
Plant Genotype Shapes the Soil Nematode Community in the Rhizosphere of Tomatoes with Different Resistance to Meloidognye incognita


Abstract
:1. Introduction
2. Results
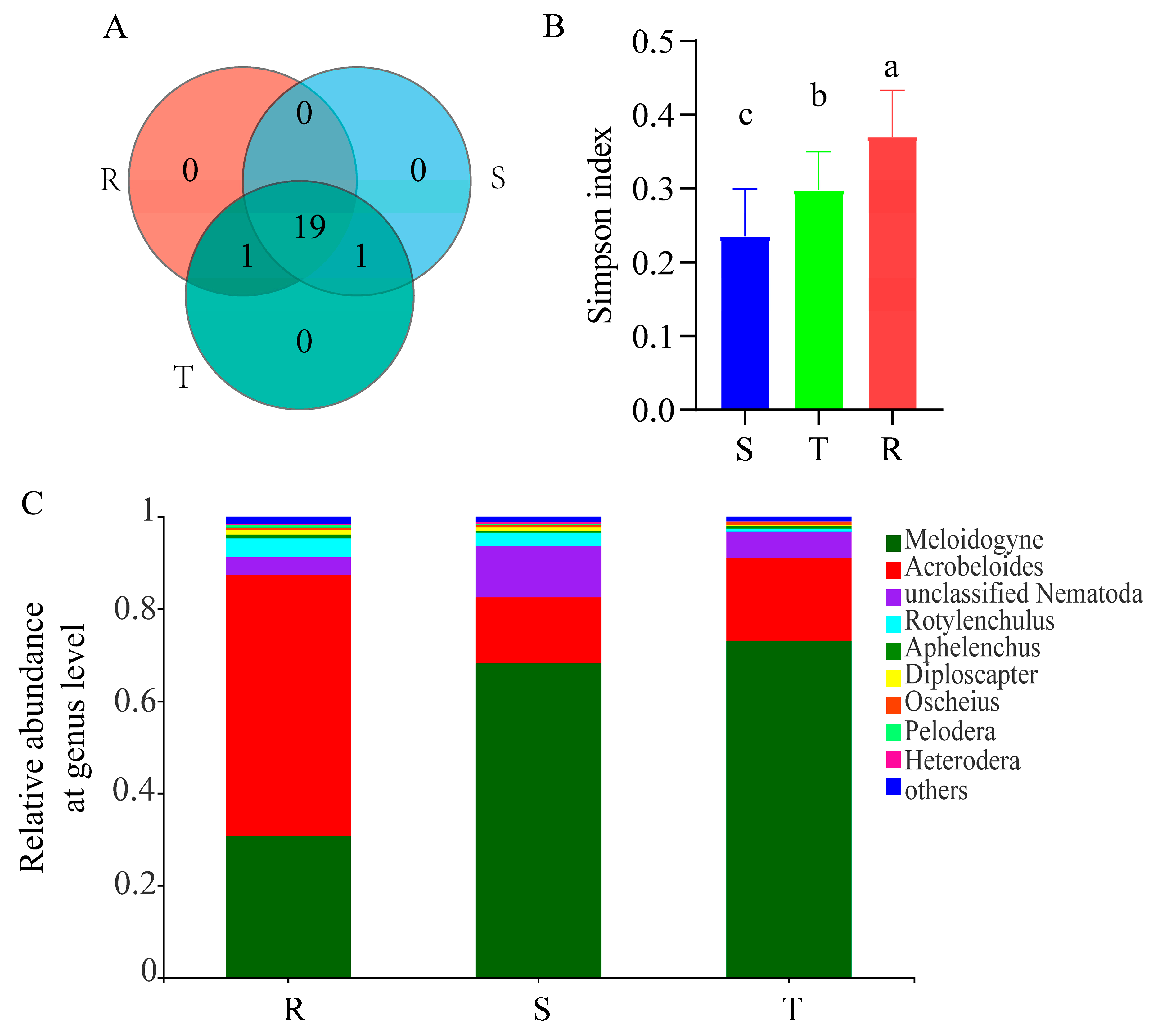
2.1. Alpha Diversity Analysis
2.2. Beta Diversity Analysis
2.3. Comparison of Ecological Indices of Three Soil Nematode Communities and Disease Index
2.4. Effect of Root Exudates on Soil Nematode Community
2.5. Effects of Root Exudates on M. incognita
3. Discussion
4. Materials and Methods
4.1. Tomato and M. incognita Materials
4.2. Collection of Root Exudates and Gas Chromatography/Mass Spectrometry Analysis
4.3. Tomato Planting in the Greenhouse
4.4. Soil Nematode Isolation and DNA Extraction
4.5. PCR Amplification and Pyrosequencing
4.6. Bioinformation Analysis
4.7. Ecological Index Analysis
4.8. Effect of Root Exudates on Mortality Rate of M. incognita J2
4.9. Effect of Root Exudates on M. incognita Egg-Hatching
4.10. Effect of Root Exudates on the Chemotaxis of M. incognita J2
4.11. Data Analysis
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tian, X.L.; Zhao, X.M.; Mao, Z.C.; Xie, B.Y. Variation and dynamics of soil nematode communities in greenhouses with different continuous cropping periods. Hortic. Plant J. 2020, 6, 301–312. [Google Scholar] [CrossRef]
- Grabau, Z.J.; Liu, C.; Sandoval-Ruiz, R. Meloidogyne incognita management by nematicides in tomato production. J. Nematol. 2021, 53, e2021–e2055. [Google Scholar] [CrossRef]
- Jacquet, M.; Bongiovanni, M.; Martinez, M.; Verschave, P.; Wajnberg, E.; Castagnone-Sereno, P. Variation in resistance to the root-knot nematode Meloidogyne incognita in tomato genotypes bearing the Mi gene. Plant Pathol. 2005, 54, 93–99. [Google Scholar] [CrossRef]
- Rick, C.M. Hybrids between Lycopersicon esculentum Mill and Solanum Lycopersicoides Dun. Proc. Natl. Acad. Sci. USA 1951, 37, 741–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cortada, L.; Sorribas, F.J.; Ornat, C.; Andrés, M.F.; Verdejo-Lucas, S. Response of tomato rootstocks carrying the Mi-resistance gene to populations of Meloidogyne arenaria, M. incognita and M. javanica. Eur. J. Plant Pathol. 2009, 124, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Ferris, H.; Bongers, T.; De Goede, R.G.M. Nematode faunal indicators of soil food web condition. J. Nematol. 1999, 31, 63–67. [Google Scholar]
- Hu, J.; Wu, J.; Ma, M.; Nielsen, U.N.; Wang, J.; Du, G. Nematode communities response to long-term grazing disturbance on Tibetan plateau. Eur. J. Soil Biol. 2015, 69, 24–32. [Google Scholar] [CrossRef]
- Zhao, J.; Neher, D.A. Soil nematode genera that predict specific types of disturbance. Appl. Soil Ecol. 2013, 64, 135–141. [Google Scholar] [CrossRef]
- Yeates, G.W. Abundance, diversity, and resilience of nematode assemblages in forest soils. Can. J. For. Res. 2007, 37, 216–225. [Google Scholar] [CrossRef]
- Bell, C.A.; Namaganda, J.; Urwin, P.E.; Atkinson, H.J. Next-generation sequencing of the soil nematode community enables the sustainability of banana plantations to be monitored. Appl. Soil Ecol. 2021, 166, 103999. [Google Scholar] [CrossRef]
- Davey, M.L.; Utaaker, K.S.; Fossøy, F. Characterizing parasitic nematode faunas in faeces and soil using DNA metabarcoding. Parasit. Vectors 2021, 14, 422. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interaction with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, B.S. The role of bacterial feeding nematodes and protozoa in rhizosphere nutrient cycling. Asp. Appl. Biol. 1989, 22, 141–145. [Google Scholar]
- Rocha, R.O.; Morais, J.K.; Oliveira, J.T.; Oliveira, H.D.; Sousa, D.O.; Souza, C.E.; Moreno, F.; Monteiro-Moreira, A.C.; de Souza Júnior, J.D.; de Sá, M.F.; et al. Proteome of soybean seed exudates contains plant defense-related proteins active against the root-knot nematode Meloidogyne incognita. J. Agric. Food Chem. 2015, 63, 5335–5343. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Masler, E.P.; Rogers, S.T. Responses of Heterodera glycines and Meloidogyne incognita infective juveniles to root tissues, root exudates, and root extracts from three plant species. Plant Dis. 2018, 102, 1733–1740. [Google Scholar] [CrossRef] [Green Version]
- Wuyts, N.; Swennen, R.; De Waele, D. Effects of plant phenylpropanoid pathway products and selected terpenoids and alkaloids on the behaviour of the plant-parasitic nematodes Radopholus similis, Pratylenchus penetrans and Meloidogyne incognita. Nematology 2006, 8, 89–101. [Google Scholar] [CrossRef]
- Sikder, M.M.; Vestergård, M.; Kyndt, T.; Fomsgaard, I.S.; Kudjordjie, E.N.; Nicolaisen, M. Benzoxazinoids selectively affect maize root-associated nematode taxa. J. Exp. Bot. 2021, 72, 3835–3845. [Google Scholar] [CrossRef]
- Vaid, S.; Shah, A.; Ahmad, R.; Hussain, A. Diversity of soil inhabiting nematodes in Dera Ki Gali forest of Poonch district, Jammuand Kashmir, India. Int. J. Nematol. 2014, 24, 97–102. [Google Scholar]
- Yeates, G.W.; Newton, P.C. Long-term changes in topsoil nematode populations in grazed pasture under elevated atmospheric carbon dioxide. Biol. Fertil. Soils 2009, 45, 799–808. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, D.; Li, H.; Qi, X.; Gao, Y.; Zhang, Y.; Han, Y.; Jiang, Y.; Li, H. Soil nematode community and crop productivity in response to 5-year biochar and manure addition to yellow cinnamon soil. BMC Ecol. 2020, 20, 39. [Google Scholar] [CrossRef] [PubMed]
- Bulluck, L.R.; Ristaino, J.B. Effect of synthetic and organic soil fertility amendments on southern blight, soil microbial communities, and yield of processing tomatoes. Phytopathology 2002, 92, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Zhou, D.; Feng, H.; Schuelke, T.; De Santiago, A.; Zhang, Q.; Zhang, J.; Luo, C.; Wei, L. Rhizosphere microbiomes from root knot nematode non-infested plants suppress nematode infection. Microb. Ecol. 2019, 78, 470–481. [Google Scholar] [CrossRef] [Green Version]
- Korthals, G.W.; Smilauer, P.; Van, D.C.; Van, W.H. Linking above and below-ground biodiversity: Abundance and trophic complexity in soil as a response to experimental plant communities on abandoned arable land. Funct. Ecol. 2001, 15, 506–514. [Google Scholar] [CrossRef]
- Quist, C.W.; Schrama, M.; de Haan, J.J.; Smant, G.; Bakker, J.; van der Putten, W.H.; Helder, J. Organic farming practices result in compositional shifts in nematode communities that exceed crop-related changes. Appl. Soil. Ecol. 2016, 98, 254–260. [Google Scholar] [CrossRef]
- Yan, J.; Zhang, Y.; Crawford, K.M.; Chen, X.; Yu, S.; Wu, J. Plant genotypic diversity effects on soil nematodes vary with trophic level. New Phytol. 2021, 229, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Palomares-Rius, J.E.; Castillo, P.; Montes-Borrego, M.; Navas-Cortés, J.A.; Landa, B.B. Soil properties and olive cultivar determine the structure and diversity of plant-parasitic nematode communities infesting olive orchards soils in southern Spain. PLoS ONE 2015, 10, e0116890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dries, L.; Bussotti, S.; Pozzi, C.; Kunz, R.; Schnell, S.; Löhnertz, O.; Vortkamp, A. Rootstocks shape their microbiome-bacterial communities in the rhizosphere of different grapevine rootstocks. Microorganisms 2021, 9, 822. [Google Scholar] [CrossRef]
- García-Sánchez, M.; Souche, M.; Trives-Segura, C.; Plassard, C. The grazing activity of Acrobeloides sp. drives phytate mineralisation within its trophic relationship with bacteria. J. Nematol. 2021, 53, e2021. [Google Scholar] [CrossRef]
- Faske, T.R.; Kandel, Y.; Allen, T.W.; Grabau, Z.J.; Hu, J.; Kemerait, R.C.; Lawrence, G.W.; Lawrence, K.S.; Mehl, H.L.; Overstreet, C.; et al. Meta-analysis of the field efficacy of seed- and soil-applied nematicides on Meloidogyne incognita and Rotylenchulus reniformis across the U.S. cotton belt. Plant Dis. 2022, 106, 2228–2238. [Google Scholar] [CrossRef]
- Liu, Y.; Hua, J.; Jiang, Y.; Li, Q.; Wen, D. Nematode communities in greenhouse soil of different ages from Shenyang suburb. Helminthologia 2006, 43, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Bongers, T.; Meulen, H.; Korthals, G. Inverse relationship between the nematode maturity index and plant parasite index under enriched nutrient conditions. Appl. Soil Ecol. 1997, 6, 195–199. [Google Scholar] [CrossRef]
- Wasilewska, L. The effect of age of meadows on succession and diversity in soil nematode communities. Pedobiologia 1994, 38, 1–11. [Google Scholar]
- Vives-Peris, V.; de Ollas, C.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Root exudates: From plant to rhizosphere and beyond. Plant Cell Rep. 2020, 39, 3–17. [Google Scholar] [CrossRef]
- Chitwood, D.J. Phytochemical-based strategies for nematode control. Annu. Rev. Phytopathol. 2002, 40, 221–249. [Google Scholar] [CrossRef] [Green Version]
- Bonkowski, M.; Villenave, C.; Griths, B. Rhizosphere fauna: The functional and structural diversity of intimate interactions of soil fauna with plant roots. Plant Soil 2009, 321, 213–233. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moenne-Loccoz, Y. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Zhou, B.; Zhang, X.; Zhang, Z.; Wu, Y.; Zhang, Y.; Lü, S.; Zou, Q.; Gao, Y.; Teng, L. Effects of tomato root exudates on Meloidogyne incognita. PLoS ONE 2016, 11, e0154675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirwa, H.K.; Murungi, L.K.; Beck, J.J.; Torto, B. Elicitation of differential responses in the root-knot nematode Meloidogyne incognita to tomato root exudate Cytokinin, Flavonoids, and Alkaloids. J. Agric. Food Chem. 2018, 66, 11291–11300. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.H.; Zhou, B.L.; Zhang, F.L.; Zhang, Q. Allelopathic effects of root extracts of egg plants on verticillium wilt (Verticillium dahliae). Allelopath. J. 2005, 15, 75–83. [Google Scholar]
- Dube, G.C.; Smart, J. Biological control of Meloidogyne incognita by Paecilomyces lilacinus and Pasteuria penetrans. J. Nematol. 1987, 19, 222–227. [Google Scholar]
- Mishra, C.C.; Dash, M.C. Distribution and population dynamics of nematodes in a rice field and pasture in India. J. Nematol. 1981, 13, 538–543. [Google Scholar] [PubMed]
- Porazinska, D.L.; Giblin-Davis, R.M.; Faller, L.; Farmerie, W.; Kanzaki, N.; Morris, K.; Powers, T.O.; Tucker, A.E.; Sung, W.; Thomas, W.K. Evaluating high-throughput sequencing as a method for metagenomic analysis of nematode diversity. Mol. Ecol. Resour. 2009, 9, 1439–1450. [Google Scholar] [CrossRef] [PubMed]
- Darby, B.; Todd, T.C.; Herman, M.A. High-throughput amplicon sequencing of rRNA genes requires a copy number correction to accurately reflect the effects of management practices on soil nematode community structure. Mol. Ecol. 2013, 22, 5456–5471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Porazinska, D.L.; Giblin-Davis, R.M.; Powers, T.O.; Thomas, W.K. Nematode spatial and ecological patterns from tropical and temperate rainforests. PLoS ONE 2012, 7, e44641. [Google Scholar] [CrossRef] [Green Version]
- Morise, H.; Miyazaki, E.; Yoshimitsu, S.; Eki, T. Profiling nematode communities in unmanaged flowerbed and agricultural field soils in Japan by DNA barcode sequencing. PLoS ONE 2012, 7, e51785. [Google Scholar] [CrossRef]
- Rajilić-Stojanović, M.; Biagi, E.; Heilig, H.G.; Kajander, K.; Kekkonen, R.A.; Tims, S.; de Vos, W.M. Global and deep molecular analysis of microbiota signatures in fecal samples from patients with irritable bowel syndrome. Gastroenterology 2011, 14, 1792–1801. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; He, Z.; Tao, X.; Zhou, J.; Yang, Y.; Zhao, M.; Zhang, X.; Zheng, Z.; Tong, Y.; Liu, P.; et al. The shifts of sediment microbial community phylogenetic and functional structures during chromium (VI) reduction. Ecotoxicology 2016, 25, 1759–1770. [Google Scholar] [CrossRef] [Green Version]
- Segata, N.; Izard, J.; Waldron, L. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 2, R60. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.0-4. 2013. Available online: http://CRAN.R-project.org/package=vegan (accessed on 8 September 2021).
- Zaneveld, J.R.; Burkepile, D.E.; Shantz, A.A.; Pritchard, C.E.; McMinds, R.; Payet, J.P.; Welsh, R.; Correa, A.M.; Lemoine, N.P.; Rosales, S.; et al. Overfishing and nutrient pollution interact with temperature to disrupt coral reefs down to microbial scales. Nat. Commun. 2016, 7, 11833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bongers, T. The maturity index: An ecological measure of environmental disturbance based on nematode species composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Bongers, T.; Bongers, M. Functional diversity of nematodes. Appl. Soil Ecol. 1998, 10, 239–251. [Google Scholar] [CrossRef]
- Yeates, G.W. Nematodes as soil indicators: Functional and biodiversity aspects. Biol. Fert. Soils 2003, 37, 199–210. [Google Scholar] [CrossRef]
- Harada, Y.; Yoshiga, T. Distinguishing between inactivated and dead second stage juveniles of Meloidogyne incognita using the NaOH method. J. Nematol. 2016, 45, 51–55. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Root Exudates | Susceptible Cultivar | Tolerant Cultivar | Resistant Cultivar |
|---|---|---|---|
| Tetradecane | 7.28 ± 0.07 a | 6.85 ± 0.08 b | 8.86 ± 0.05 c |
| N-dodecane | 4.94 ± 0.05 a | 4.41 ± 0.04 b | 4.31 ± 0.05 c |
| 1,3-ditert-butylbenzene | 1.61 ± 0.02 a | 2.26 ± 0.02 b | 3.06 ± 0.06 c |
| Heptadecane | 3.06 ± 0.05 a | 3.63 ± 0.03 b | 5.85 ± 0.06 c |
| N-eicosane | 9.75 ± 0.04 a | 12.20 ± 0.09 b | 12.60 ± 0.03 c |
| 2,6-ditert-butylp-cresol | 16.44 ± 0.04 a | 16.18 ± 0.05 b | 14.94 ± 0.05 c |
| Palmitamide | 4.78 ± 0.04 a | 4.14 ± 0.03 b | 3.96 ± 0.02 c |
| Oleamide | 26.62 ± 0.14 a | 25.15 ± 0.05 b | 22.29 ± 0.08 c |
| 1-naphthyl aminobenzene | 1.27 ± 0.03 a | 1.47 ± 0.03 b | 1.50 ± 0.05 b |
| Stearamide | 4.05 ± 0.02 a | 4.27 ± 0.79 a | 3.73 ± 0.05 a |
| Undecane | 2.49 ± 0.05 a | 2.91 ± 0.03 b | 2.20 ± 0.04 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Wang, C.; Chen, R.; Wang, W.; Wang, D.; Tian, X. Plant Genotype Shapes the Soil Nematode Community in the Rhizosphere of Tomatoes with Different Resistance to Meloidognye incognita. Plants 2023, 12, 1528. https://doi.org/10.3390/plants12071528
Wang X, Wang C, Chen R, Wang W, Wang D, Tian X. Plant Genotype Shapes the Soil Nematode Community in the Rhizosphere of Tomatoes with Different Resistance to Meloidognye incognita. Plants. 2023; 12(7):1528. https://doi.org/10.3390/plants12071528
Chicago/Turabian StyleWang, Xiangmei, Chaoyan Wang, Ru Chen, Wenxing Wang, Diandong Wang, and Xueliang Tian. 2023. "Plant Genotype Shapes the Soil Nematode Community in the Rhizosphere of Tomatoes with Different Resistance to Meloidognye incognita" Plants 12, no. 7: 1528. https://doi.org/10.3390/plants12071528