
Alleviation of Associated Drought and Salinity Stress’ Detrimental Impacts on an Eggplant Cultivar (‘Bonica F1’) by Adding Biochar


Abstract
:1. Introduction
2. Results
2.1. Plant Phenology, Growth, and Yield Properties
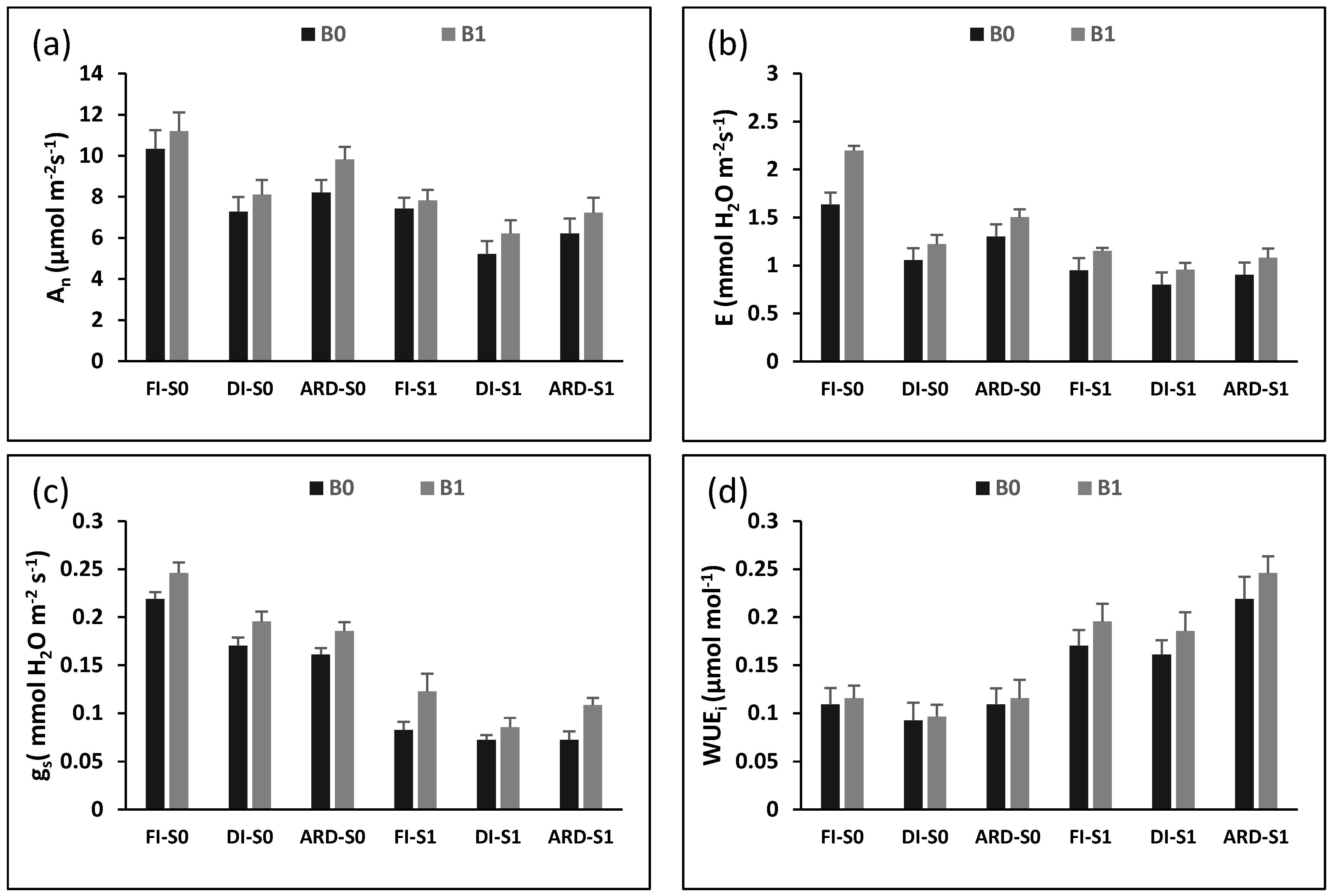
2.2. Physiological Parameters
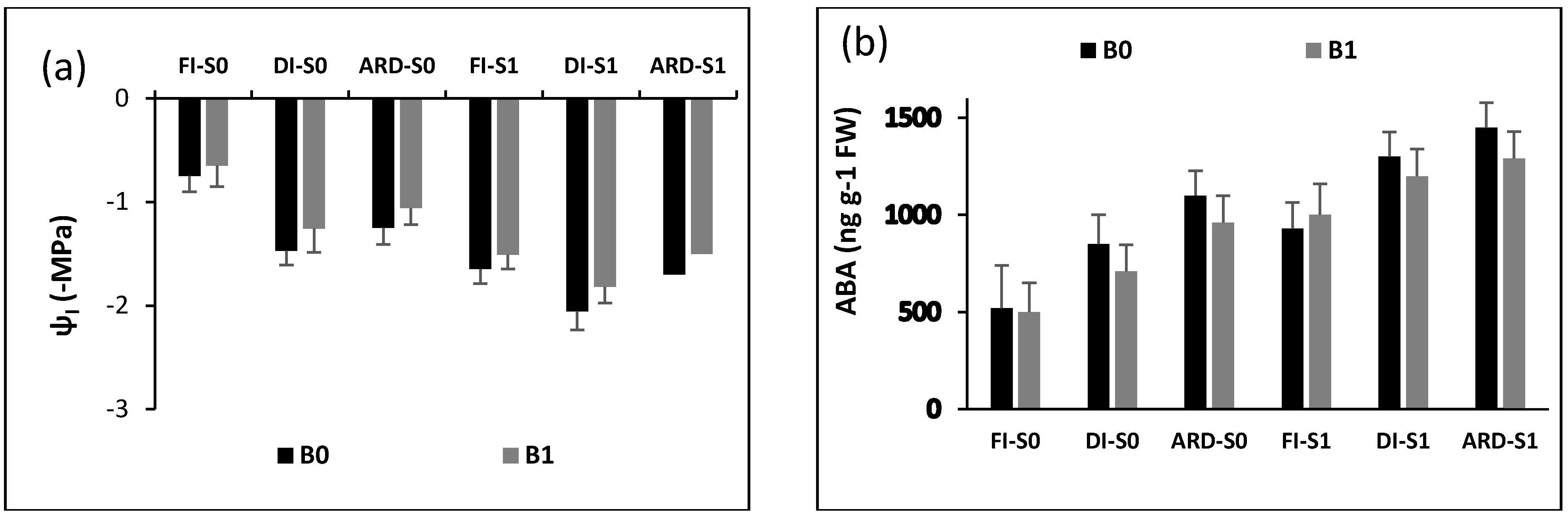
2.3. Plant Water Potential and Abscisic Acid Content
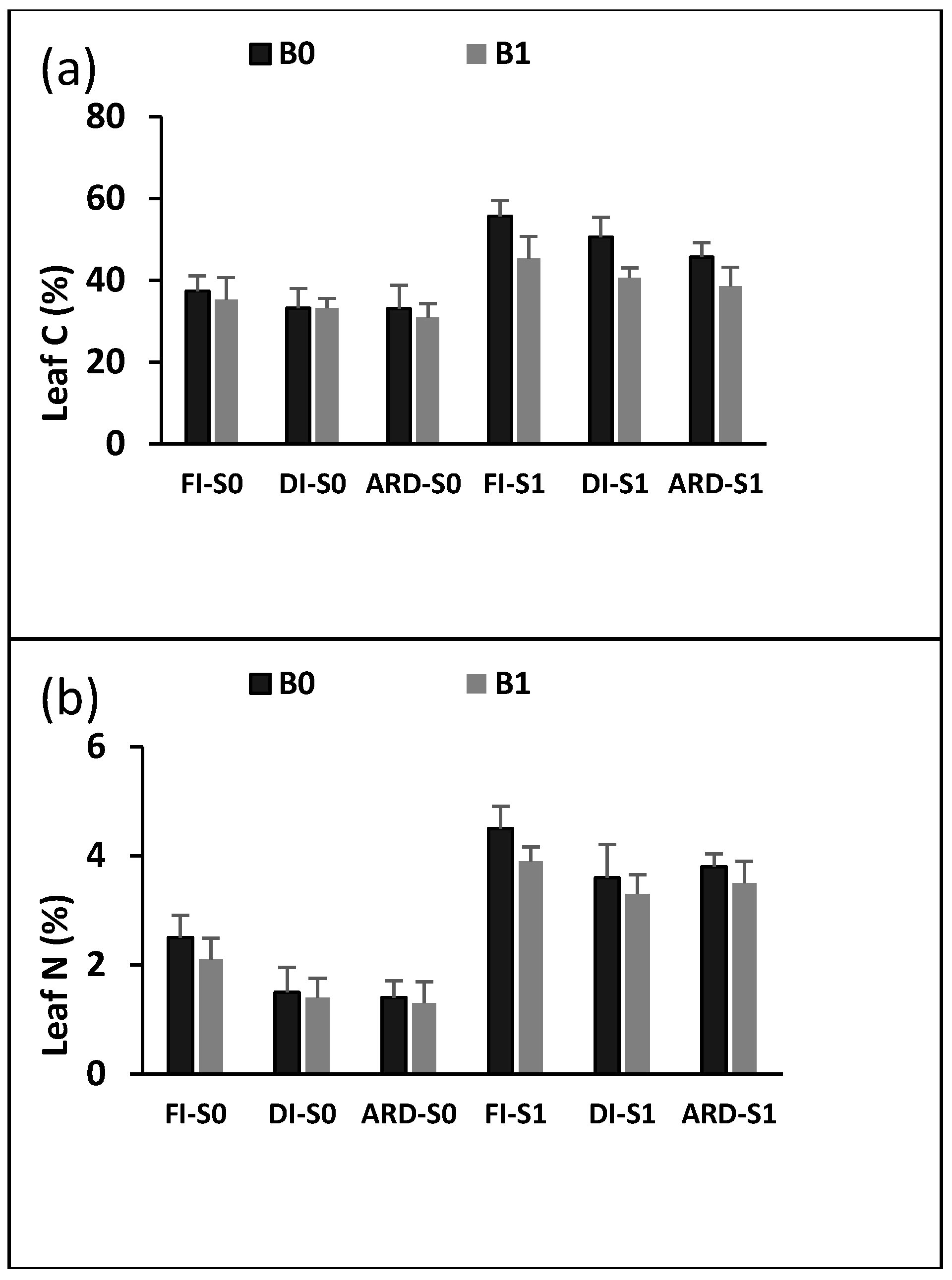
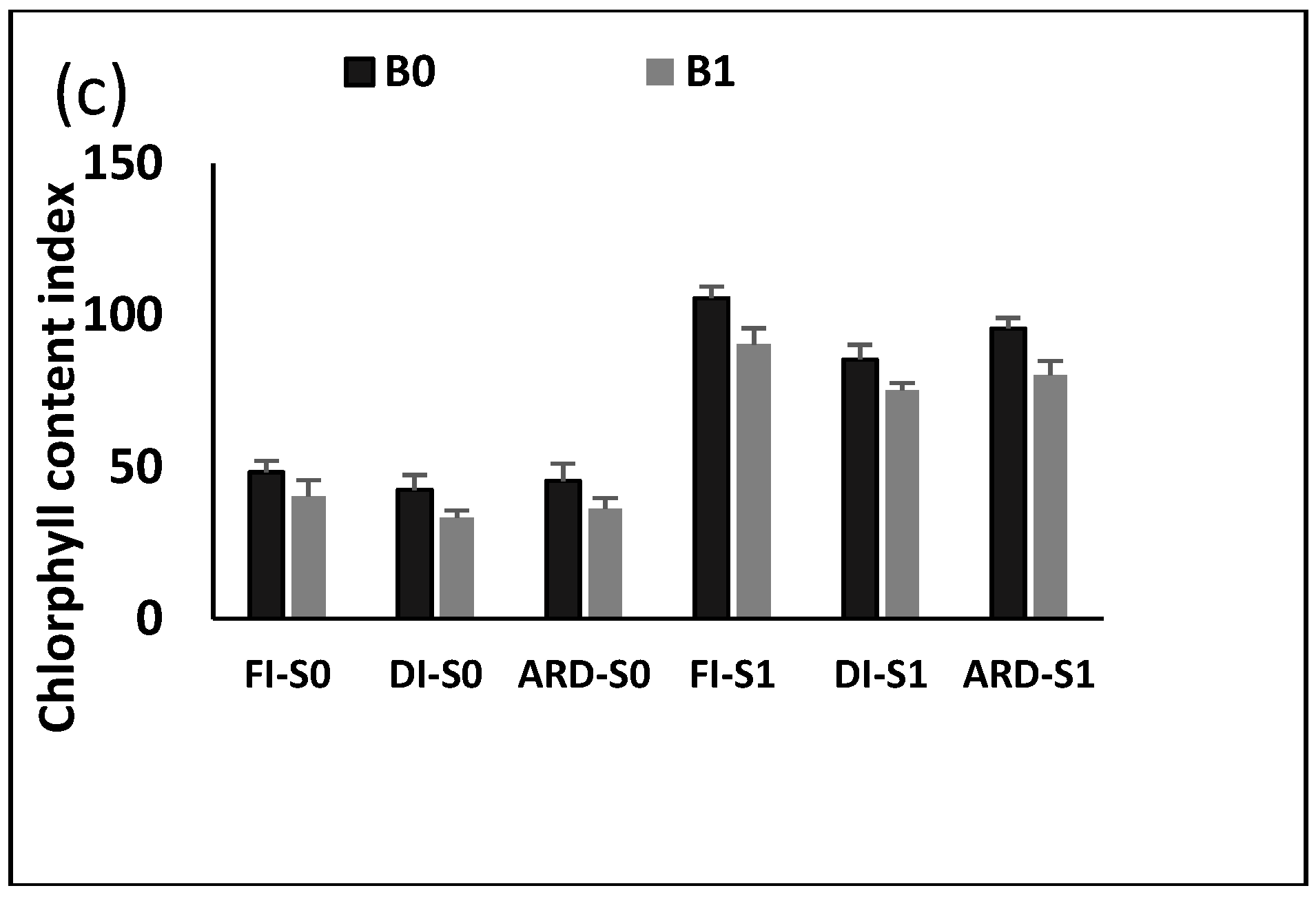
2.4. Leaf C, and N Content and Chlorophyll Content Index
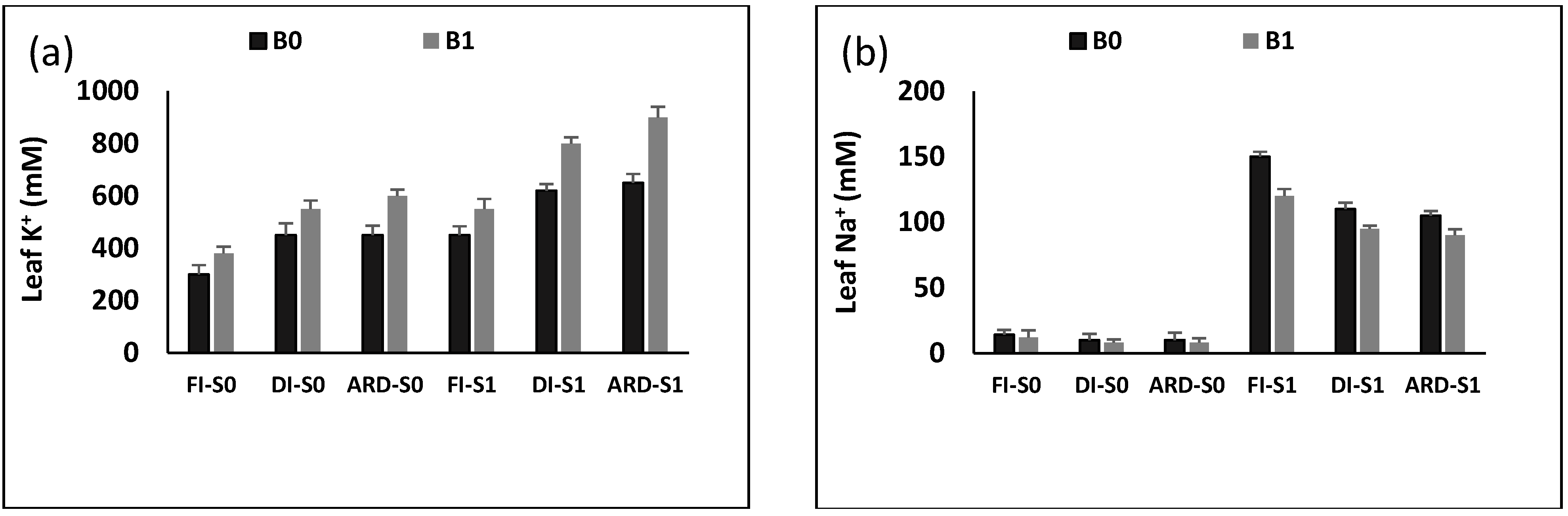
2.5. Na+ and K+ Content
3. Discussion
4. Materials and Methods
4.1. Biochar Description
4.2. Pot Experiment
4.3. Irrigation Treatments
4.4. Phylogeny, Yield and Growth Parameters
4.5. Physiological Parameters and Water Status Measurements
4.6. Leaf Abscisic Acid (ABA), Nitrogen, Carbon, Na+, and K+ Content
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Glick, B.R. Beneficial Plant-Bacterial Interactions; Springer: Berlin/Heidelberg, Germany, 2020; 383p. [Google Scholar]
- Blum, A. Drought resistance—Is it really a complex trait? Funct. Plant Biol. 2011, 38, 753–757. [Google Scholar] [CrossRef]
- FAO. The State of Food and Agriculture. 2016. Available online: https://www.fao.org/publications/sofa/sofa-2021/en/ (accessed on 3 March 2021).
- Lobell, D.; Roberts, M.; Schlenker, W.; Braun, N.; Little, B.; Rejesus, R.; Hammer, G. Greater sensitivity to drought accompanies maize yield increase in the U.S. Midwest. Nature 2014, 344, 516–519. [Google Scholar] [CrossRef]
- Parvin, S.; Uddin, S.; Fitzgerald, G.; Tausz-Posch, S.; Armstrong, R.; Tausz, M. Free air CO2 enrichment (FACE) improves water use efficiency and moderates drought effect on N2 fixation of Pisum sativum L. Plant Soil 2019, 436, 587–606. [Google Scholar] [CrossRef]
- IPCC Climate Change. The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Edwards, C.; Ewers, B.; McClung, C.; Lou, P.; Weinig, C. Quantitative variation in water-use efficiency across water regimes and its relationship with circadian vegetative, reproductive, and leaf gas-exchange traits. Mol. Plant 2012, 5, 653–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, M.; Farooq, S.; Hasan, W.; Ul-Allah, S.; Tanveer, M.; Farooq, M.; Nawaz, A. Drought stress in sunflower: Physiological effects and its management through breeding and agronomic alternatives. Agric. Water Manag. 2018, 201, 152–166. [Google Scholar] [CrossRef]
- Zhang, Y.; Ding, J.; Wang, H. Biochar addition alleviate the negative effects of drought and salinity stress on soybean productivity and water use efficiency. BMC Plant Biol. 2020, 20, 288. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Turner, N.; Yan, G.; Li, F.; Siddique, K. Flower numbers, pod production, pollen viability, and pistil function are reduced, and flower and pod abortion increased in chickpea (Cicer arietinum L.) under terminal drought. J. Exp. Bot. 2010, 61, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Hammer, G.; Dong, Z.; McLean, G.; Doherty, A.; Messina, C.; Schussler, J.; Zinselmeier, C.; Paszkiewicz, S.; Cooper, M. Can changes in canopy and/or root system architecture explain historical maize yield trends in the US corn belt? Crop Sci. 2009, 49, 299–312. [Google Scholar] [CrossRef]
- Munns, R.; Gilliham, M. Salinity tolerance of crops—What is the cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef] [Green Version]
- Hannachi, S.; Van Labeke, M.C. Salt stress affects germination, seedling growth and physiological responses differentially in eggplant cultivars (Solanum melongena L.). Sci. Hortic. 2018, 228, 56–65. [Google Scholar] [CrossRef]
- Stavridou, E.; Hastings, A.; Webster, R.; Robson, P. The impact of soil salinity on the yield, composition and physiology of the bioenergy grass Miscanthus × giganteus. GCB Bioenergy 2017, 9, 92–104. [Google Scholar] [CrossRef] [Green Version]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Wang, C.; Wang, W.; Jiang, L. Mitogen-activated protein kinase 6 controls root growth in Arabidopsis by modulating Ca2+ −based Na+ flux in root cell under salt stress. J. Plant Physiol. 2014, 171, 26–34. [Google Scholar] [CrossRef] [Green Version]
- Jeffery, S.; Verheijen, F.; van der Velde, M.; Bastos, A. A quantitative review of the effects of biochar application to soils on crop productivity using metaanalysis. Agric. Ecosyst. Environ. 2011, 144, 175–187. [Google Scholar] [CrossRef]
- Raboin, L.; Razafimahafaly, A.; Rabenjarisoa, M.; Rabary, B.; Dusserre, J.; Becquer, T. Improving the fertility of tropical acid soils: Liming versus biochar application? A long-term comparison in the highlands of Madagascar. Field Crops Res. 2016, 199, 99–108. [Google Scholar] [CrossRef]
- Antala, M.; Sytar, O.; Rastogi, A.; Brestic, M. Potential of Karrikins as Novel Plant Growth Regulators in Agriculture. Plants 2020, 9, 43. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Chen, B.; Zhu, L.; Xing, B. Effects and mechanisms of biochar-microbe interactions in soil improvement and pollution remediation: A review. Environ. Pollut. 2017, 227, 98–115. [Google Scholar] [CrossRef]
- Atkinson, C.J.; Fitzgerald, J.D.; Hipps, N.A. Potential mechanisms for achieving agricultural benefits from biochar application to temperate soils: A review. Plant Soil 2010, 337, 1–18. [Google Scholar] [CrossRef]
- Lehmann, J. Bio-energy in the black. Front. Ecol. Environ. 2007, 5, 381–387. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, S.S.; Andersen, M.N.; Liu, F. Residual effects of biochar on improving growth, physiology and yield of wheat under salt stress. Agr. Water Manag. 2015, 158, 61–68. [Google Scholar] [CrossRef]
- Chapman, M.A. Introduction: The Importance of Eggplant. In Compendium of Plant Genomes; Springer: Singapore, 2019; pp. 1–10. [Google Scholar]
- FAO. FAOSTAT Statistics Database; FAO: Rome, Italy, 2020. [Google Scholar]
- Kunert, K.; Vorster, B.; Fenta, B.; Kibido, T.; Dionisio, G.; Foyer, C. Drought stress responses in soybean roots and nodules. Front. Plant Sci. 2016, 7, 1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.; Du, Y.; Wang, T.; Turner, N.; Yang, R.; Jin, Y.; Xi, Y.; Zhang, C.; Cui, T.; Fang, X.; et al. Conserved water use improves the yield performance of soybean (Glycine max (L.) Merr.) under drought. Agric. Water Manag. 2017, 179, 236–245. [Google Scholar] [CrossRef]
- Baroowa, B.; Gogoi, N. Biochemical changes in black gram and green gram genotypes after imposition of drought stress. J. Food Legumes 2014, 27, 350–353. [Google Scholar]
- Akhtar, S.S.; Li, G.T.; Andersen, M.N.; Liu, F.L. Biochar enhances yield and quality of tomato under reduced irrigation. Agric. Water Manag. 2014, 138, 37–44. [Google Scholar] [CrossRef]
- Ali, S.; Rizwan, M.; Qayyum, M.F.; Ok, Y.S.; Ibrahim, M.; Riaz, M.; Arif, M.S.; Hafeez, F.; Al-Wabel, M.I.; Shahzad, A.N. Biochar soil amendment on alleviation of drought and salt stress in plants: A critical review. Environ. Sci. Pollut. Res. 2017, 24, 12700–12712. [Google Scholar] [CrossRef]
- Baiamonte, G.; De Pasquale, C.; Marsala, V.; Cimo, G.; Alonzo, G.; Crescimanno, G.; Conte, P. Structure alteration of a sandy-clay soil by biochar amendments. J. Soils Sedim. 2015, 15, 816–824. [Google Scholar] [CrossRef]
- Artiola, J.F.; Rasmussen, C.; Freitas, R. Effects of a Biochar-Amended Alkaline Soil on the Growth of Romaine Lettuce and Bermudagrass. Soil Sci. 2012, 177, 561–570. [Google Scholar] [CrossRef]
- Akhtar, S.S.; Andersen, M.N.; Naveed, M.; Zahir, Z.A.; Liu, F.L. Interactive effect of biochar and plant growth-promoting bacterial endophytes on ameliorating salinity stress in maize. Funct. Plant Biol. 2015, 42, 770–781. [Google Scholar] [CrossRef]
- Gurav, R.; Virdhaval, N.; Chetan, A.; Govind, V.; Shashi, K.; Yung-Hun, Y.; Vishwas, B.; Jyoti, J. Microbial degradation of poultry feather biomass in a constructed bioreactor and application of hydrolysate as bioenhancer to vegetable crops. Environ. Sci. Pollut. Res. 2020, 27, 2027–2035. [Google Scholar] [CrossRef]
- Yildirim, E.; Melek, E.; Metin, T. Impact of biochar in mitigating the negative effect of drought stress on cabbage seedlings. J. Soil Sci. Plant Nutr. 2021, 21, 2297–2309. [Google Scholar] [CrossRef]
- Tahjib-UI-Arif, M.; Sohag, A.A.M.; Afrin, S.; Bashar, K.K.; Afrin, T.; Mahamud, A.G.M.S.U.; Polash, M.A.S.; Hossain, M.T.; Sohel, M.A.T.; Brestic, M.; et al. Differential Response of Sugar Beet to Long-Term Mild to Severe Salinity in a Soil-Pot Culture. Agriculture 2019, 9, 223. [Google Scholar] [CrossRef] [Green Version]
- Saifullah; Dahlawi, S.; Naeem, A.; Rengel, Z.; Naidu, R. Biochar application for the remediation of salt-affected soils: Challenges and opportunities. Sci. Total Environ. 2018, 625, 320–335. [Google Scholar] [CrossRef] [PubMed]
- Palansooriya, K.N.; Ok, Y.S.; Awad, Y.M.; Lee, S.S.; Sung, J.-K.; Koutsospyros, A.; Moon, D.H. Impacts of biochar application on upland agriculture: A review. J. Environ. Manag. 2019, 234, 52–64. [Google Scholar] [CrossRef]
- Yang, A.; Akhtar, S.S.; Li, L.; Fu, Q.; Li, Q.; Naeem, M.A.; He, X.; Zhang, Z.; Jacobsen, S.E. Biochar mitigates combined effects of drought and salinity stress in quinoa. Agronomy 2020, 10, 912. [Google Scholar] [CrossRef]
- Kammann, C.; Graber, E.R. Biochar effects on plant eco-physiology. In Biochar for Environmental Management, 2nd ed.; Lehman, J., Joseph, S., Eds.; Sci Tech, Implementation Abingdon; Routledge: London, UK, 2015; pp. 391–420. [Google Scholar]
- Sun, Z.W.; Ren, L.K.; Fan, J.W.; Li, Q.; Wang, K.J.; Guo, M.M.; Wang, L.; Li, J.; Zhang, G.X.; Yang, Z.Y. Salt response of photosynthetic electron transport system in wheat cultivars with contrasting tolerance. Plant Soil Environ. 2016, 62, 515–521. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Davies, W.J. ABA in roots and leaves of flooded pea-plants. J. Exp. Bot. 1987, 38, 649–659. [Google Scholar] [CrossRef]
- Yang, A.; Akhtar, S.S.; Amjad, M.; Iqbal, S.; Jacobsen, S.E. Growth and Physiological Responses of Quinoa to Drought and Temperature Stress. J. Agron. Crop Sci. 2016, 202, 445–453. [Google Scholar] [CrossRef]
- Degaris, K.A.; Walker, R.R.; Loveys, B.R.; Tyerman, S.D. Comparative effects of deficit and partial root-zone drying irrigation techniques using moderately saline water on ion partitioning in Shiraz and Grenache grapevines. Aust. J. Grape Wine Res. 2016, 22, 296–306. [Google Scholar] [CrossRef]
- Akhtar, S.S.; Andersen, M.N.; Liu, F. Biochar Mitigates Salinity Stress in Potato. J. Agron. Crop. Sci. 2015, 201, 368–378. [Google Scholar] [CrossRef]
- Taghizadeh-Toosi, A.; Clough, T.J.; Sherlock, R.R.; Condron, L.M. Biochar adsorbed ammonia is bioavailable. Plant Soil 2012, 350, 57–69. [Google Scholar] [CrossRef]
- Clough, T.J.; Condron, L.M. Biochar and the Nitrogen Cycle: Introduction. J. Environ. Qual. 2010, 39, 1218–1223. [Google Scholar] [CrossRef] [PubMed]
- Tammeorg, P.; Simojoki, A.; Makela, P.; Stoddard, F.L.; Alakukku, L.; Helenius, J. Biochar application to a fertile sandy clay loam in boreal conditions: Effects on soil properties and yield formation of wheat, turnip rape and faba bean. Plant Soil 2014, 374, 89–107. [Google Scholar] [CrossRef]
- Zheng, H.; Wang, Z.; Deng, X.; Herbert, S.; Xing, B. Impacts of adding biochar on nitrogen retention and bioavailability in agricultural soil. Geoderma 2013, 206, 32–39. [Google Scholar] [CrossRef]
- Jeffery, S.; Bezemer, T.M.; Cornelissen, G.; Kuyper, T.W.; Lehmann, J.; Mommer, L.; Sohi, S.P.; van de Voorde, T.F.J.; Wardle, D.A.; van Groenigen, J.W. The way forward in biochar research: Targeting trade-offs between the potential wins. Glob. Change Biol. Bioenergy 2015, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Manzoor, M.; Gul, S.; Hidayatullah, K. Influence of Biochars on Yield and Nitrogen and Phosphorus Use Efficiency of Pisum sativum under Groundwater and Wastewater Irrigation in Arid Climate. Commun. Soil Sci. Plant Anal. 2019, 50, 1563–1579. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, J.W.; Wang, S.Q.; Xing, G.X. Successive straw biochar application as a strategy to sequester carbon and improve fertility: A pot experiment with two rice/wheat rotations in paddy soil. Plant Soil 2014, 378, 279–294. [Google Scholar] [CrossRef]
- Priha, O.; Smolander, A. Nitrogen transformations in soil under Pinus sylvestris, Picea abies and Betula pendula at two forest sites. Soil Biol. Biochem. 1999, 31, 965–977. [Google Scholar] [CrossRef]
- O’Kelly, B.C. Accurate Determination of Moisture Content of Organic Soils Using the Oven Drying Method. Dry. Technol. 2004, 22, 1767–1776. [Google Scholar] [CrossRef]
- Philosophhadas, S.; Hadas, E.; Aharoni, N. Characterization and use in ELISA of a new monoclonal-antibody for quantitation of abscisic-acid in senescing rice leaves. Plant Growth Regul. 1993, 12, 71–78. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Flowering Time (FT) (Days) | Plant Height (PH) (cm) | Aerial Biomass (AB) (g) | Fruit per Plant (N) | Mean Fresh Weight per Fruit (FW) (g) | ||
|---|---|---|---|---|---|---|---|
| S0 | FI | B0 | 46.75 ± 0.32 g | 11.4 ± 0.5 b | 120.3 ± 1.2 b | 8 ± 0.1 b | 180 ± 1.9 b |
| B1 | 43.5 ± 0.42 h | 13.3 ± 0.2 a | 150.1 ± 3.5 a | 10 ± 0.2 a | 195 ± 3.6 a | ||
| DI | B0 | 52.34 ± 0.26 e | 7.1 ± 0.1 d | 70.5 ± 2.1 f | 5 ± 0.4 d | 140 ± 2.5 d | |
| B1 | 49.36 ± 0.41 f | 8.5 ± 0.4 c | 85.3 ± 5.2 f | 6 ± 0.6 c | 150 ± 1.9 c | ||
| ARD | B0 | 51.02 ± 0.31 e | 9.4 ± 0.2 c | 95.2 ± 3.2 e | 6 ± 0.1 d | 147 ± 4.1 d | |
| B1 | 49.96 ± 0.41 f | 11.1 ± 0.3 b | 110.1 ± 1.7 d | 7 ± 0.3 c | 155 ± 2.9 c | ||
| S1 | FI | B0 | 55.56 ± 0.32 d | 6.5 ± 0.3 f | 100.2 ± 4.1 d | 3 ± 0.2 e | 70 ± 1.3 e |
| B1 | 54.10 ± 0.40 d | 7.6 ± 0.2 e | 118.3 ± 3.3 c | 3.5 ± 0.2 e | 75 ± 5.0 e | ||
| DI | B0 | 63.08 ± 0.42 a | 5.3 ± 0.5 h | 45.7 ± 1.5 h | 1.5 ± 0.3 h | 35 ± 2.2 h | |
| B1 | 60.54 ± 0.26 b | 5.7 ± 0.6 g | 55.8 ± 2.0 h | 1.8 ± 0.1 g | 40 ± 3.1 g | ||
| ARD | B0 | 59.17 ± 0.40 b | 5.8 ± 0.4 g | 67.8 ± 3.9 g | 1.9 ± 0.2 g | 45 ± 2.0 g | |
| B1 | 56.12 ± 0.30 c | 6.5 ± 0.2 f | 75.9 ± 1.7 f | 2.4 ± 0.2 f | 55 ± 1.9 f |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hannachi, S.; Signore, A.; Mechi, L. Alleviation of Associated Drought and Salinity Stress’ Detrimental Impacts on an Eggplant Cultivar (‘Bonica F1’) by Adding Biochar. Plants 2023, 12, 1399. https://doi.org/10.3390/plants12061399
Hannachi S, Signore A, Mechi L. Alleviation of Associated Drought and Salinity Stress’ Detrimental Impacts on an Eggplant Cultivar (‘Bonica F1’) by Adding Biochar. Plants. 2023; 12(6):1399. https://doi.org/10.3390/plants12061399
Chicago/Turabian StyleHannachi, Sami, Angelo Signore, and Lassaad Mechi. 2023. "Alleviation of Associated Drought and Salinity Stress’ Detrimental Impacts on an Eggplant Cultivar (‘Bonica F1’) by Adding Biochar" Plants 12, no. 6: 1399. https://doi.org/10.3390/plants12061399