
Allelopathic Potential of the Cyanotoxins Microcystin-LR and Cylindrospermopsin on Green Algae

 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Results
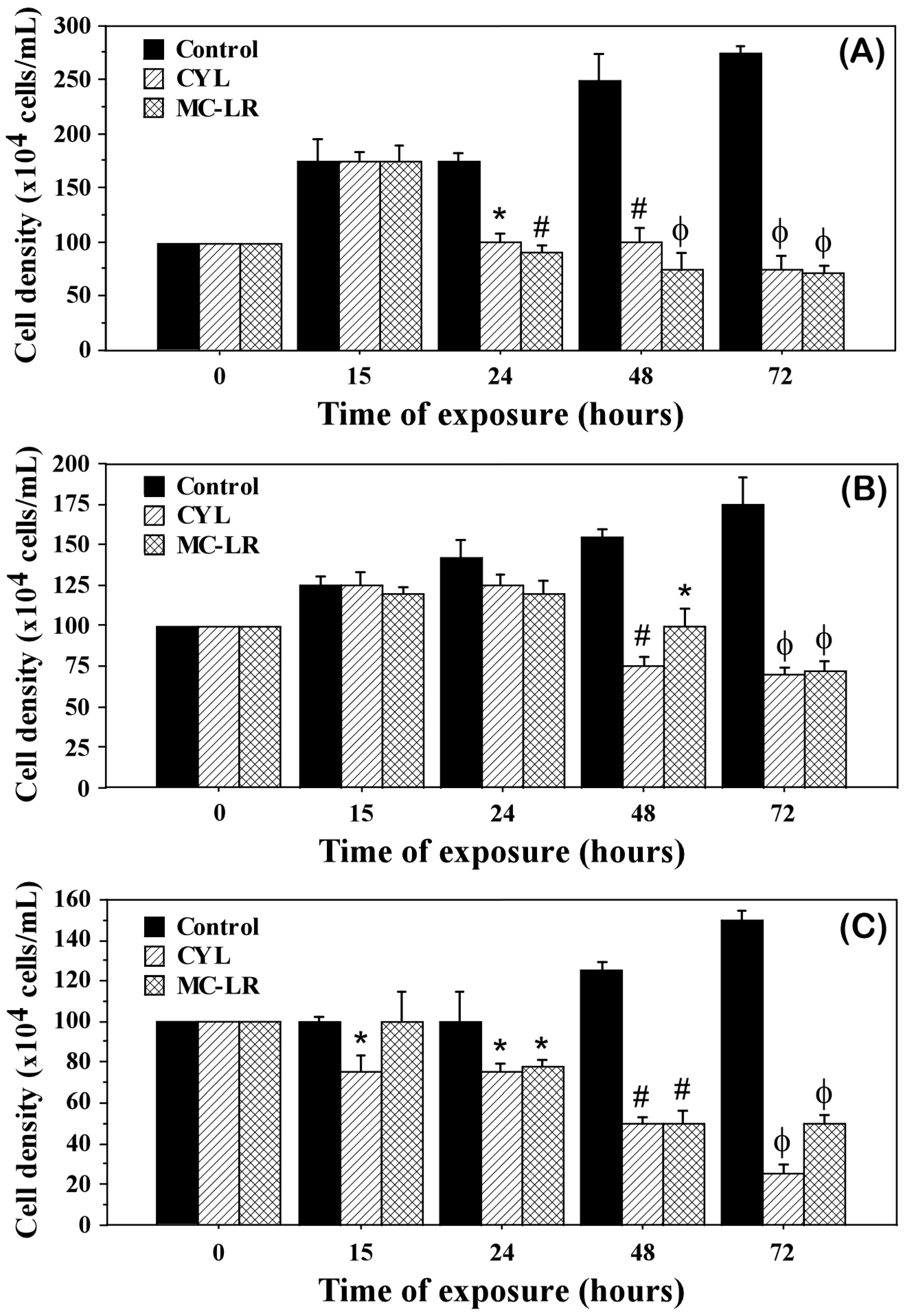
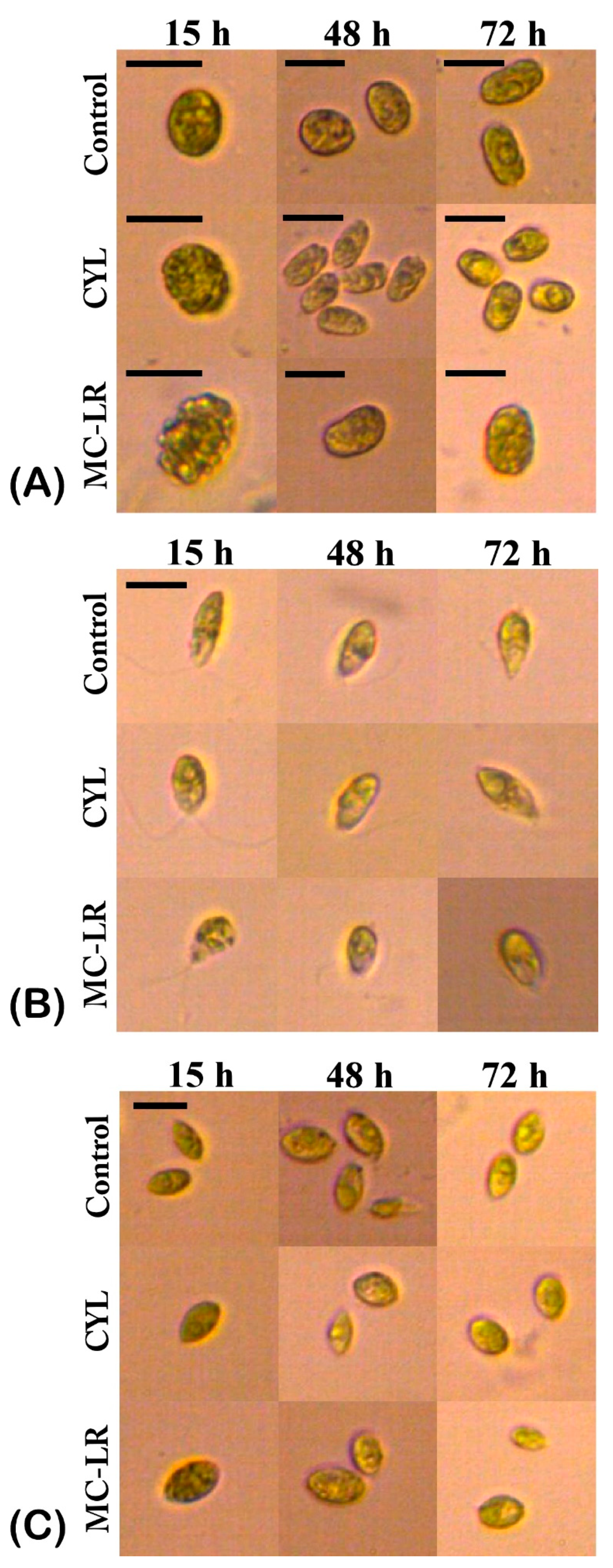
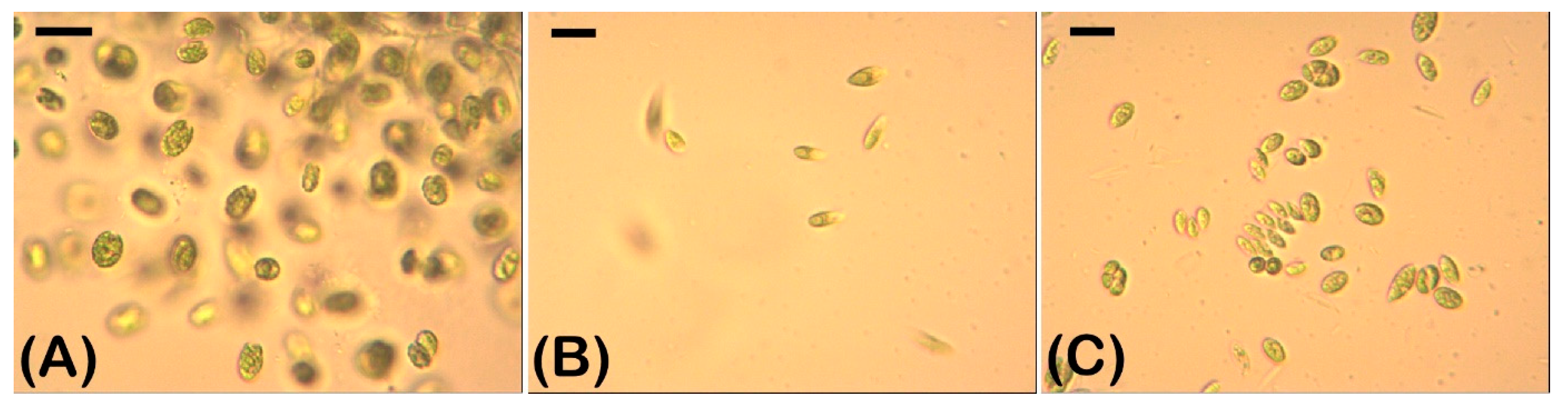
2.1. Density and Morphology of the Algae Exposed to Cyanotoxins
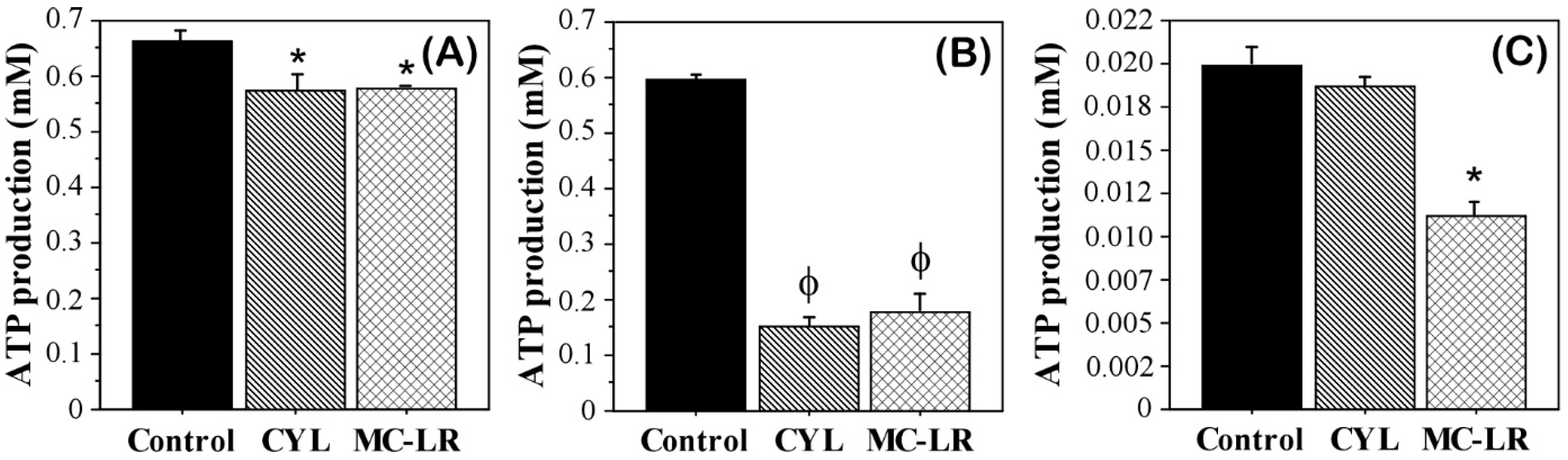
2.2. Intracellular ATP in Algae Exposed to Cyanotoxins
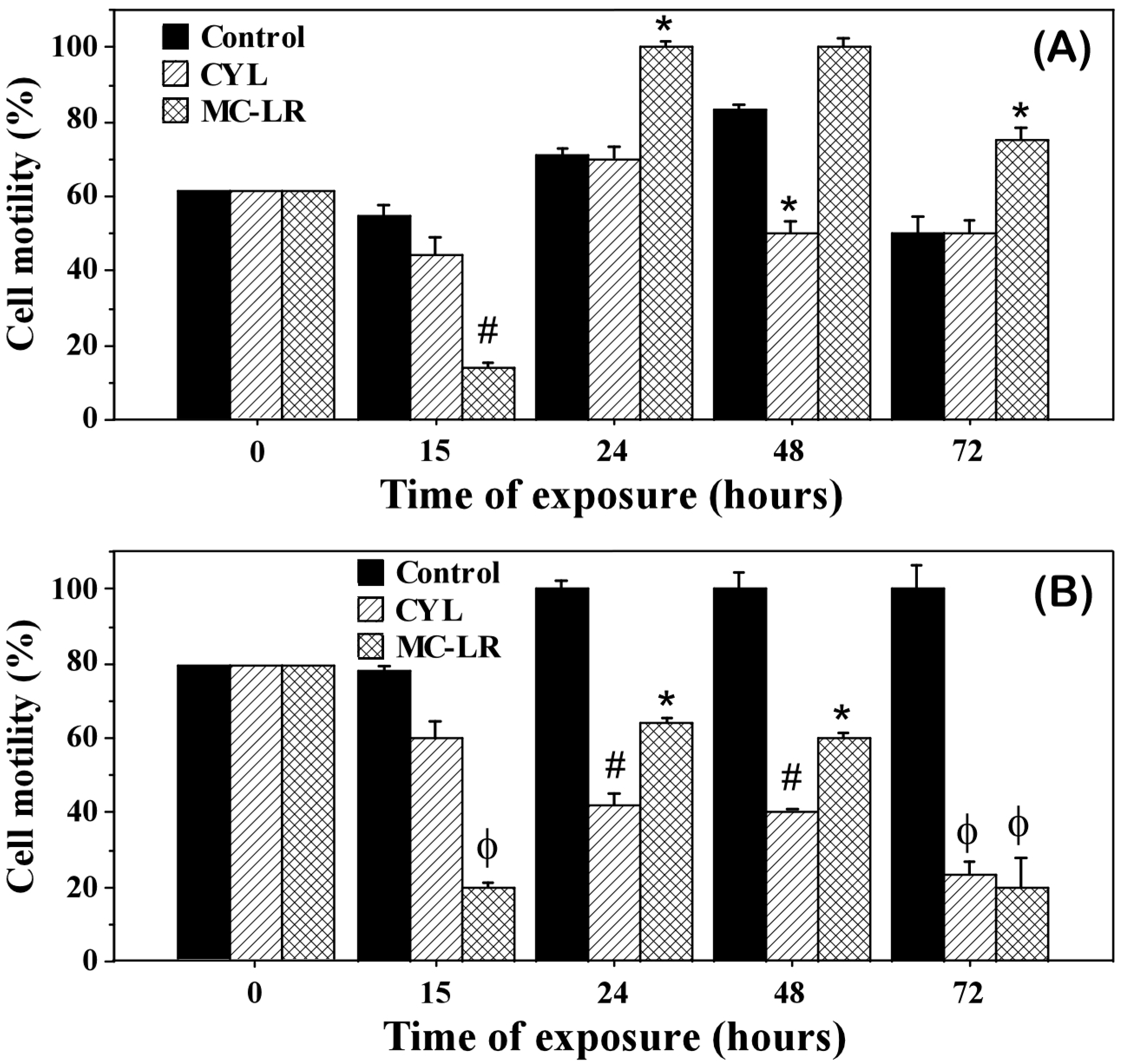
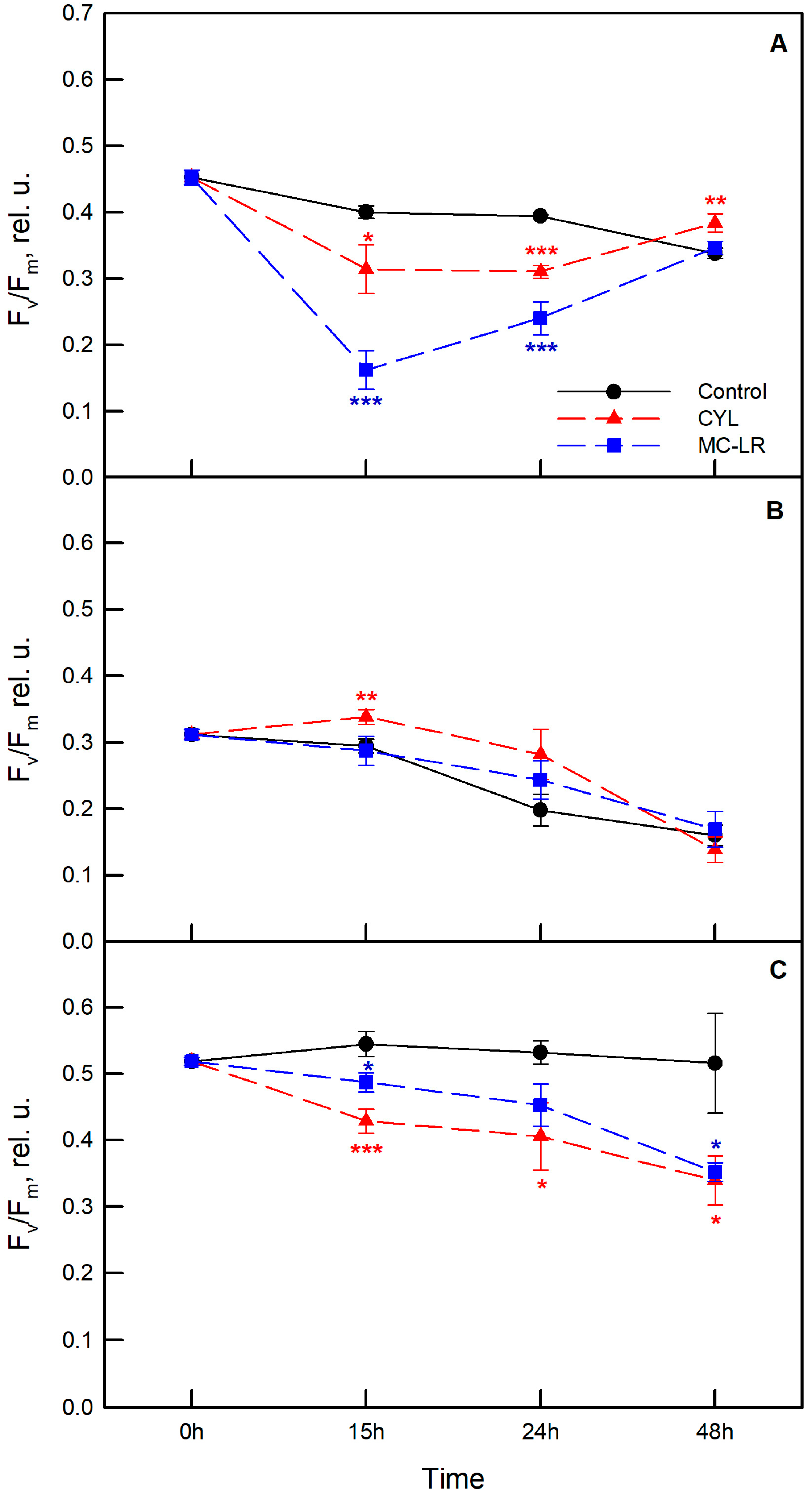
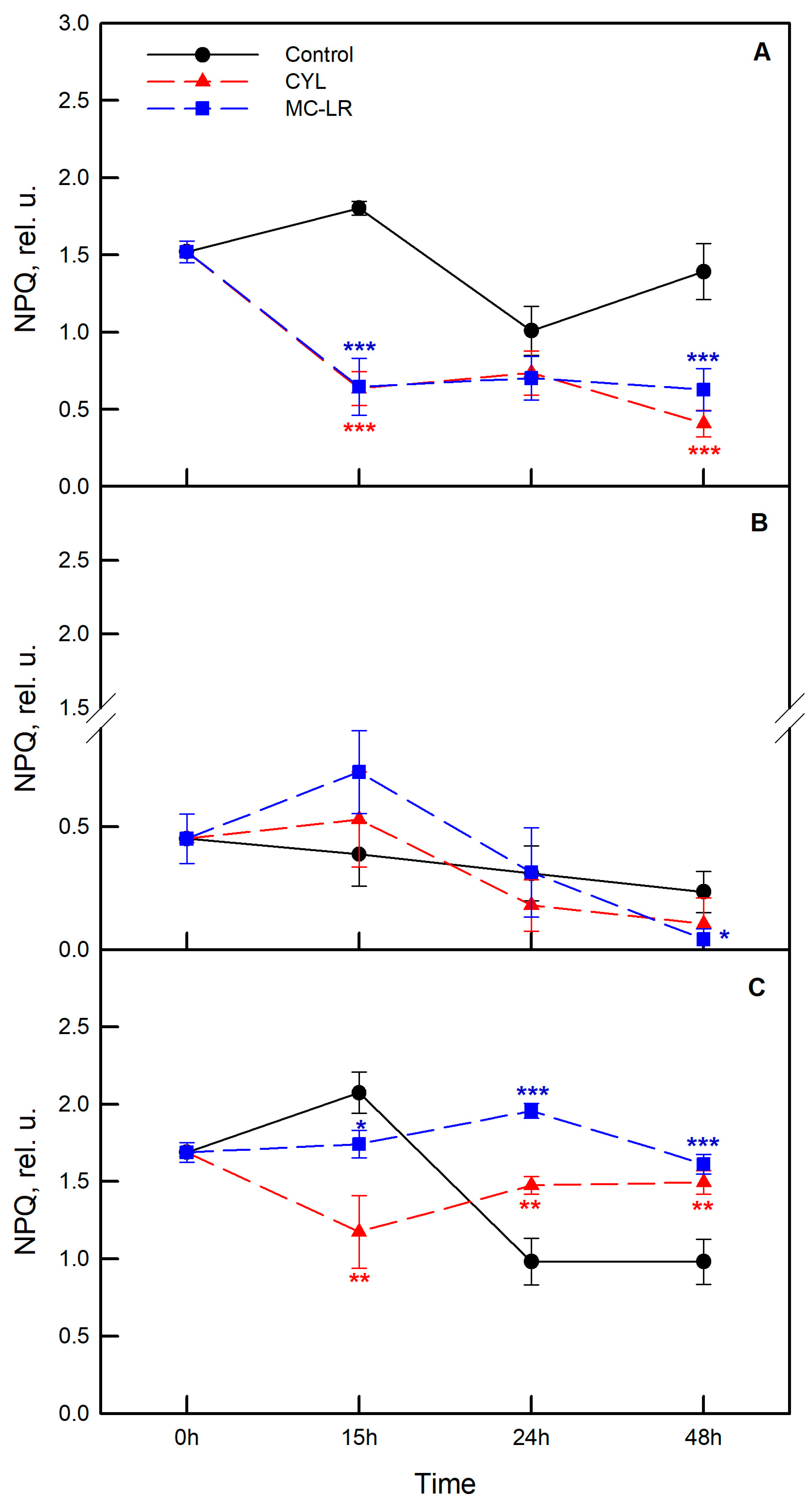
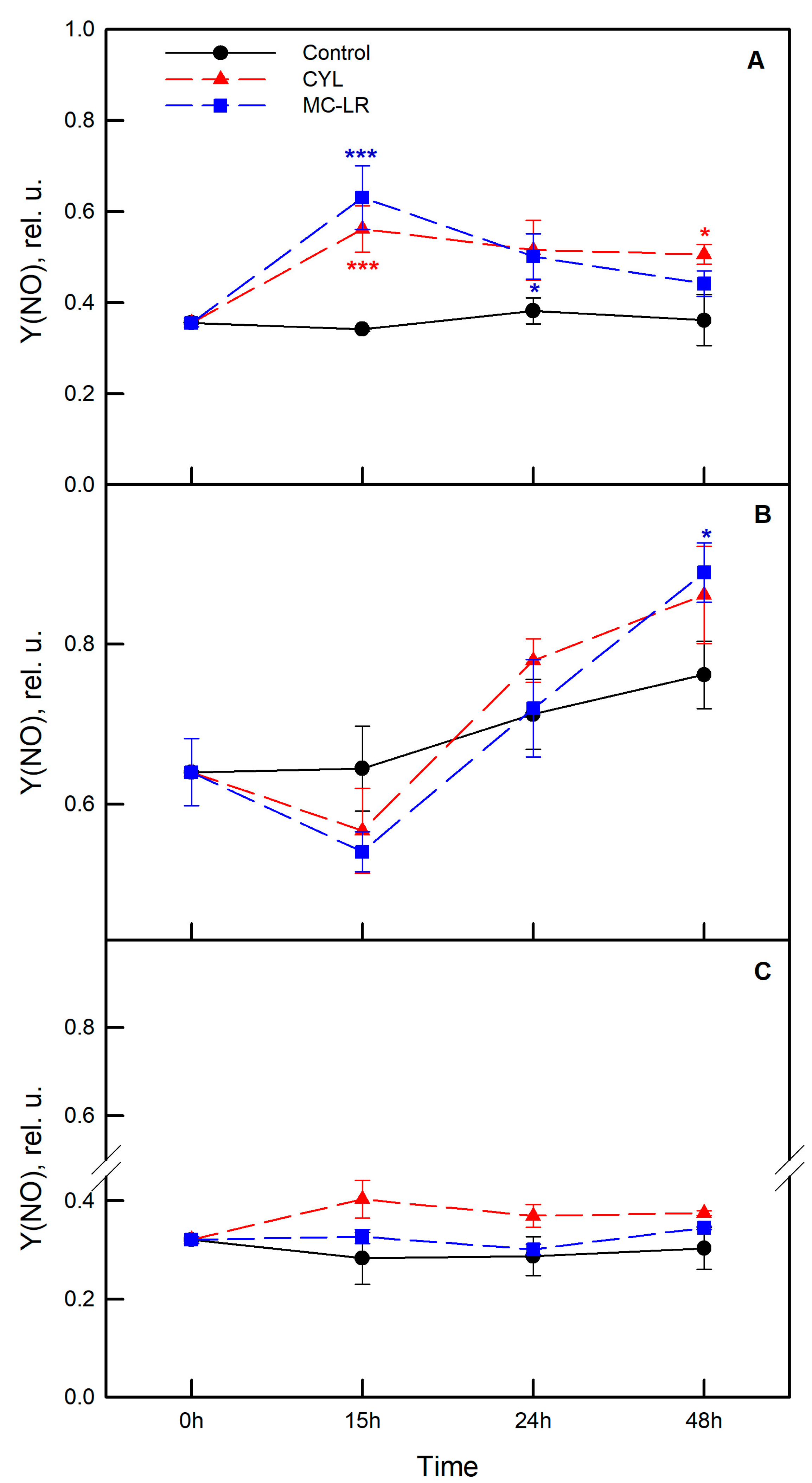
2.3. Photosynthetic Activity of the Algae Exposed to Cyanotoxins
3. Discussion
4. Materials and Methods
4.1. Algae, Culture Conditions and Exposure to Cyanotoxins
4.2. Counting of Cell Density and Morphological Analysis of the Algae
4.3. Measurement of Intracellular Adenosine Triphosphate (ATP) Concentration
4.4. Measurement of Photosynthetic Activity of the Algae
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water, 2nd ed.; CRC Press: Boca Raton, FL, USA; World Health Organization: Geneva, Switzerland, 2021; p. 859. [Google Scholar]
- Zhang, W.; Liu, J.; Xiao, Y.; Zhang, Y.; Yu, Y.; Zheng, Z.; Liu, Y.; Li, Q. The Impact of Cyanobacteria Blooms on the Aquatic Environment and Human Health. Toxins 2022, 14, 658. [Google Scholar] [CrossRef] [PubMed]
- Sazdova, I.; Keremidarska-Markova, M.; Chichova, M.; Uzunov, B.; Nikolaev, G.; Mladenov, M.; Schubert, R.; Stoyneva-Gärtner, M.; Gagov, H.S.; Hseu, Y.-C. Review of Cyanotoxicity Studies Based on Cell Cultures. J. Toxicol. 2022, 2022, 5647178. [Google Scholar] [CrossRef] [PubMed]
- Ahari, H.; Nowruzi, B.; Anvar, A.A.; Porzani, S.J. The Toxicity Testing of Cyanobacterial Toxins In vivo and In vitro by Mouse Bioassay: A Review. Mini-Rev. Med. Chem. 2022, 22, 1131–1151. [Google Scholar] [CrossRef] [PubMed]
- Zepernick, B.N.; Wilhelm, S.W.; Bullerjahn, G.S.; Paerl, H.W. Climate change and the aquatic continuum: A cyanobacterial comeback story. Environ. Microbiol. Rep. 2022, 15, 3–12. [Google Scholar] [CrossRef]
- Suikkanen, S.; Fistarol, G.O.; Granéli, E. Allelopathic effects of the Baltic cyanobacteria Nodularia spumdigena, Aphanizomenon flos-aquae and Anabaena lemmermannii on algal monocultures. J. Exp. Mar. Biol. Ecol. 2004, 308, 85–101. [Google Scholar] [CrossRef]
- Rzymski, P.; Poniedziałek, B. In search of environmental role of cylindrospermopsin: A review on global distribution and ecology of its producers. Water Res. 2014, 66, 320–337. [Google Scholar] [CrossRef]
- B-Béres, V.; Vasas, G.; Dobronoki, D.; Gonda, S.; Nagy, S.; Bácsi, I. Effects of Cylindrospermopsin Producing Cyanobacterium and Its Crude Extracts on a Benthic Green Alga—Competition or Allelopathy? Mar. Drugs 2015, 13, 6703–6722. [Google Scholar] [CrossRef] [Green Version]
- Omidi, A.; Pflugmacher, S.; Kaplan, A.; Kim, Y.J.; Esterhuizen, M. Reviewing Interspecies Interactions as a Driving Force Affecting the Community Structure in Lakes via Cyanotoxins. Microorganisms 2021, 9, 1583. [Google Scholar] [CrossRef]
- Holland, A.; Kinnear, S. Interpreting the Possible Ecological Role(s) of Cyanotoxins: Compounds for Competitive Advantage and/or Physiological Aide? Mar. Drugs 2013, 11, 2239–2258. [Google Scholar] [CrossRef] [Green Version]
- Omidi, A.; Esterhuizen-Londt, M.; Pflugmacher, S. Still challenging: The ecological function of the cyanobacterial toxin microcystin—What we know so far. Toxin Rev. 2017, 37, 87–105. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S.; Kumari, N.; Häder, D.-P.; Sinha, R.P. Cyanobacterial Blooms and Their Implications in the Changing Environment. Adv. Environ. Eng. Res. 2022, 3, 1. [Google Scholar] [CrossRef]
- Bittencourt-Oliveira, M.d.C.; Chia, M.A.; de Oliveira, H.S.B.; Cordeiro Araújo, M.K.; Molica, R.J.R.; Dias, C.T.S. Allelopathic interactions between microcystin-producing and non-microcystin-producing cyanobacteria and green microalgae: Implications for microcystins production. J. Appl. Phycol. 2014, 27, 275–284. [Google Scholar] [CrossRef]
- Wiegand, C.; Peuthert, A.; Pflugmacher, S.; Carmeli, S. Effects of microcin SF608 and microcystin-LR, two cyanotobacterial compounds produced by Microcystis sp., on aquatic organisms. Environ. Toxicol. 2002, 17, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Berry, J.P. Cyanobacterial Toxins as Allelochemicals with Potential Applications as Algaecides, Herbicides and Insecticides. Mar. Drugs 2008, 6, 117–146. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, A.; Kaur, A. Allelopathic impact of Cyanobacteria on pathogenic fungi. Int. J. Pure Appl. Biosci. 2014, 2, 63–70. [Google Scholar]
- Śliwińska-Wilczewska, S.; Maculewicz, J.; Barreiro Felpeto, A.; Latała, A. Allelopathic and Bloom-Forming Picocyanobacteria in a Changing World. Toxins 2018, 10, 48. [Google Scholar] [CrossRef] [Green Version]
- Gross, E.M. Allelopathy of Aquatic Autotrophs. Crit. Rev. Plant Sci. 2003, 22, 313–339. [Google Scholar] [CrossRef] [Green Version]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.H.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Moreira, C.; Vasconcelos, V.; Antunes, A. Cyanobacterial Blooms: Current Knowledge and New Perspectives. Earth 2022, 3, 10. [Google Scholar] [CrossRef]
- Švercel, M. Negative allelopathy among Cyanobacteria. In Cyanobacteria: Ecology, Toxicology and Management; Ferrão-Filho, A.D.S., Ed.; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2013; pp. 27–46. [Google Scholar]
- Dahms, H.-U.; Ying, X.; Pfeiffer, C. Antifouling potential of cyanobacteria: A mini-review. Biofouling 2006, 22, 317–327. [Google Scholar] [CrossRef]
- Leão, P.N.; Vasconcelos, M.T.S.D.; Vasconcelos, V.M. Allelopathy in freshwater cyanobacteria. Crit. Rev. Microbiol. 2009, 35, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Śliwińska-Wilczewska, S.; Wiśniewska, K.; Konarzewska, Z.; Cieszyńska, A.; Barreiro Felpeto, A.; Lewandowska, A.U.; Latała, A. The current state of knowledge on taxonomy, modulating factors, ecological roles, and mode of action of phytoplankton allelochemicals. Sci. Total Environ. 2021, 773, 145681. [Google Scholar] [CrossRef]
- Kearns, K.D.; Hunter, M.D. Toxin-producing Anabaena flos-aquae induces settling of Chlamydomonas reinhardtii, a competing motile alga. Microb. Ecol. 2001, 42, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Gantar, M.; Berry, J.P.; Thomas, S.; Wang, M.; Perez, R.; Rein, K.S. Allelopathic activity among Cyanobacteria and microalgae isolated from Florida freshwater habitats. FEMS Microbiol. Ecol. 2008, 64, 55–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagmann, L.; Jüttner, F. Fischerellin A, a novel photosystem-II-inhibiting allelochemical of the cyanobacterium Fischerella muscicola with antifungal and herbicidal activity. Tetrahedron Lett. 1996, 37, 6539–6542. [Google Scholar] [CrossRef]
- Jüttner, F.; Lüthi, H. Topology and enhanced toxicity of bound microcystins in Microcystis PCC 7806. Toxicon 2008, 51, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Fastner, J.; Humpage, A. Hepatotoxic cyclic peptides—microcystins and nodularins. In Toxic Cyanobacteria in Water, 2nd ed.; Chorus, I., Welker, M., Eds.; CRC Press: Boca Raton, FL, USA; World Health Organization: Geneva, Switzerland, 2021; pp. 21–52. [Google Scholar]
- Codd, G.A.; Morrison, L.F.; Metcalf, J.S. Cyanobacterial toxins: Risk management for health protection. Toxicol. Appl. Pharmacol. 2005, 203, 264–272. [Google Scholar] [CrossRef]
- Sedmak, B.; Eleršek, T. Microcystins Induce Morphological and Physiological Changes in Selected Representative Phytoplanktons. Microb. Ecol. 2005, 50, 298–305. [Google Scholar] [CrossRef]
- Rapala, J.; Sivonen, K.; Lyra, C.; Niemelä, S.I. Variation of microcystins, cyanobacterial hepatotoxins, in Anabaena spp. as a function of growth stimuli. Appl. Environ. Microbiol. 1997, 63, 2206–2212. [Google Scholar] [CrossRef] [Green Version]
- Pflugmacher, S. Possible allelopathic effects of cyanotoxins, with reference to microcystin-LR, in aquatic ecosystems. Environ. Toxicol. 2002, 17, 407–413. [Google Scholar] [CrossRef]
- Bakr, A.; Alzain, M.N.; Alzamel, N.M.; Loutfy, N. Accumulation of Microcystin from Oscillatoria limnetica Lemmermann and Microcystis aeruginosa (Kützing) in Two Leafy Green Vegetable Crop Plants Lactuca sativa L. and Eruca sativa. Plants 2022, 11, 1733. [Google Scholar] [CrossRef] [PubMed]
- Veerabadhran, M.; Manivel, N.; Sarvalingam, B.; Seenivasan, B.; Srinivasan, H.; Davoodbasha, M.; Yang, F. State-of-the-art review on the ecotoxicology, health hazards, and economic loss of the impact of microcystins and their ultrastructural cellular changes. Aquat. Toxicol. 2023, 256, 106417. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Vo Duy, S.; Munoz, G.; Sauvé, S. Phytotoxic effects of microcystins, anatoxin-a and cylindrospermopsin to aquatic plants: A meta-analysis. Sci. Total Environ. 2022, 810, 152104. [Google Scholar] [CrossRef] [PubMed]
- Máthé, C.; M-Hamvas, M.; Vasas, G.; Garda, T.; Freytag, C. Subcellular Alterations Induced by Cyanotoxins in Vascular Plants—A Review. Plants 2021, 10, 984. [Google Scholar] [CrossRef] [PubMed]
- Kearns, K.D.; Hunter, M.D. Green algal extracellular products regulate antialgal toxin production in a cyanobacterium. Environ. Microbiol. 2000, 2, 291–297. [Google Scholar] [CrossRef]
- Singh, D.P.; Tyagi, M.B.; Kumar, A.; Thakur, J.K.; Kumar, A. Antialgal activity of a hepatotoxin-producing cyanobacterium, Microcystis aeruginosa. World J. Microbiol. Biotechnol. 2001, 17, 15–22. [Google Scholar] [CrossRef]
- Borowitzka, M.A. Chemically-Mediated Interactions in Microalgae. In The Physiology of Microalgae; Springer: Cham, Switzerland, 2016; pp. 321–357. [Google Scholar]
- Humpage, A.; Fastner, J. Cylindrospermopsins. In Toxic Cyanobacteria in Water, 2nd ed.; Chorus, I., Welker, M., Eds.; CRC Press: Boca Raton, FL, USA; World Health Organization: Geneva, Switzerland, 2021; p. 5372. [Google Scholar]
- Bormans, M.; Lengronne, M.; Brient, L.; Duval, C. Cylindrospermopsin Accumulation and Release by the Benthic Cyanobacterium Oscillatoria sp. PCC 6506 under Different Light Conditions and Growth Phases. Bull. Environ. Contam. Toxicol. 2013, 92, 243–247. [Google Scholar] [CrossRef] [Green Version]
- Pietsch, C.; Wiegand, C.; Amé, M.V.; Nicklisch, A.; Wunderlin, D.; Pflugmacher, S. The effects of a cyanobacterial crude extract on different aquatic organisms: Evidence for cyanobacterial toxin modulating factors. Environ. Toxicol. 2001, 16, 535–542. [Google Scholar] [CrossRef]
- Chia, M.A.; Cordeiro-Araujo, M.K.; Lorenzi, A.S.; Bittencourt-Oliveira, M.D.C. Cylindrospermopsin induced changes in growth, toxin production and antioxidant response of Acutodesmus acuminatus and Microcystis aeruginosa under differing light and nitrogen conditions. Ecotoxicol. Environ. Saf. 2017, 142, 189–199. [Google Scholar] [CrossRef]
- Campos, A.; Araújo, P.; Pinheiro, C.; Azevedo, J.; Osório, H.; Vasconcelos, V. Effects on growth, antioxidant enzyme activity and levels of extracellular proteins in the green alga Chlorella vulgaris exposed to crude cyanobacterial extracts and pure microcystin and cylindrospermopsin. Ecotoxicol. Environ. Saf. 2013, 94, 45–53. [Google Scholar] [CrossRef]
- Pinheiro, C.; Azevedo, J.; Campos, A.; Loureiro, S.; Vasconcelos, V. Absence of negative allelopathic effects of cylindrospermopsin and microcystin-LR on selected marine and freshwater phytoplankton species. Hydrobiologia 2012, 705, 27–42. [Google Scholar] [CrossRef] [Green Version]
- Bar-Yosef, Y.; Sukenik, A.; Hadas, O.; Viner-Mozzini, Y.; Kaplan, A. Enslavement in the Water Body by Toxic Aphanizomenon ovalisporum, Inducing Alkaline Phosphatase in Phytoplanktons. Curr. Biol. 2010, 20, 1557–1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, A.; Harel, M.; Kaplan-Levy, R.N.; Hadas, O.; Sukenik, A.; Dittmann, E. The Languages Spoken in the Water Body (or the Biological Role of Cyanobacterial Toxins). Front. Microbiol. 2012, 3, 00138. [Google Scholar] [CrossRef] [Green Version]
- Munkes, B.; Löptien, U.; Dietze, H. Cyanobacteria blooms in the Baltic Sea: A review of models and facts. Biogeosciences 2021, 18, 2347–2378. [Google Scholar] [CrossRef]
- Katsiapi, M.; Moustaka-Gouni, M.; Michaloudi, E.; Kormas, K.A. Phytoplankton and water quality in a Mediterranean drinking-water reservoir (Marathonas Reservoir, Greece). Environ. Monit. Assess. 2011, 181, 563–575. [Google Scholar] [CrossRef]
- Semyalo, R.; Rohrlack, T.; Kayiira, D.; Kizito, Y.S.; Byarujali, S.; Nyakairu, G.; Larsson, P. On the diet of Nile tilapia in two eutrophic tropical lakes containing toxin producing cyanobacteria. Limnologica 2011, 41, 30–36. [Google Scholar] [CrossRef] [Green Version]
- Montoya, H. Algal and cyanobacterial saline biofilms of the Grande Coastal Lagoon, Lima, Peru. Nat. Resour. Environ. Issues 2009, 15, 127–134. [Google Scholar]
- Khona, D.K.; Shirolikar, S.M.; Gawde, K.K.; Hom, E.; Deodhar, M.A.; D’Souza, J.S. Characterization of salt stress-induced palmelloids in the green alga, Chlamydomonas reinhardtii. Algal Res. 2016, 16, 434–448. [Google Scholar] [CrossRef]
- Polle, J.E.W.; Roth, R.; Ben-Amotz, A.; Goodenough, U. Ultrastructure of the green alga Dunaliella salina strain CCAP19/18 (Chlorophyta) as investigated by quick-freeze deep-etch electron microscopy. Algal Res. 2020, 49, 101953. [Google Scholar] [CrossRef]
- Sivarajah, B.; Simmatis, B.; Favot, E.J.; Palmer, M.J.; Smol, J.P. Eutrophication and climatic changes lead to unprecedented cyanobacterial blooms in a Canadian sub-Arctic landscape. Harmful Algae 2021, 105, 102036. [Google Scholar] [CrossRef]
- Teneva, I.; Basheva, D.; Mladenova, T.; Stoyanov, P.; Belkinova, D.; Mladenov, R. Species Composition and Toxic Potential of Cyanobacteria in Some Western Rhodopes Dams. Ecol. Balk. 2018, 10, 111–121. [Google Scholar]
- Wu, P.; Lu, Y.; Lu, Y.; Dai, J.; Huang, T. Response of the photosynthetic activity and biomass of the phytoplankton community to increasing nutrients during cyanobacterial blooms in Meiliang Bay, Lake Taihu. Water Environ. Res. 2019, 92, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Ballot, A.; Swe, T.; Mjelde, M.; Cerasino, L.; Hostyeva, V.; Miles, C.O. Cylindrospermopsin- and Deoxycylindrospermopsin-Producing Raphidiopsis raciborskii and Microcystin-Producing Microcystis spp. in Meiktila Lake, Myanmar. Toxins 2020, 12, 232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Akbar, S.; Sun, Y.; Gu, L.; Zhang, L.; Lyu, K.; Huang, Y.; Yang, Z. Cyanobacterial dominance and succession: Factors, mechanisms, predictions, and managements. J. Environ. Manag. 2021, 297, 113281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yang, Z.; Shi, X.; Yu, Y. The synergistic effect of rising temperature and declining light boosts the dominance of bloom-forming cyanobacteria in spring. Harmful Algae 2022, 116, 102252. [Google Scholar] [CrossRef] [PubMed]
- Lenard, T.; Poniewozik, M. Planktothrix agardhii versus Planktothrix rubescens: Separation of Ecological Niches and Consequences of Cyanobacterial Dominance in Freshwater. Int. J. Environ. Res. Public Health 2022, 19, 14897. [Google Scholar] [CrossRef]
- Teneva, I.; Belkinova, D.; Mladenov, R.; Stoyanov, P.; Moten, D.; Basheva, D.; Kazakov, S.; Dzhambazov, B. Phytoplankton composition with an emphasis of Cyanobacteria and their toxins as an indicator for the ecological status of Lake Vaya (Bulgaria)—part of the Via Pontica migration route. Biodivers. Data J. 2020, 8, e57507. [Google Scholar] [CrossRef]
- Chia, M.; Kramer, B.; Jankowiak, J.; Bittencourt-Oliveira, M.; Gobler, C. The Individual and Combined Effects of the Cyanotoxins, Anatoxin-a and Microcystin-LR, on the Growth, Toxin Production, and Nitrogen Fixation of Prokaryotic and Eukaryotic Algae. Toxins 2019, 11, 43. [Google Scholar] [CrossRef] [Green Version]
- Dzhambazov, B.; Moten, D.; Basheva, D.; Belkinova, D.; Teneva, I. The allelopathic effects of toxin-producing cyanobacteria are pH-dependent. In Proceedings of the 18th International Multidisciplinary Scientific GeoConference SGEM 2018, Albena, Bulgaria, 30 June–9 July 2018; pp. 905–912. [Google Scholar]
- Omidi, A.; Esterhuizen-Londt, M.; Pflugmacher, S. Interspecies interactions between Microcystis aeruginosa PCC 7806 and Desmodesmus subspicatus SAG 86.81 in a co-cultivation system at various growth phases. Environ. Int. 2019, 131. [Google Scholar] [CrossRef]
- Cheloni, G.; Slaveykova, V.I. Morphological plasticity in Chlamydomonas reinhardtii and acclimation to micropollutant stress. Aquat. Toxicol. 2021, 231, 105711. [Google Scholar] [CrossRef]
- Ding, W.-X.; Shen, H.-M.; Ong, C.-N. Critical Role of Reactive Oxygen Species and Mitochondrial Permeability Transition in Microcystin-Induced Rapid Apoptosis in Rat Hepatocytes. Hepatology 2000, 32, 547–555. [Google Scholar] [CrossRef]
- Campos, A.; Vasconcelos, V. Molecular Mechanisms of Microcystin Toxicity in Animal Cells. Int. J. Mol. Sci. 2010, 11, 268–287. [Google Scholar] [CrossRef] [Green Version]
- Dilov, C.; Bozkova, M.; Staev, S. A practical block for cultivation of microalgae in laboratory conditions. Dokl Akad Selskoch Nauk Bolg 1972, 4, 169–175. [Google Scholar]
- Bischoff, H.; Bold, H. Phycological Studies IV. Some Soil Algae From Enchanted Rock and Related Algal Species; University of Texas: Austin, TX, USA, 1963; Volume 6318. [Google Scholar]
- Eddy, B.P. The Suitability of Some Algae for Mass Cultivation for Food, with Special Reference toDunaliella bioculata. J. Exp. Bot. 1956, 7, 372–380. [Google Scholar] [CrossRef]
- WHO. Guidelines for Drinking-Water Quality: Fourth Edition Incorporating the First and Second Addenda; WHO: Geneva, Switzerland, 2022; p. 614. [Google Scholar]
- Marques, S.M.; Esteves da Silva, J.C.G. Firefly bioluminescence: A mechanistic approach of luciferase catalyzed reactions. IUBMB Life 2009, 61, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Bilger, W.; Björkman, O. Temperature dependence of violaxanthin de-epoxidation and non-photochemical fluorescence quenching in intact leaves of Gossypium hirsutum L. and Malva parviflora L. Planta 1991, 184, 226–234. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
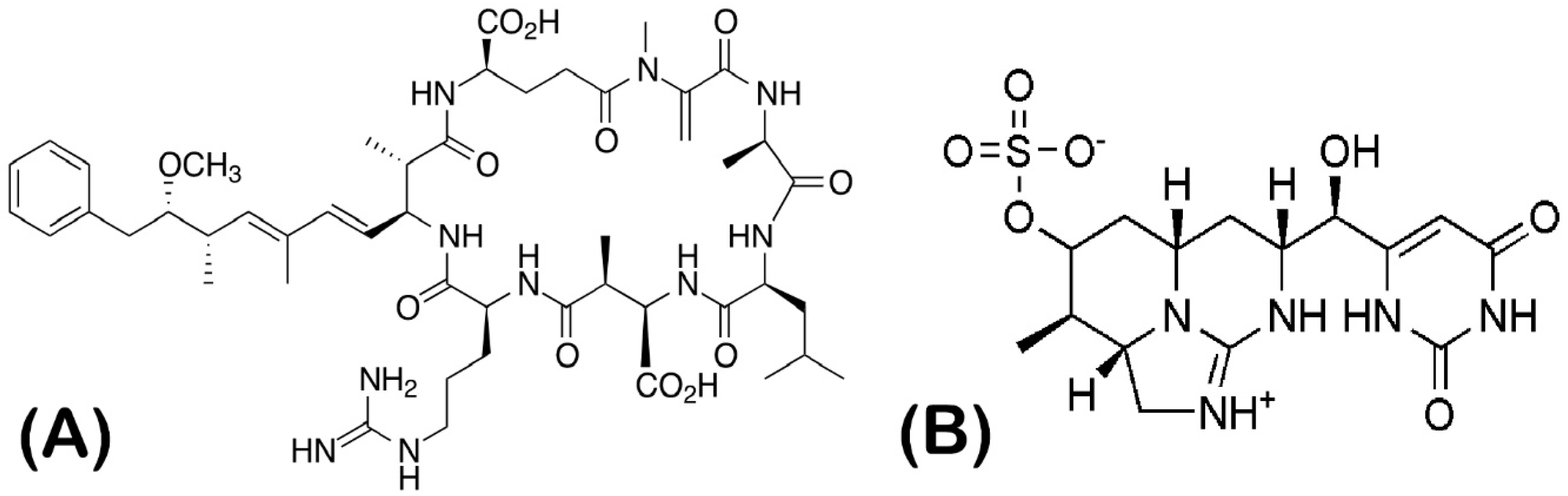{kind=link}
| Algal Species | Microcystin-LR (MC-LR) § | Cylindrospermopsin (CYL) § | ||||||
|---|---|---|---|---|---|---|---|---|
| 15 h | 24 h | 48 h | 72 h | 15 h | 24 h | 48 h | 72 h | |
| C. asymmetrica | − | 48.57 # | 70 ϕ | 74.18 ϕ | − | 42.86 * | 60 # | 72.73 ϕ |
| D. salina | 4 | 15.49 | 35.48 * | 58.86 ϕ | − | 11.97 | 51.61 # | 60 ϕ |
| S. obtusiusculus | − | 22 * | 60 # | 66.67 ϕ | 25 * | 25 * | 60 # | 83.33 ϕ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teneva, I.; Velikova, V.; Belkinova, D.; Moten, D.; Dzhambazov, B. Allelopathic Potential of the Cyanotoxins Microcystin-LR and Cylindrospermopsin on Green Algae. Plants 2023, 12, 1403. https://doi.org/10.3390/plants12061403
Teneva I, Velikova V, Belkinova D, Moten D, Dzhambazov B. Allelopathic Potential of the Cyanotoxins Microcystin-LR and Cylindrospermopsin on Green Algae. Plants. 2023; 12(6):1403. https://doi.org/10.3390/plants12061403
Chicago/Turabian StyleTeneva, Ivanka, Violeta Velikova, Detelina Belkinova, Dzhemal Moten, and Balik Dzhambazov. 2023. "Allelopathic Potential of the Cyanotoxins Microcystin-LR and Cylindrospermopsin on Green Algae" Plants 12, no. 6: 1403. https://doi.org/10.3390/plants12061403