
Prediction of the Potential Distribution of the Endangered Species Meconopsis punicea Maxim under Future Climate Change Based on Four Species Distribution Models


Abstract
:1. Introduction
2. Results
2.1. Evaluation of Model Prediction Accuracy and Significance of Bioclimatic Variables
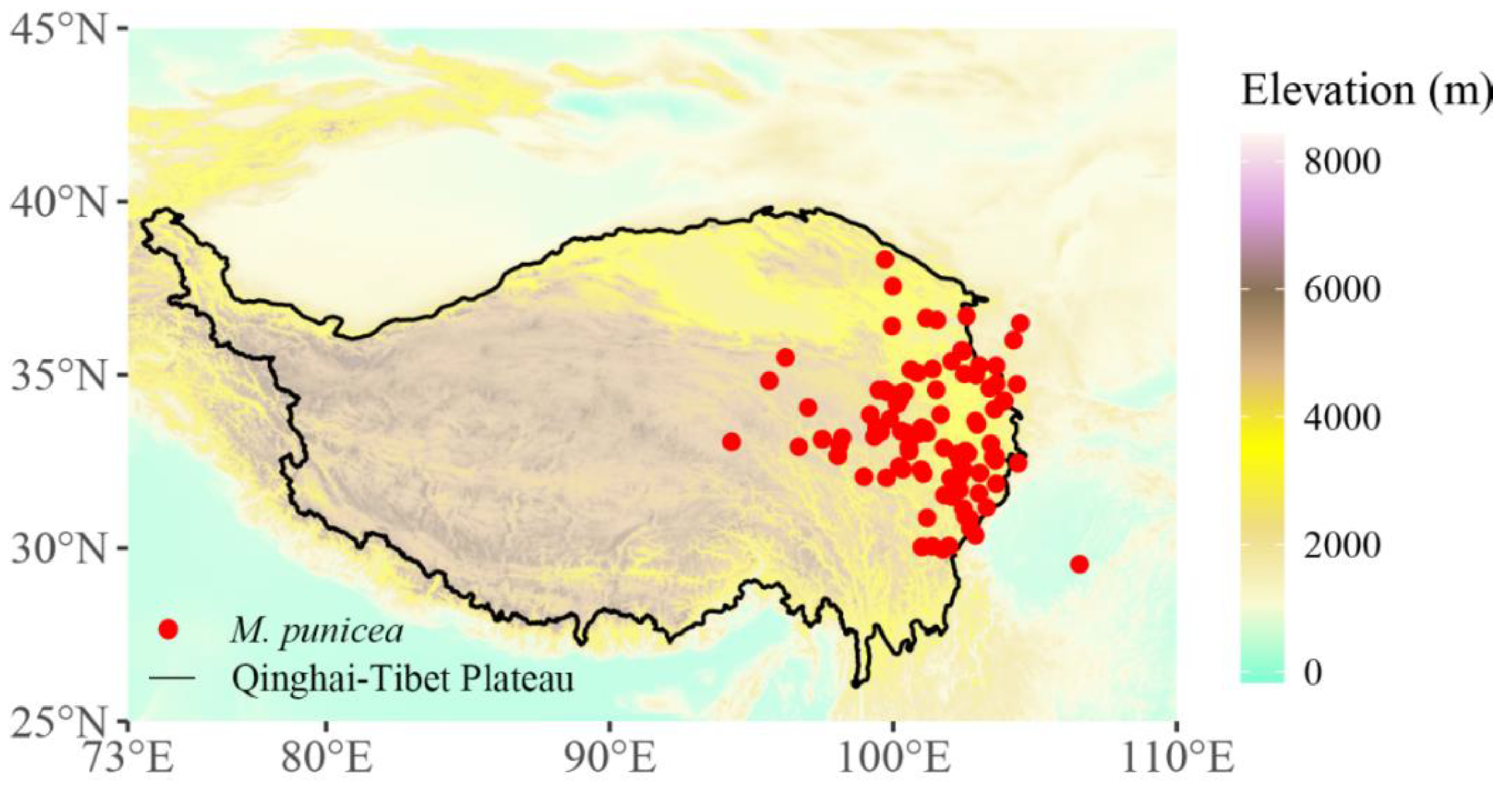
2.2. Potential Distribution of M. punicea in Current Climate
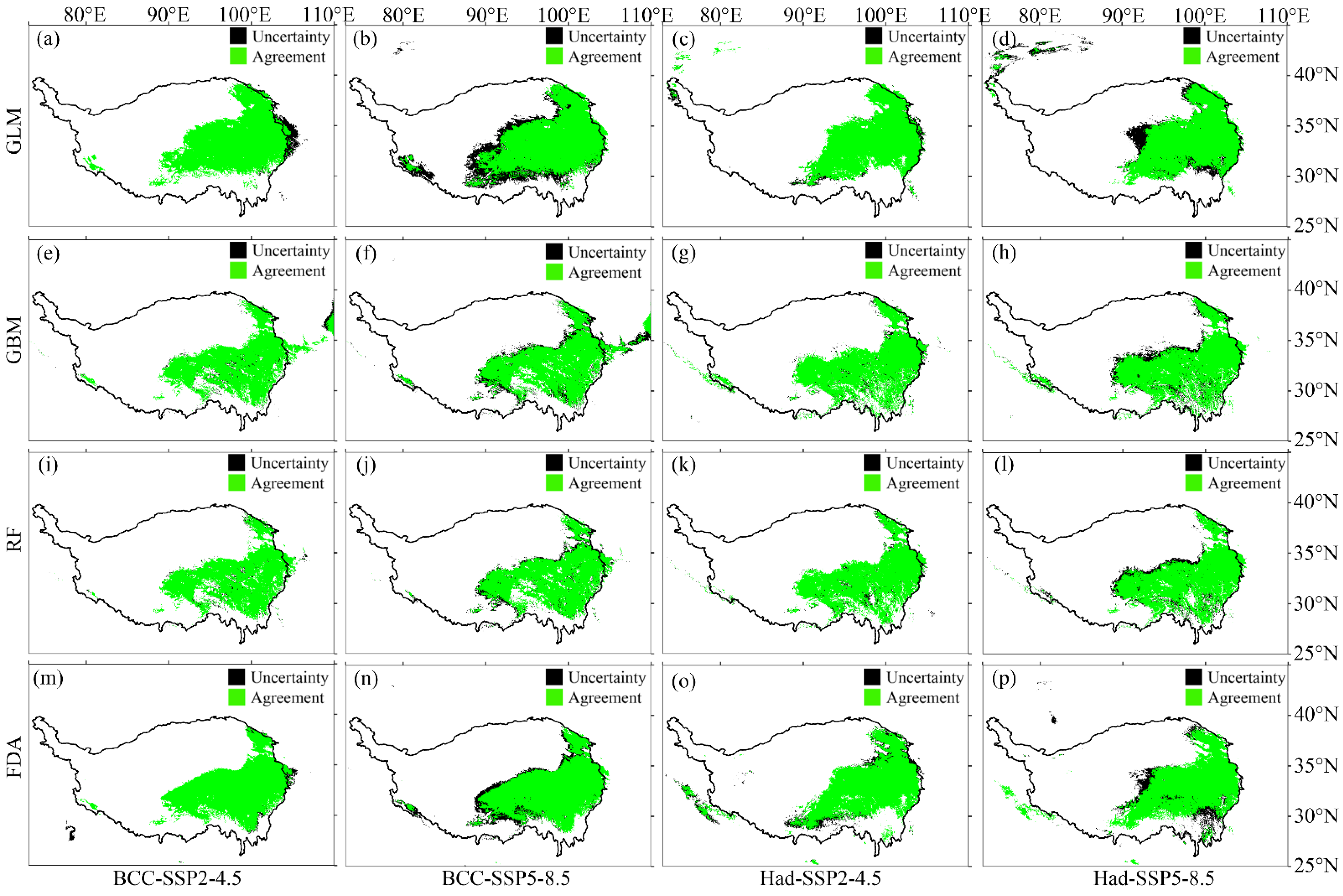
2.3. Changes in Distribution of M. punicea under Future Climate Change
3. Discussion
3.1. Comparison of Prediction Results of Four SDMs
3.2. Significant Variables Affecting the Distribution of M. punicea
3.3. Impacts of Climate Change on the Potential Distribution of M. Punicea
3.4. Protection Strategies for M. punicea
4. Materials and Methods
4.1. Overview
4.2. Species Occurrence Data
4.3. Environmental Variables
4.4. Species Distribution Model
4.4.1. Data Preparation
4.4.2. Parameter Setting of the Model
4.5. Data Analysis
4.5.1. Model Evaluation Metrics
4.5.2. Comparison of Current and Future Potential Distribution Areas
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Araújo, M.B.; Rahbek, C. How does climate change affect biodiversity? Science 2006, 313, 1396–1397. [Google Scholar] [CrossRef]
- Beaumont, N.; Austen, M.; Atkins, J.; Burdon, D.; Degraer, S.; Dentinho, T.; Derous, S.; Holm, P.; Horton, T.; Van Ierland, E. Identification, definition and quantification of goods and services provided by marine biodiversity: Implications for the ecosystem approach. Mar. Pollut. Bull. 2007, 54, 253–265. [Google Scholar] [CrossRef]
- Pauli, H.; Gottfried, M.; Dullinger, S.; Abdaladze, O.; Akhalkatsi, M.; Alonso, J.L.B.; Coldea, G.; Dick, J.; Erschbamer, B.; Calzado, R.F. Recent plant diversity changes on Europe’s mountain summits. Science 2012, 336, 353–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flagmeier, M.; Long, D.G.; Genney, D.R.; Hollingsworth, P.M.; Ross, L.C.; Woodin, S.J. Fifty years of vegetation change in oceanic-montane liverwort-rich heath in Scotland. Plant Ecol. Divers. 2014, 7, 457–470. [Google Scholar] [CrossRef]
- Sproull, G.J.; Quigley, M.F.; Sher, A.; González, E. Long-term changes in composition, diversity and distribution patterns in four herbaceous plant communities along an elevational gradient. J. Veg. Sci. 2015, 26, 552–563. [Google Scholar] [CrossRef]
- Christmas, M.J.; Breed, M.F.; Lowe, A.J. Constraints to and conservation implications for climate change adaptation in plants. Conserv. Genet. 2016, 17, 305–320. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.L.; Yao, L.J.; Meng, J.S.; Tao, J. Maxent modeling for predicting the potential geographical distribution of two peony species under climate change. Sci. Total Environ. 2018, 634, 1326–1334. [Google Scholar] [CrossRef]
- Hu, H.W.; Wei, Y.Q.; Wang, W.Y.; Wang, C.Y. The Influence of climate change on three dominant alpine species under different scenarios on the Qinghai–Tibetan Plateau. Diversity 2021, 13, 682. [Google Scholar] [CrossRef]
- Wei, Y.Q.; Zhang, L.; Wang, J.N.; Wang, W.W.; Niyati, N.; Guo, Y.L.; Wang, X.F. Chinese caterpillar fungus (Ophiocordyceps sinensis) in China: Current distribution, trading, and futures under climate change and overexploitation. Sci. Total Environ. 2021, 755, 142548. [Google Scholar] [CrossRef]
- Chen, I.-C.; Hill, J.K.; Ohlemüller, R.; Roy, D.B.; Thomas, C.D. Rapid range shifts of species associated with high levels of climate warming. Science 2011, 333, 1024–1026. [Google Scholar] [CrossRef]
- Xiaodan, W.; Genwei, C.; Xianghao, Z. Assessing potential impacts of climatic change on subalpine forests on the eastern Tibetan Plateau. Clim. Chang. 2011, 108, 225–241. [Google Scholar] [CrossRef]
- He, X.; Burgess, K.S.; Yang, X.F.; Ahrends, A.; Gao, L.M.; Li, D.Z. Upward elevation and northwest range shifts for alpine Meconopsis species in the Himalaya–Hengduan Mountains region. Ecol. Evol. 2019, 9, 4055–4064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Chen, T.; Xu, G.; Liu, X.; Wang, W.; Wu, G.; Zhang, Y. Alpine timberline population dynamics under climate change: A comparison between Qilian juniper and Qinghai spruce tree species in the middle Qilian Mountains of northeast Tibetan Plateau. Boreas 2016, 45, 411–422. [Google Scholar] [CrossRef]
- Bugmann, H. A review of forest gap models. Clim. Chang. 2001, 51, 259–305. [Google Scholar] [CrossRef]
- Hörsch, B. Modelling the spatial distribution of montane and subalpine forests in the central Alps using digital elevation models. Ecol. Model. 2003, 168, 267–282. [Google Scholar] [CrossRef]
- Chauchard, S.; Carcaillet, C.; Guibal, F. Patterns of land-use abandonment control tree-recruitment and forest dynamics in Mediterranean mountains. Ecosystems 2007, 10, 936–948. [Google Scholar] [CrossRef]
- Guisan, A.; Theurillat, J.-P. Assessing alpine plant vulnerability to climate change: A modeling perspective. Integr. Assess. 2000, 1, 307–320. [Google Scholar] [CrossRef]
- Halloy, S.R.; Mark, A.F. Climate-change effects on alpine plant biodiversity: A New Zealand perspective on quantifying the threat. Arct. Antarct. Alp. Res. 2003, 35, 248–254. [Google Scholar] [CrossRef]
- Shi, N.; Naudiyal, N.; Wang, J.N.; Gaire, N.P.; Wu, Y.; Wei, Y.Q.; He, J.L.; Wang, C.Y. Assessing the impact of climate change on potential distribution of Meconopsis punicea and its influence on ecosystem services supply in the southeastern margin of Qinghai-Tibet Plateau. Front. Plant Sci. 2022, 12, 3338. [Google Scholar] [CrossRef]
- Zhang, Y.; Shao, X.; Wilmking, M. Dynamic relationships between Picea crassifolia growth and climate at upper treeline in the Qilian Mts., Northeast Tibetan Plateau, China. Dendrochronologia 2011, 29, 185–199. [Google Scholar] [CrossRef]
- Singh, S.; Sharma, S.; Dhyani, P. Himalayan arc and treeline: Distribution, climate change responses and ecosystem properties. Biodivers. Conserv. 2019, 28, 1997–2016. [Google Scholar] [CrossRef]
- Dorji, T.; Hopping, K.A.; Meng, F.; Wang, S.; Jiang, L.; Klein, J.A. Impacts of climate change on flowering phenology and production in alpine plants: The importance of end of flowering. Agric. Ecosyst. Environ. 2020, 291, 106795. [Google Scholar] [CrossRef]
- Yang, F.S.; Qin, A.L.; Li, Y.F.; Wang, X.Q. Great genetic differentiation among populations of Meconopsis integrifolia and its implication for plant speciation in the Qinghai-Tibetan Plateau. PLoS ONE 2012, 7, e37196. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.F.; Wu, Z.J.; Zhu, H.J.; Peng, S.L.; Zhang, X.F. Chemical constituents of Meconopsis punicea. Nat. Prod. Res. Dev. 2011, 23, 202–207. [Google Scholar]
- Wang, W.T.; Guo, W.Y.; Jarvie, S.; Svenning, J.C. The fate of Meconopsis species in the Tibeto-Himalayan region under future climate change. Ecol. Evol. 2021, 11, 887–899. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Wang, D.; Miao, X.; Wang, Y.; Zhang, J.; Wang, X.; Zhang, Y.; Pan, H. Antinociceptive and anti-tussive activities of the ethanol extract of the flowers of Meconopsis punicea Maxim. BMC Complement. Altern. Med. 2015, 15, 154. [Google Scholar] [CrossRef] [Green Version]
- Yasuhara, M.; Hunt, G.; Breitburg, D.; Tsujimoto, A.; Katsuki, K. Human-induced marine ecological degradation: Micropaleontological perspectives. Ecol. Evol. 2012, 2, 3242–3268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kearney, M.R.; Wintle, B.A.; Porter, W.P. Correlative and mechanistic models of species distribution provide congruent forecasts under climate change. Conserv. Lett. 2010, 3, 203–213. [Google Scholar] [CrossRef]
- Baker, D.J.; Hartley, A.J.; Butchart, S.H.; Willis, S.G. Choice of baseline climate data impacts projected species’ responses to climate change. Glob. Chang. Biol. 2016, 22, 2392–2404. [Google Scholar] [CrossRef] [Green Version]
- Pouteau, R.; Meyer, J.-Y.; Taputuarai, R.; Stoll, B. Support vector machines to map rare and endangered native plants in Pacific islands forests. Ecol. Inform. 2012, 9, 37–46. [Google Scholar] [CrossRef]
- Ahn, Y.; Lee, D.-K.; Kim, H.G.; Park, C.; Kim, J.; Kim, J.-U. Estimating Korean Pine (Pinus koraiensis) habitat distribution considering climate change uncertainty-using species distribution models and RCP Scenarios. J. Korean Soc. Environ. Restor. Technol. 2015, 18, 51–64. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.G.; Lee, D.K.; Ryu, J.E. Riparian connectivity assessment using species distribution model of fish assembly. J. Korean Soc. Geospat. Inf. Sci. 2015, 23, 17–26. [Google Scholar]
- Wellmann, T.; Lausch, A.; Scheuer, S.; Haase, D. Earth observation based indication for avian species distribution models using the spectral trait concept and machine learning in an urban setting. Ecol. Indic. 2020, 111, 106029. [Google Scholar] [CrossRef]
- Yan, H.Y.; Feng, L.; Zhao, Y.F.; Feng, L.; Zhu, C.P.; Qu, Y.F.; Wang, H.Q. Predicting the potential distribution of an invasive species, Erigeron canadensis L. in China with a maximum entropy model. Glob. Ecol. Conserv. 2020, 21, e00822. [Google Scholar] [CrossRef]
- Koo, K.A.; Park, S.U.; Kong, W.-S.; Hong, S.; Jang, I.; Seo, C. Potential climate change effects on tree distributions in the Korean Peninsula: Understanding model & climate uncertainties. Ecol. Model. 2017, 353, 17–27. [Google Scholar]
- Li, X.; Wang, Y. Applying various algorithms for species distribution modelling. Integr. Zool. 2013, 8, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Brun, P.; Thuiller, W.; Chauvier, Y.; Pellissier, L.; Wüest, R.O.; Wang, Z.; Zimmermann, N.E. Model complexity affects species distribution projections under climate change. J. Biogeogr. 2020, 47, 130–142. [Google Scholar] [CrossRef]
- Paź-Dyderska, S.; Jagodziński, A.M.; Dyderski, M.K. Possible changes in spatial distribution of walnut (Juglans regia L.) in Europe under warming climate. Reg. Environ. Chang. 2021, 21, 18. [Google Scholar] [CrossRef]
- Olszewski, P.; Dyderski, M.K.; Dylewski, Ł.; Bogusch, P.; Schmid-Egger, C.; Ljubomirov, T.; Zimmermann, D.; Le Divelec, R.; Wiśniowski, B.; Twerd, L. European beewolf (Philanthus triangulum) will expand its geographic range as a result of climate warming. Reg. Environ. Chang. 2022, 22, 129. [Google Scholar] [CrossRef]
- Riahi, K.; Van Vuuren, D.P.; Kriegler, E.; Edmonds, J.; O’neill, B.C.; Fujimori, S.; Bauer, N.; Calvin, K.; Dellink, R.; Fricko, O. The Shared Socioeconomic Pathways and their energy, land use, and greenhouse gas emissions implications: An overview. Glob. Environ. Chang. 2017, 42, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Beaumont, L.J.; Hughes, L.; Poulsen, M. Predicting species distributions: Use of climatic parameters in BIOCLIM and its impact on predictions of species’ current and future distributions. Ecol. Model. 2005, 186, 251–270. [Google Scholar] [CrossRef]
- Molina-Montenegro, M.A.; Atala, C.; Gianoli, E. Phenotypic plasticity and performance of Taraxacum officinale (dandelion) in habitats of contrasting environmental heterogeneity. Biol. Invasions 2010, 12, 2277–2284. [Google Scholar] [CrossRef]
- Ganjurjav, H.; Gornish, E.S.; Hu, G.Z.; Schwartz, M.W.; Wan, Y.F.; Li, Y.; Gao, Q.Z. Warming and precipitation addition interact to affect plant spring phenology in alpine meadows on the central Qinghai-Tibetan Plateau. Agric. For. Meteorol. 2020, 287, 107943. [Google Scholar] [CrossRef]
- Iwasa, Y.; Cohen, D. Optimal growth schedule of a perennial plant. Am. Nat. 1989, 133, 480–505. [Google Scholar] [CrossRef]
- Rohde, A.; Bhalerao, R.P. Plant dormancy in the perennial context. Trends Plant Sci. 2007, 12, 217–223. [Google Scholar] [CrossRef]
- Fadón, E.; Rodrigo, J. Unveiling winter dormancy through empirical experiments. Environ. Exp. Bot. 2018, 152, 28–36. [Google Scholar] [CrossRef]
- Zhao, W.; Chen, H.; Liu, L.; Qi, M.; Zhang, J.; Du, T.; Jin, L. The impact of climate change on the distribution pattern of the suitable growing region for endangered Tibetan medicine Meconopsis punicea. Chin. Pharm. J. 2021, 56, 1306–1312. [Google Scholar]
- Xin, X.G.; Wu, T.W.; Zhang, J.; Zhang, F.; Li, W.P.; Zhang, Y.W.; Lu, Y.X.; Fang, Y.J.; Jie, W.H.; Zhang, L.; et al. Introduction of BCC models and its participation in CMIP6. Adv. Clim. Chang. Res. 2019, 15, 533. [Google Scholar]
- Gao, T.; Xu, Q.; Liu, Y.; Zhao, J.Q.; Shi, J. Predicting the potential geographic distribution of Sirex nitobei in China under climate change using maximum entropy model. Forests 2021, 12, 151. [Google Scholar] [CrossRef]
- Arias, P.; Bellouin, N.; Coppola, E.; Jones, R.; Krinner, G.; Marotzke, J.; Naik, V.; Palmer, M.; Plattner, G.-K.; Rogelj, J. Technical Summary. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2021; pp. 33–144. [Google Scholar]
- Wiens, J.A.; Stralberg, D.; Jongsomjit, D.; Howell, C.A.; Snyder, M.A. Niches, models, and climate change: Assessing the assumptions and uncertainties. Proc. Natl. Acad. Sci. USA 2009, 106, 19729–19736. [Google Scholar] [CrossRef] [Green Version]
- Ackerly, D.; Loarie, S.; Cornwell, W.; Weiss, S.; Hamilton, H.; Branciforte, R.; Kraft, N. The geography of climate change: Implications for conservation biogeography. Divers. Distrib. 2010, 16, 476–487. [Google Scholar] [CrossRef]
- Wu, Y.; Liu, Y.; Peng, H.; Yang, Y.; Liu, G.; Cao, G.; Zhang, Q. Pollination ecology of alpine herb Meconopsis integrifolia at different altitudes. Chin. J. Plant Ecol. 2015, 39, 1–13. [Google Scholar]
- Roff, D.A. The evolution of flightlessness in insects. Ecol. Monogr. 1990, 60, 389–421. [Google Scholar] [CrossRef]
- McCulloch, G.A.; Foster, B.J.; Dutoit, L.; Ingram, T.; Hay, E.; Veale, A.J.; Dearden, P.K.; Waters, J.M. Ecological gradients drive insect wing loss and speciation: The role of the alpine treeline. Mol. Ecol. 2019, 28, 3141–3150. [Google Scholar] [CrossRef]
- Kinlan, B.P.; Gaines, S.D. Propagule dispersal in marine and terrestrial environments: A community perspective. Ecology 2003, 84, 2007–2020. [Google Scholar] [CrossRef]
- Thomson, F.J.; Moles, A.T.; Auld, T.D.; Kingsford, R.T. Seed dispersal distance is more strongly correlated with plant height than with seed mass. J. Ecol. 2011, 99, 1299–1307. [Google Scholar] [CrossRef]
- Morgan, J.W.; Venn, S. Alpine plant species have limited capacity for long-distance seed dispersal. Plant Ecol. 2017, 218, 813–819. [Google Scholar] [CrossRef]
- Di Musciano, M.; Di Cecco, V.; Bartolucci, F.; Conti, F.; Frattaroli, A.R.; Di Martino, L. Dispersal ability of threatened species affects future distributions. Plant Ecol. 2020, 221, 265–281. [Google Scholar] [CrossRef]
- Qu, Y.; Qu, Z. The research advancement on the genus Meconpsis. North. Hortic. 2012, 2, 191–194. [Google Scholar]
- Khoury, C.; Laliberté, B.; Guarino, L. Trends in ex situ conservation of plant genetic resources: A review of global crop and regional conservation strategies. Genet. Resour. Crop Evol. 2010, 57, 625–639. [Google Scholar] [CrossRef]
- Niyati, N.; Wang, J.N.; Wu, N.; Narayan Prasad, G.; Shi, P.L.; Wei, Y.Q.; He, J.L.; Shi, N. Potential distribution of Abies, Picea, and Juniperus species in the sub-alpine forest of Minjiang headwater region under current and future climate scenarios and its implications on ecosystem services supply. Ecol. Indic. 2021, 121, 107131. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Kumar, P. Assessment of impact of climate change on Rhododendrons in Sikkim Himalayas using Maxent modelling: Limitations and challenges. Biodivers. Conserv. 2012, 21, 1251–1266. [Google Scholar] [CrossRef]
- Moss, R.H.; Edmonds, J.A.; Hibbard, K.A.; Manning, M.R.; Rose, S.K.; Van Vuuren, D.P.; Carter, T.R.; Emori, S.; Kainuma, M.; Kram, T. The next generation of scenarios for climate change research and assessment. Nature 2010, 463, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Van Vuuren, D.P.; Edmonds, J.; Kainuma, M.; Riahi, K.; Thomson, A.; Hibbard, K.; Hurtt, G.C.; Kram, T.; Krey, V.; Lamarque, J.-F. The representative concentration pathways: An overview. Clim. Chang. 2011, 109, 5–31. [Google Scholar] [CrossRef]
- Fan, X.W.; Miao, C.Y.; Duan, Q.Y.; Shen, C.W.; Wu, Y. The performance of CMIP6 versus CMIP5 in simulating temperature extremes over the global land surface. J. Geophys. Res. Atmos. 2020, 125, e2020JD033031. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Hilden, J. The area under the ROC curve and its competitors. Med. Decis. Mak. 1991, 11, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [Green Version]
- Manel, S.; Williams, H.C.; Ormerod, S.J. Evaluating presence-absence models in ecology: The need to account for prevalence. J. Appl. Ecol. 2001, 38, 921–931. [Google Scholar] [CrossRef]
- Cohen, J. A coefficient of agreement for nominal scales. Educ. Psychol. Meas. 1960, 20, 37–46. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P.; Liu, C. Modelling species distributions in Britain: A hierarchical integration of climate and land-cover data. Ecography 2004, 27, 285–298. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting thresholds of occurrence in the prediction of species distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Freeman, E.A.; Moisen, G.G. A comparison of the performance of threshold criteria for binary classification in terms of predicted prevalence and kappa. Ecol. Model. 2008, 217, 48–58. [Google Scholar] [CrossRef]
- Guisan, A.; Theurillat, J.P.; Kienast, F. Predicting the potential distribution of plant species in an alpine environment. J. Veg. Sci. 1998, 9, 65–74. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P.; Berry, P.M.; Harrison, P. SPECIES: A spatial evaluation of climate impact on the envelope of species. Ecol. Model. 2002, 154, 289–300. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GLM | GBM | RF | FDA | |
|---|---|---|---|---|
| BIO2 | 0.1502 | 0.0452 | 0.0198 | 0.8284 |
| BIO3 | 0.1276 | 0.0254 | 0.0194 | 0.6464 |
| BIO4 | 0.9842 | 0.0092 | 0.0468 | 0.9972 |
| BIO5 | 0.5024 | 0.0282 | 0.0398 | 0.0620 |
| BIO11 | 0.6760 | 0.1458 | 0.0742 | 0.0096 |
| BIO15 | 0.48186 | 0.2244 | 0.1282 | 0.1318 |
| BIO17 | 0.4210 | 0.0084 | 0.0244 | 0.1826 |
| BIO18 | 0.7922 | 0.6958 | 0.3162 | 0.3958 |
| Bioclimatic Variables | Meaning of Variables |
|---|---|
| BIO2 | Mean Diurnal Range (mean of monthly (max temp-min temp))/°C |
| BIO3 | Isothermality ((BIO2/BIO7) × 100) |
| BIO4 | Temperature Seasonality (standard deviation ×100) |
| BIO5 | Max Temperature of Warmest Month/°C |
| BIO11 | Mean Temperature of Coldest Quarter/°C |
| BIO15 | Precipitation Seasonality (coefficient of variation) |
| BIO17 | Precipitation of Driest Quarter/mm |
| BIO18 | Precipitation of Warmest Quarter/mm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.-T.; Wang, W.-T. Prediction of the Potential Distribution of the Endangered Species Meconopsis punicea Maxim under Future Climate Change Based on Four Species Distribution Models. Plants 2023, 12, 1376. https://doi.org/10.3390/plants12061376
Zhang H-T, Wang W-T. Prediction of the Potential Distribution of the Endangered Species Meconopsis punicea Maxim under Future Climate Change Based on Four Species Distribution Models. Plants. 2023; 12(6):1376. https://doi.org/10.3390/plants12061376
Chicago/Turabian StyleZhang, Hao-Tian, and Wen-Ting Wang. 2023. "Prediction of the Potential Distribution of the Endangered Species Meconopsis punicea Maxim under Future Climate Change Based on Four Species Distribution Models" Plants 12, no. 6: 1376. https://doi.org/10.3390/plants12061376