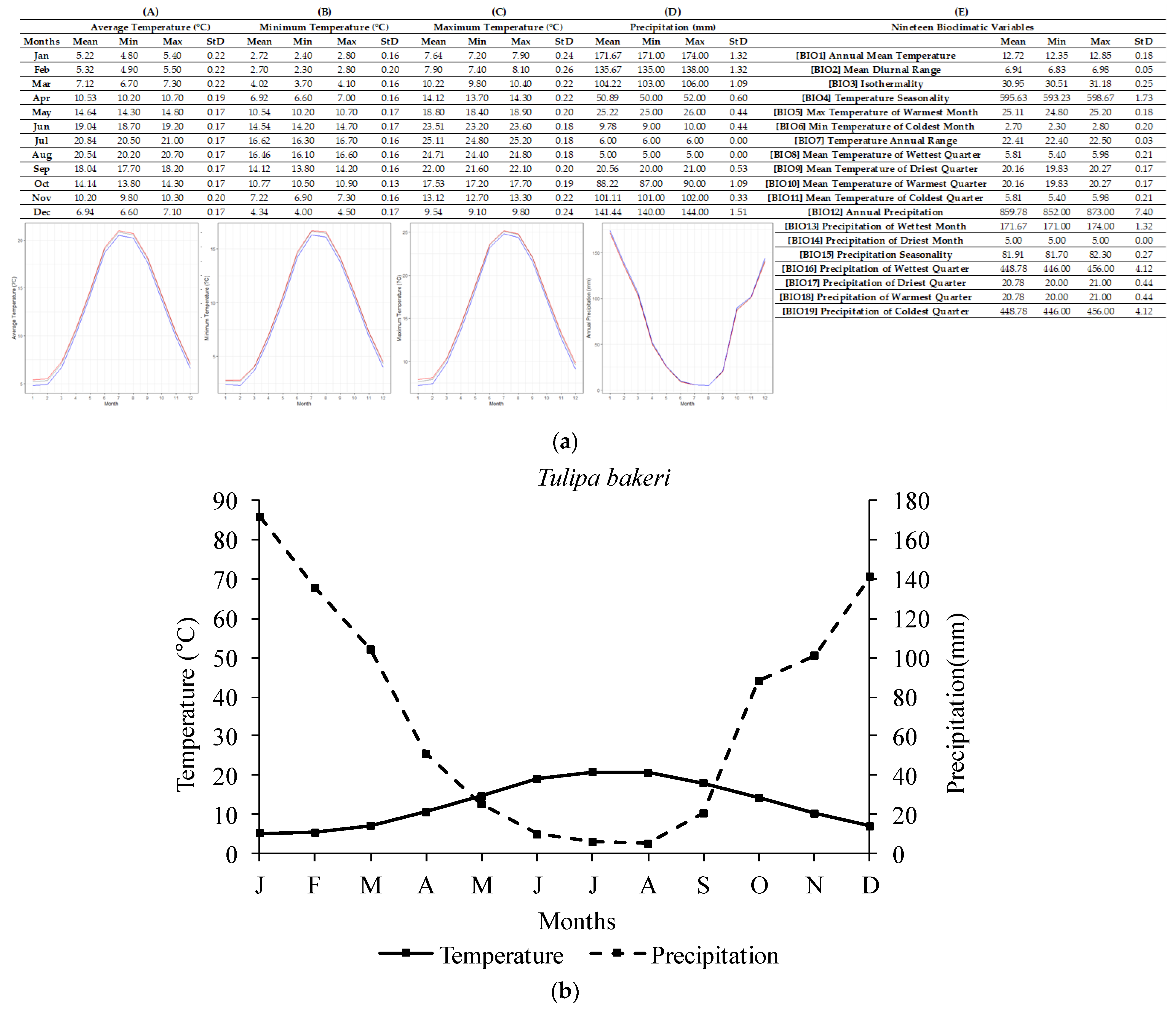
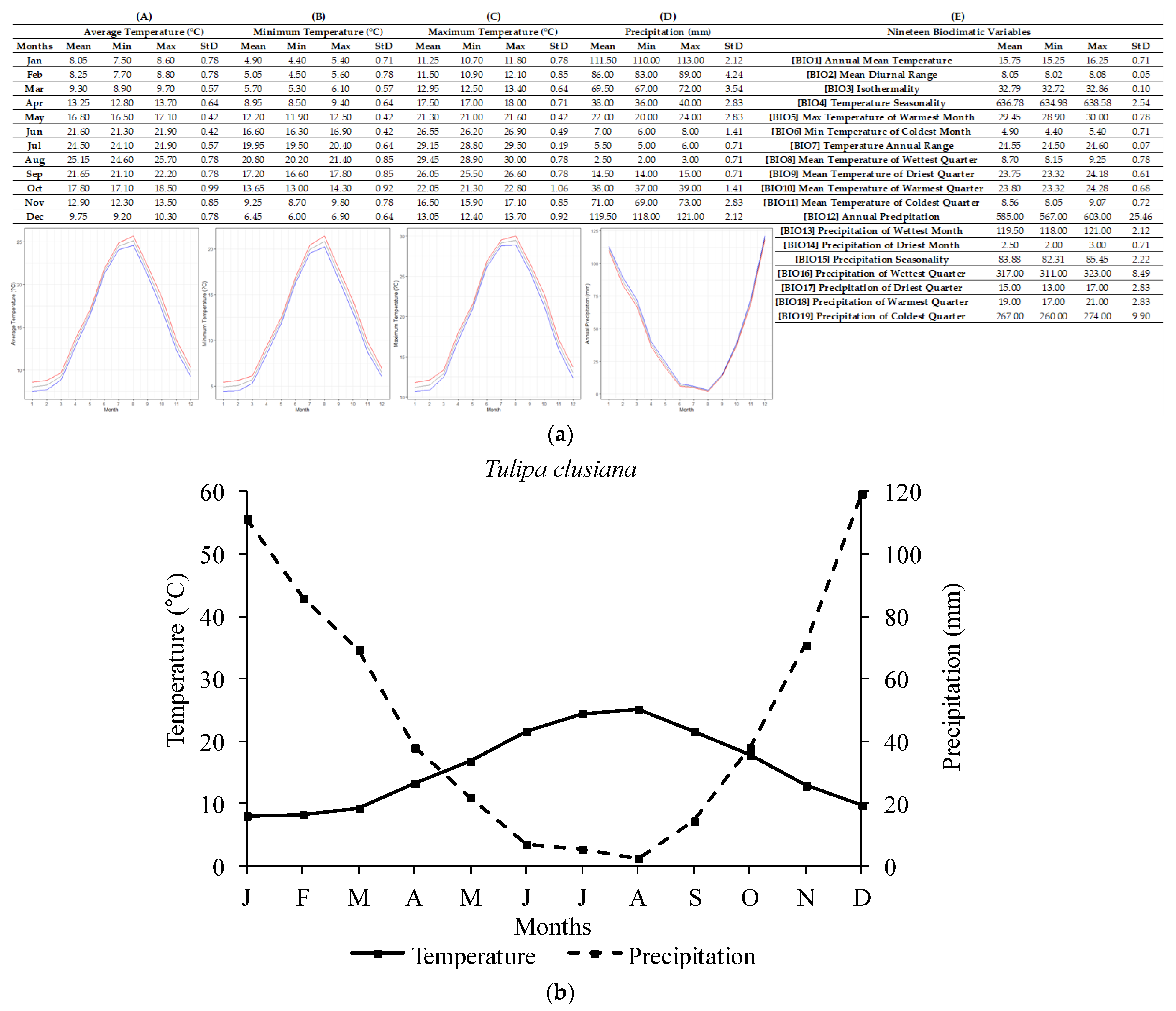
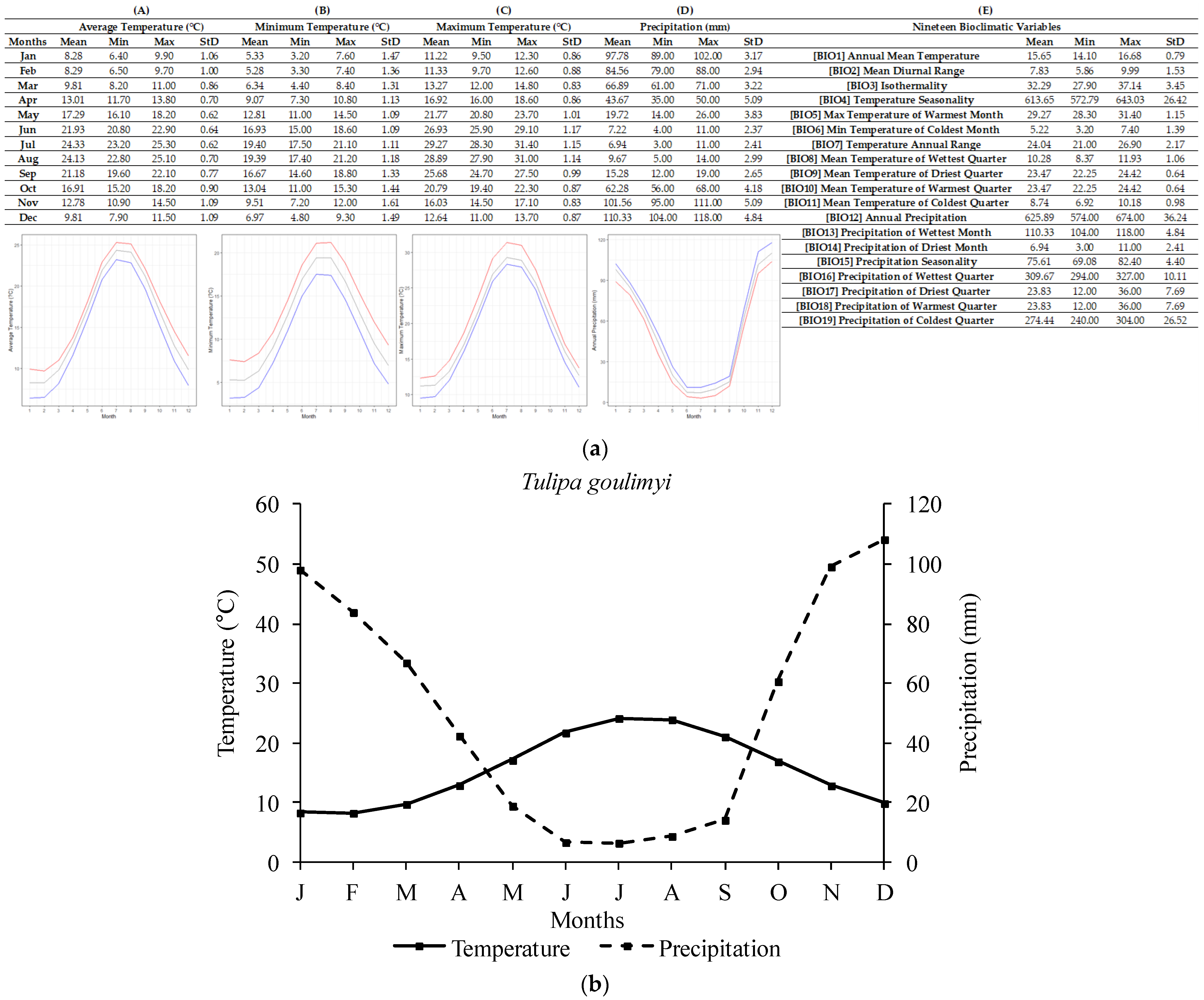
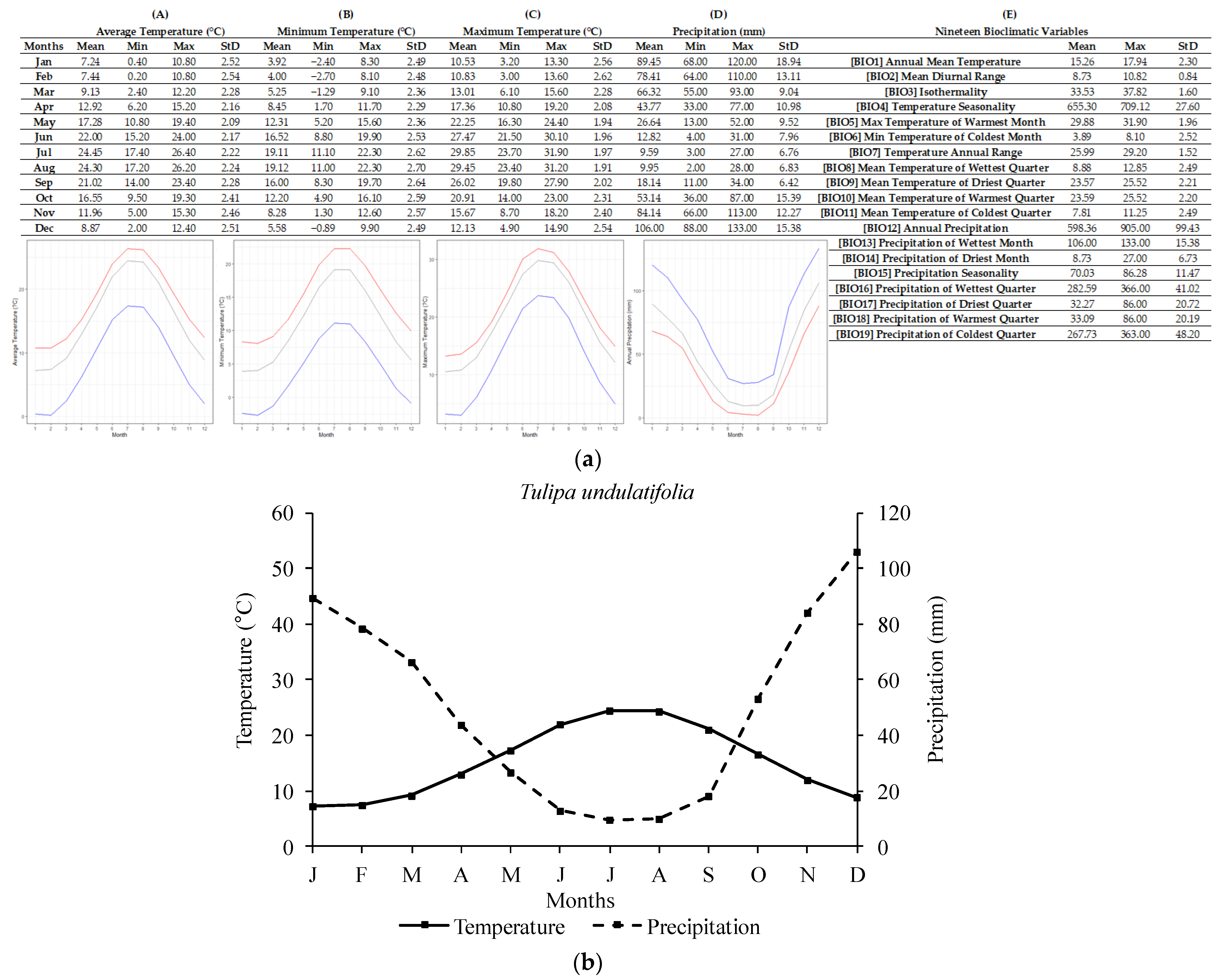
2.1. Ecological Profiling of Studied Species
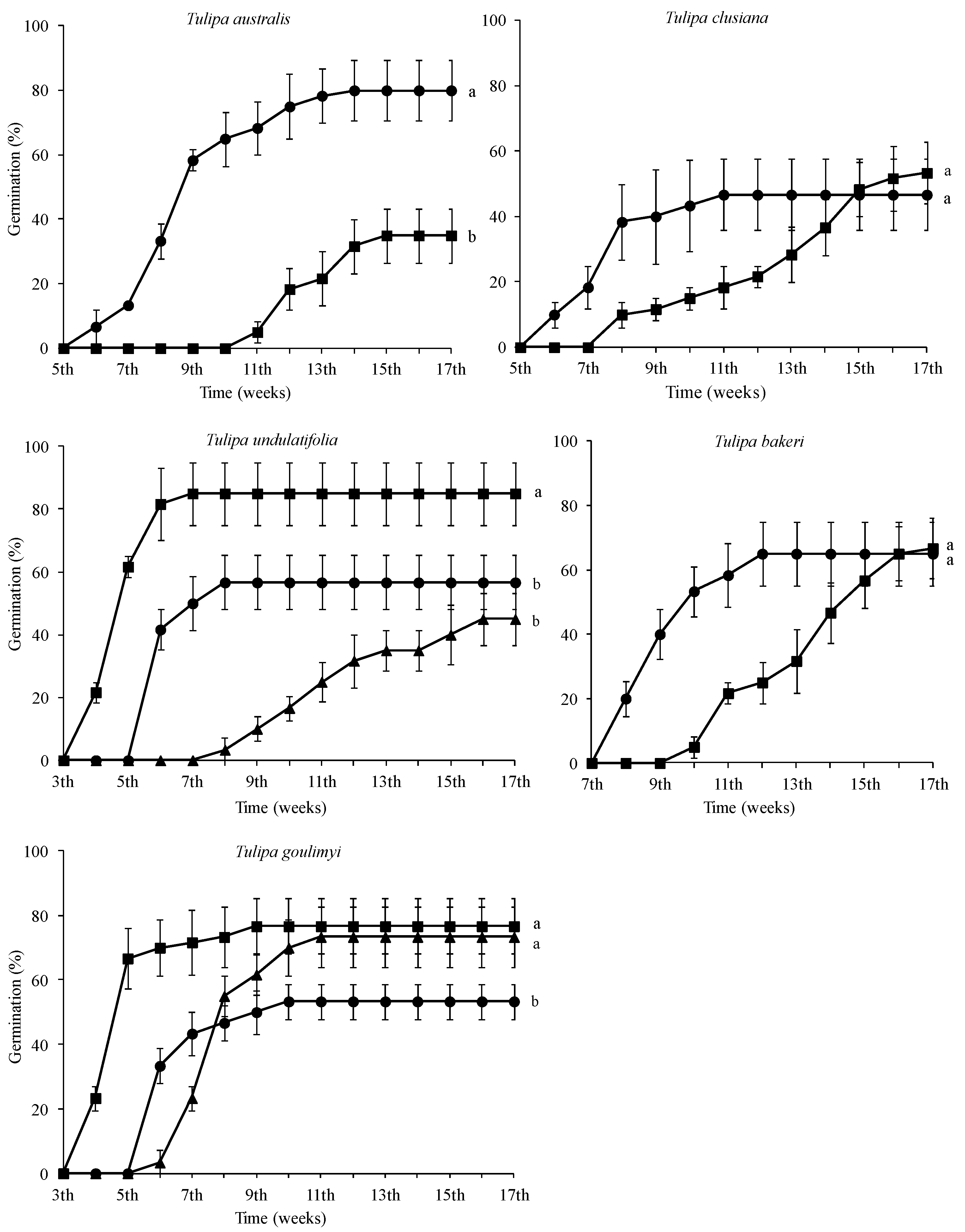
2.2. Seed Germination Tests
2.3. Seedling and Bulblet Production
3. Discussion
4. Materials and Methods
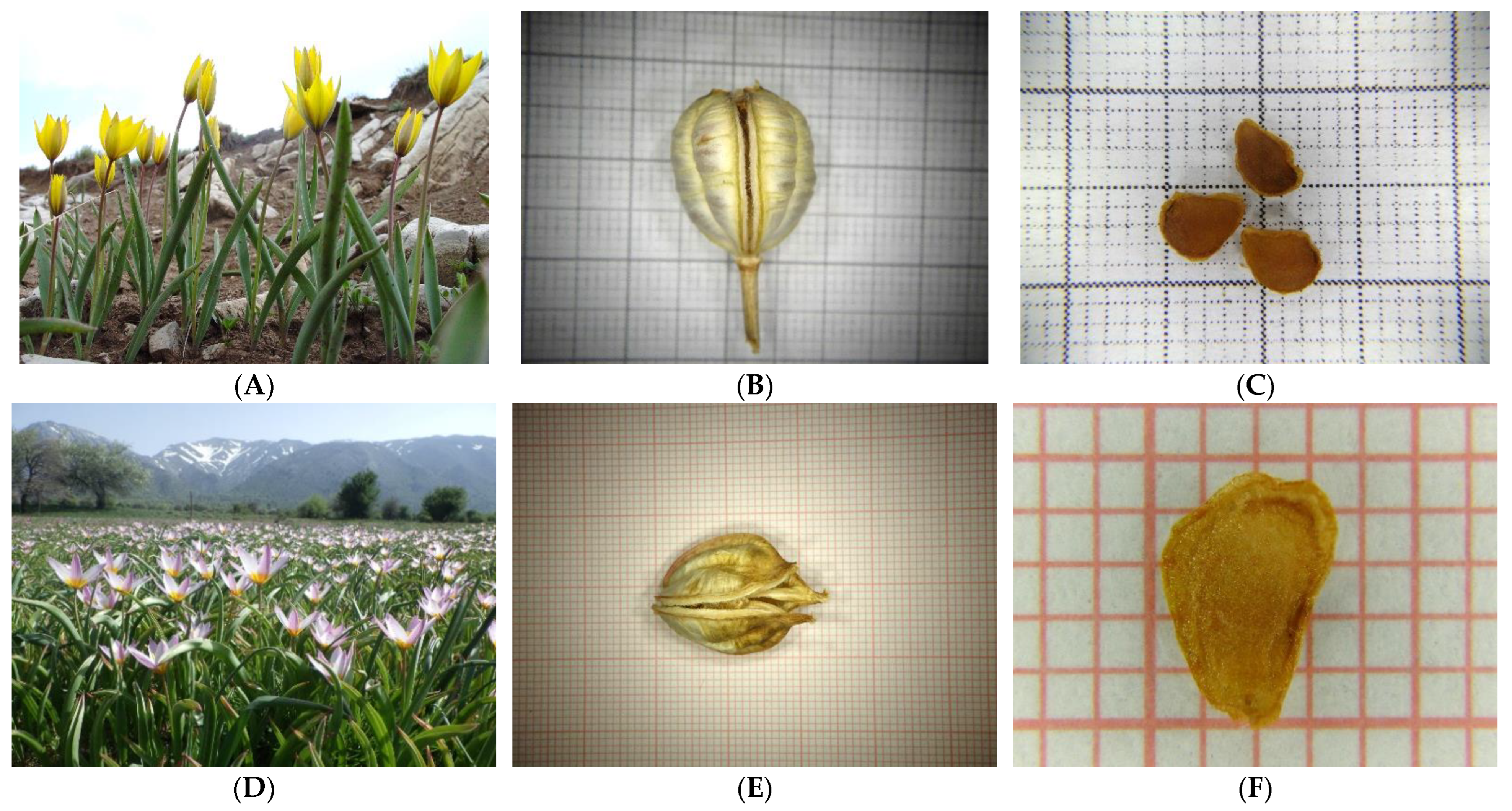
4.1. Focal Greek Tulips and Seed Collection Data
4.2. Distribution Data
4.3. Ecological Data Mining
4.4. Germination Tests
4.5. Seedling Growth
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xing, G.; Qu, L.; Zhang, Y.; Xue, L.; Su, J.; Lei, J. Collection and evaluation of wild tulip (Tulipa spp.) resources in China. Genet. Resour. Crop. Evol. 2017, 64, 641–652. [Google Scholar] [CrossRef]
- Zhang, W.; Zhao, J.; Xue, L.; Dai, H.; Lei, J. Seed morphology and germination of native Tulipa species. Agriculture 2023, 13, 466. [Google Scholar] [CrossRef]
- Krigas, N.; Lykas, C.; Ipsilantis, I.; Matsi, T.; Weststrand, S.; Havström, M.; Tsoktouridis, G. Greek tulips: Worldwide electronic trade over the internet, global ex situ conservation and current sustainable exploitation challenges. Plants 2021, 10, 580. [Google Scholar] [CrossRef] [PubMed]
- Van Bragt, J.; van Ast, K.J. Substitution of the cold requirement of tulip cv. ‘Apeldoorn’ by GA3. Sci. Horticult. 1976, 4, 117–122. [Google Scholar] [CrossRef]
- Rice, R.D.; Alderson, P.G.; Wright, N.A. Induction of bulbing of tulip shoots in vitro. Sci. Horticult. 1983, 20, 377–390. [Google Scholar] [CrossRef]
- Alderson, P.G.; Taeb, A.G. Effect of bulb storage on shoot regeneration from floral stems of tulip in vitro. Horticult. Sci. 1990, 65, 71–74. [Google Scholar] [CrossRef]
- Kuijpers, A.M.; Langens-Gerrits, M. Propagation of tulip in vitro. Acta Horticult. 1997, 430, 321–324. [Google Scholar] [CrossRef]
- Podwyszynska, M.; Marasek-Ciolakowska, A. Micropropagation of Tulip via somatic embryogenesis. Agronomy 2020, 10, 1857. [Google Scholar] [CrossRef]
- Boeken, B.; Gutterman, Y. The effect of temperature on seed germination in three common bulbous plants of different habitats in the Central Negev Desert of Israel. J. Arid. Environ. 1990, 18, 175–184. [Google Scholar] [CrossRef]
- Tang, A.J.; Tian, M.H.; Long, C.L. Seed dormancy and germination of three herbaceous perennial desert ephemerals from the Junggar Basin, China. Seed Sci. Res. 2009, 19, 183–189. [Google Scholar] [CrossRef]
- Zhang, W.; Qu, L.W.; Zhao, J.; Xue, L.; Dai, H.P.; Xing, G.M.; Lei, J.J. Practical methods for breaking seed dormancy in a wild ornamental tulip species Tulipa thianschanica Regel. Agronomy 2020, 10, 1765. [Google Scholar] [CrossRef]
- Rouhi, H.R.; Afshari, R.T.; Shakarami, K. Seed Treatments to Overcome Dormancy of Waterlily Tulip (Tulipa kaufmanniana Regel.). Aus. J. Crop. Sci. 2010, 4, 718–721. Available online: http://www.cropj.com/tavakol_4_9_2010_718_721.pdf (accessed on 6 March 2023).
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed germination and vigor. Ann. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef] [Green Version]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination, 2nd ed.; Academic Press: San Diego, CA, USA, 2014. [Google Scholar]
- Song, D.; Zhou, J.; Lai, L.; Alarcon, I.; Tar’An, B.; Abrams, S. Development of ABA antagonists to overcome ABA- and low temperature-induced inhibition of seed germination in canola, lentil, and soybean. J. Plant Growth Regul. 2019, 39, 1403–1413. [Google Scholar] [CrossRef]
- Fenner, M.; Thompson, K. The Ecology of Seeds; Cambridge University Press: Cambridge, UK, 2005; p. 250. [Google Scholar]
- Bell, D.T. The process of germination in Australian species. Aus. J. Bot. 1999, 47, 475–517. [Google Scholar] [CrossRef]
- Perez-García, F.; Iriondo, J.M.; Gonzalez-Benito, M.E.; Carnes, L.F.; Tapia, J.; Prieto, C.; Plaza, R.; Perez, C. Germination studies in endemic plant species of the Iberian Peninsula. Isr. J. Plant Sci. 1995, 43, 239–247. [Google Scholar] [CrossRef]
- Doussi, M.A.; Thanos, C.A. Ecophysiology of seed germination in Mediterranean geophytes. 1. Muscari spp. Seed Sci. Res. 2002, 12, 193–201. [Google Scholar] [CrossRef]
- Kadis, C.; Kounnamas, C.; Georghiou, K. Seed germination and conservation of endemic, rare and threatened aromatic plants of Cyprus. Isr. J. Plant Sci. 2010, 58, 251–261. [Google Scholar] [CrossRef]
- Krigas, N.; Mouflis, G.; Grigoriadou, K.; Maloupa, E. Conservation of important plants from the Ionian Islands at the Balkan Botanic Garden of Kroussia, N Greece: Using GIS to link the in situ collection data with plant propagation and ex situ cultivation. Biodivers Conserv. 2010, 19, 3583–3603. [Google Scholar] [CrossRef]
- Krigas, N.; Papadimitriou, K.; Mazaris, A.D. GIS and Ex-Situ Plant Conservation. In Application of Geographic Information Systems; Alam, B.M., Ed.; IntechOpen: Rijeka, Croatia, 2012. [Google Scholar] [CrossRef] [Green Version]
- Hatzilazarou, S.; El Haissoufi, M.; Pipinis, E.; Kostas, S.; Libiad, M.; Khabbach, A.; Lamchouri, F.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; et al. GIS-facilitated seed germination and multifaceted evaluation of the endangered Abies marocana Trab. (Pinaceae): Enabling conservation and sustainable exploitation. Plants 2021, 10, 2606. [Google Scholar] [CrossRef]
- Kostas, S.; Hatzilazarou, S.; Pipinis, E.; Bourgou, S.; Ben Haj Jilani, I.; Ben Othman, W.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Libiad, M.; Khabbach, A.; et al. DNA Barcoding, GIS-facilitated seed germination and pilot cultivation of Teucrium luteum subsp. gabesianum (Lamiaceae), a Tunisian local endemic with potential medicinal and ornamental value. Biology 2022, 11, 462. [Google Scholar] [CrossRef]
- Sarrou, E.; Doukidou, L.; Avramidou, E.V.; Martens, S.; Angeli, A.; Stagiopoulou, R.; Fyllas, N.M.; Tourvas, N.; Abraham, E.; Maloupa, E.; et al. Chemodiversity is closely linked to genetic and environmental diversity: Insights into the Endangered populations of the local endemic plant Sideritis euboea Heldr. of Evia Island (Greece). J. Appl. Res. Med. Arom. Plants 2022, 31, 100426. [Google Scholar] [CrossRef]
- Tsiftsoglou, O.S.; Stagiopoulou, R.; Krigas, N.; Lazari, D. Exploring the ecological preferences and essential oil variability in wild-growing populations of the endangered local Greek endemic Thymus holosericeus (Lamiaceae). Plants 2023, 12, 348. [Google Scholar] [CrossRef]
- Grigoriadou, K.; Sarropoulou, V.; Krigas, N.; Maloupa, E.; Tsoktouridis, G. GIS-facilitated effective propagation protocols of the Endangered local endemic of Crete Carlina diae (Rech. f.) Meusel and A. Kástner (Asteraceae): Serving ex-situ conservation needs and its future sustainable exploitation as an ornamental. Plants 2020, 9, 1465. [Google Scholar] [CrossRef] [PubMed]
- Kougioumoutzis, K.; Kokkoris, I.P.; Panitsa, M.; Strid, A.; Dimopoulos, P. Extinction risk assessment of the Greek endemic flora. Biology 2021, 10, 195. [Google Scholar] [CrossRef] [PubMed]
- Thanos, C.A.; Kadis, C.C.; Skarou, F. Ecophysiology of germination in the aromatic plants thyme, savory and oregano (Labiatae). Seed Sci. Res. 1995, 5, 161–170. [Google Scholar] [CrossRef]
- Krichen, K.; Mariem, H.B.; Chaieb, M. Ecophysiological requirements on seed germination of a Mediterranean perennial grass (Stipa tenacissima L.) under controlled temperatures and water stress. S. Afr. J. Bot. 2014, 94, 210–217. [Google Scholar] [CrossRef] [Green Version]
- Nikolaeva, M.G. Factors controlling the seed dormancy pattern. In The Physiology and Biochemistry of Seed Dormancy and Germination; Kahn, A.A., Ed.; North-Holland: Amsterdam, The Netherlands, 1977; pp. 51–74. [Google Scholar]
- Iwaya-Inoue, M.; Motooka, K.; Watanabe, G. Significance of Low Temperature Requirement on Dormancy Breaking for Tulip Bulbs as Estimated by 1H-NMR. Cryobiol. Cryotechn. 1998, 44, 21–28. Available online: https://ndlonline.ndl.go.jp/#!/detail/R300000002-I4549248-00 (accessed on 6 March 2023).
- Van Tuyl, J.M.; van Creij, M.G.M. TULIP. Tulipa gesneriana and Tulipa hybrids. In Flower Breeding and Genetics; Anderson, N.O., Ed.; Springer: Wageningen, The Netherlands, 2007; pp. 623–641. [Google Scholar]
- Strid, A. Atlas of the Aegean Flora, Part 1: Text & Plates; Part 2: Maps; Englera, Volume 33; Botanic Garden and Botanical Museum Berlin, Freie Universität Berlin: Berlin, Germany, 2016; ISBN 978-3-921800-97-3 (Volume 1). 978-3-921800-98-0 (Volume 2). [Google Scholar]
- Hijmans, R.J.; van Etten, J. Raster: Geographic Analysis and Modeling with Raster Data. R package Version 2.1-66. 2010. Available online: https://www.researchgate.net/publication/308748932_Raster_geographic_analysis_and_modeling_with_raster_data_R_package_version_21-66 (accessed on 6 February 2023).
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Intern. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Pipinis, E.; Hatzilazarou, S.; Kostas, S.; Bourgou, S.; Megdiche-Ksouri, W.; Ghrabi-Gammar, Z.; Libiad, M.; Khabbach, A.; El Haissoufi, M.; Lamchouri, F.; et al. Facilitating conservation and bridging gaps for the sustainable exploitation of the Tunisian local endemic Plant Marrubium Aschersonii (Lamiaceae). Sustainability 2022, 14, 1637. [Google Scholar] [CrossRef]
- ISTA (International Seed Testing Association) International Rules for Seed Testing. Seed Sci. Technol. 1999, 27, 333.
- Klockars, A.; Sax, G. Multiple Comparisons; Sage Publications: Newbury Park, CA, USA, 1986; p. 87. [Google Scholar]
- Snedecor, G.W.; Cochran, W.C. Statistical Methods, 7th ed.; The Iowa State University Press: Ames, IA, USA, 1980; p. 507. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Bulblet Weight (mg) | Bulblet Length (cm) | Bulblet Width (cm) |
|---|---|---|---|
| Tulipa australis | 25.26 ± 3.76 a | 0.2455 ± 0.021 a | 0.4909 ± 0.045 ab |
| Tulipa bakeri | 11.82 ± 2.50 b | 0.1667 ± 0.022 b | 0.4333 ± 0.028 bc |
| Tulipa clusiana | 15.19 ± 2.31 b | 0.1909 ± 0.021 b | 0.3545 ± 0.031 d |
| Tulipa goulimyi | 14.04 ± 1.72 b | 0.1947 ± 0.009 b | 0.3842 ± 0.016 cd |
| Tulipa undulatifolia | 23.76 ± 2.70 a | 0.2583 ± 0.019 a | 0.5083 ± 0.023 a |
| Tulipa spp. | Annual Temperature (°C) | Lowest Annual Temperature (°C) | Highest Annual Temperature (°C) |
| T. australis | 10.43 ± 3.37 | 4.22 | 17.80 |
| T. bakeri | 12.72 ± 0.18 | 13.35 | 12.85 |
| T. clusiana | 15.75 ± 0.71 | 15.25 | 16.25 |
| T. goulimyi | 15.73 ± 1.01 | 14.10 | 17.72 |
| T. undulatifolia | 15.26 ± 2.30 | 8.35 | 17.94 |
| Tulipa spp. | Annual Precipitation (mm) | Lowest Annual Precipitation (mm) | Highest Annual Precipitation (mm) |
| T. australis | 740.15 ± 163.08 | 455 | 1026 |
| T. bakeri | 859.78 ± 7.40 | 852 | 873 |
| T. clusiana | 585 ± 25.46 | 567 | 603 |
| T. goulimyi | 611.80 ± 34.27 | 568 | 666 |
| T. undulatifolia | 598 ± 99.43 | 506 | 905 |
| Scientific Name/Family | IPEN Accession | Altitude (m) | Collection Site and Area | Latitude (North) | Longitude (East) |
|---|---|---|---|---|---|
| Tulipa australis | GR-BBGK-1- 21,278 | 1614 | Mt. Chelmos, Peloponnese | 38.01447 | 22.19009 |
| Tulipa bakeri | GR-BBGK-1- 20,414 | 1200 | Omalos, Chania | 35.33333 | 23.9000 |
| Tulipa clusiana | GR-BBGK-1- 21,277 | 77 | Agios Ioannis Prodromos, Chios Island | 38.33788 | 26.09824 |
| Tulipa goulimyi | GR-BBGK-1- 20,149 | 612 | Molai, Lakonia | 36.82899 | 22.94770 |
| Tulipa undulatifolia | GR-BBGK-1- 20,144 | 152 | Didyma, Argolida | 37.45261 | 23.17304 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hatzilazarou, S.; Pipinis, E.; Kostas, S.; Stagiopoulou, R.; Gitsa, K.; Dariotis, E.; Avramakis, M.; Samartza, I.; Plastiras, I.; Kriemadi, E.; et al. Influence of Temperature on Seed Germination of Five Wild-Growing Tulipa Species of Greece Associated with Their Ecological Profiles: Implications for Conservation and Cultivation. Plants 2023, 12, 1574. https://doi.org/10.3390/plants12071574
Hatzilazarou S, Pipinis E, Kostas S, Stagiopoulou R, Gitsa K, Dariotis E, Avramakis M, Samartza I, Plastiras I, Kriemadi E, et al. Influence of Temperature on Seed Germination of Five Wild-Growing Tulipa Species of Greece Associated with Their Ecological Profiles: Implications for Conservation and Cultivation. Plants. 2023; 12(7):1574. https://doi.org/10.3390/plants12071574
Chicago/Turabian StyleHatzilazarou, Stefanos, Elias Pipinis, Stefanos Kostas, Rafaela Stagiopoulou, Konstantina Gitsa, Eleftherios Dariotis, Manolis Avramakis, Ioulietta Samartza, Ioannis Plastiras, Eleni Kriemadi, and et al. 2023. "Influence of Temperature on Seed Germination of Five Wild-Growing Tulipa Species of Greece Associated with Their Ecological Profiles: Implications for Conservation and Cultivation" Plants 12, no. 7: 1574. https://doi.org/10.3390/plants12071574