
Restoration of Vegetation Greenness and Possible Changes in Mature Forest Communities in Two Forests Damaged by the Vaia Storm in Northern Italy

Abstract
:1. Introduction
2. Materials and Methods
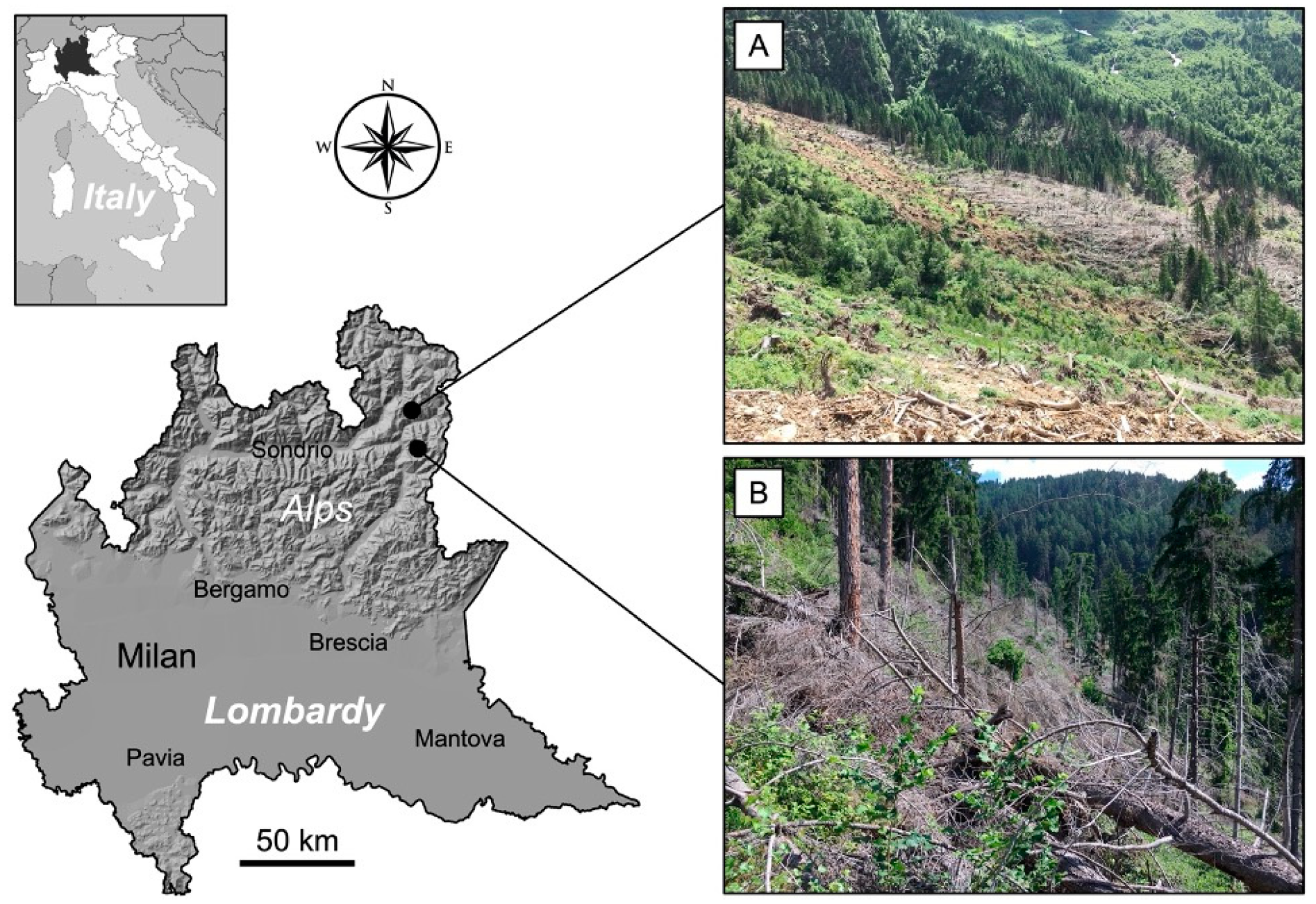
2.1. Study Areas
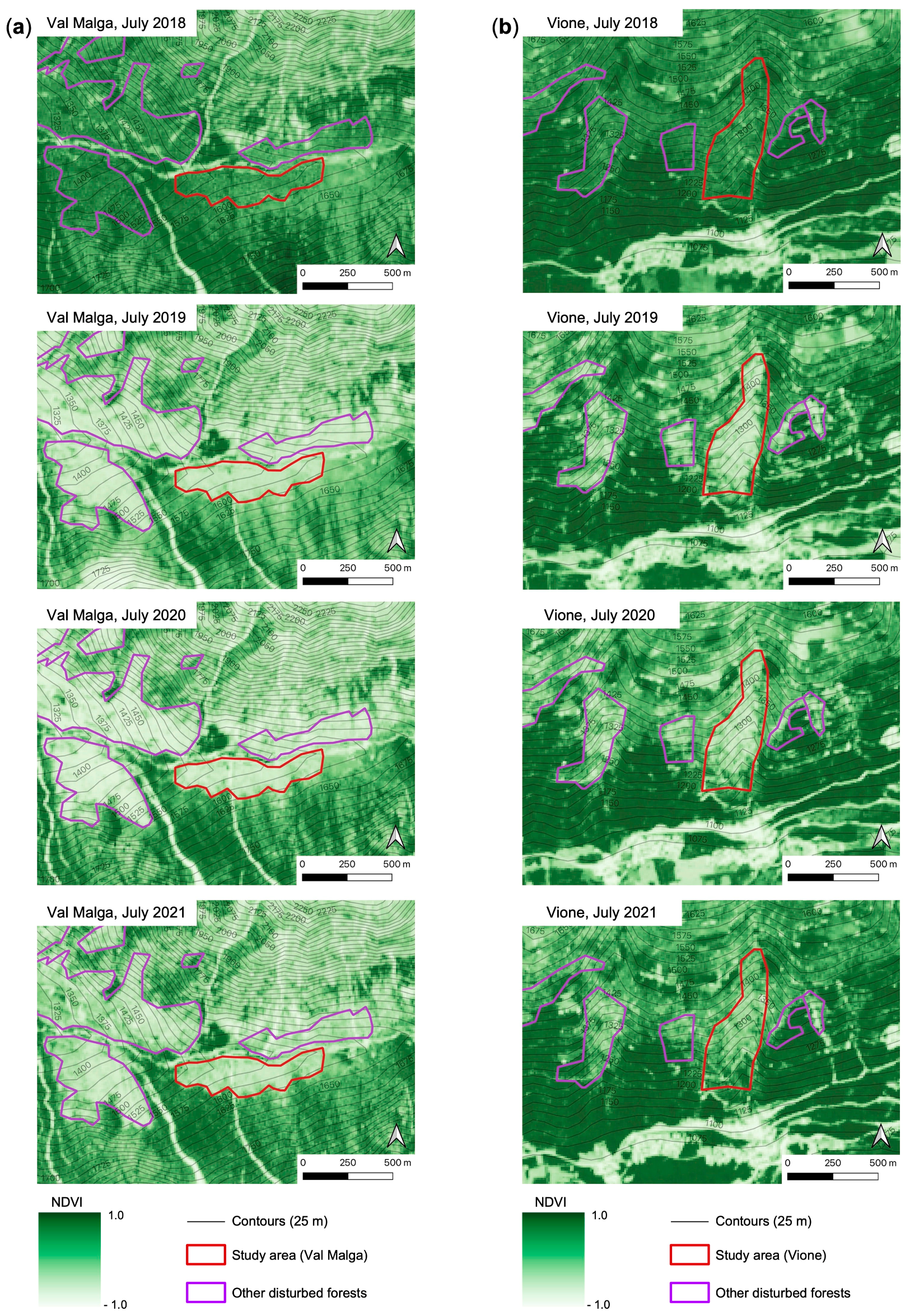
2.2. Multispectral Images and NDVI Analysis
2.3. Vegetation Sampling and Data Analysis
3. Results
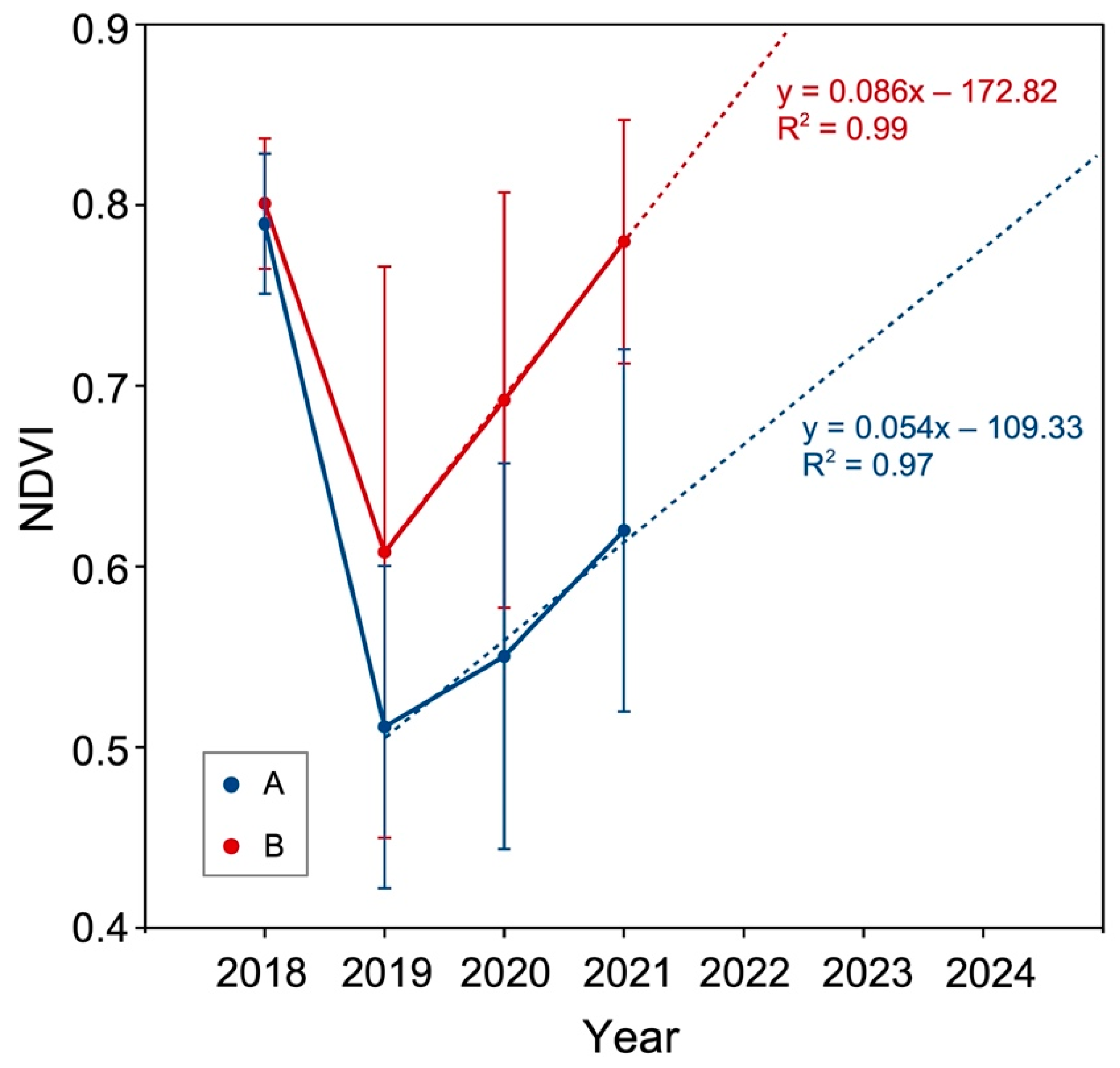
3.1. NDVI Values and Trends
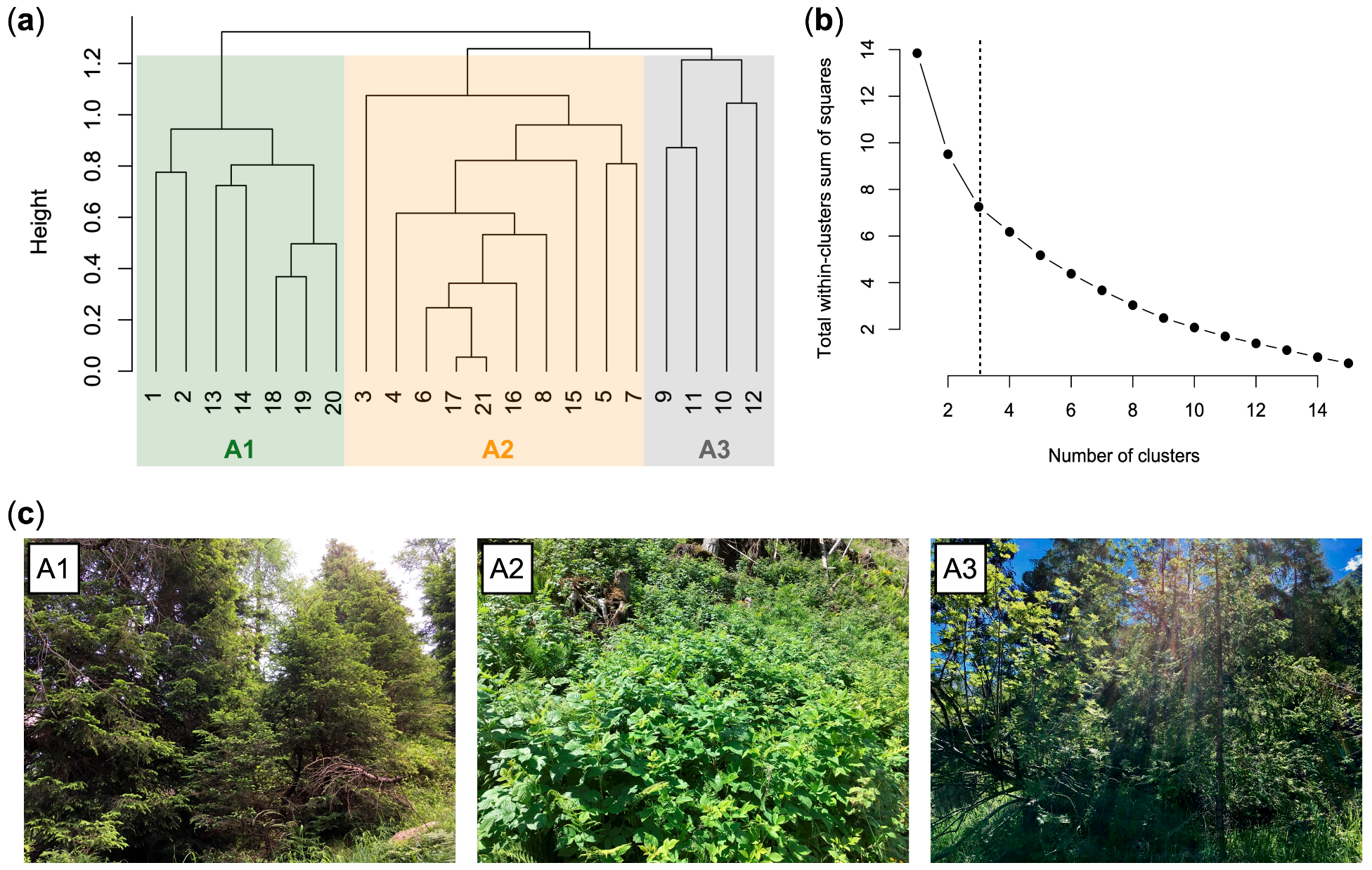
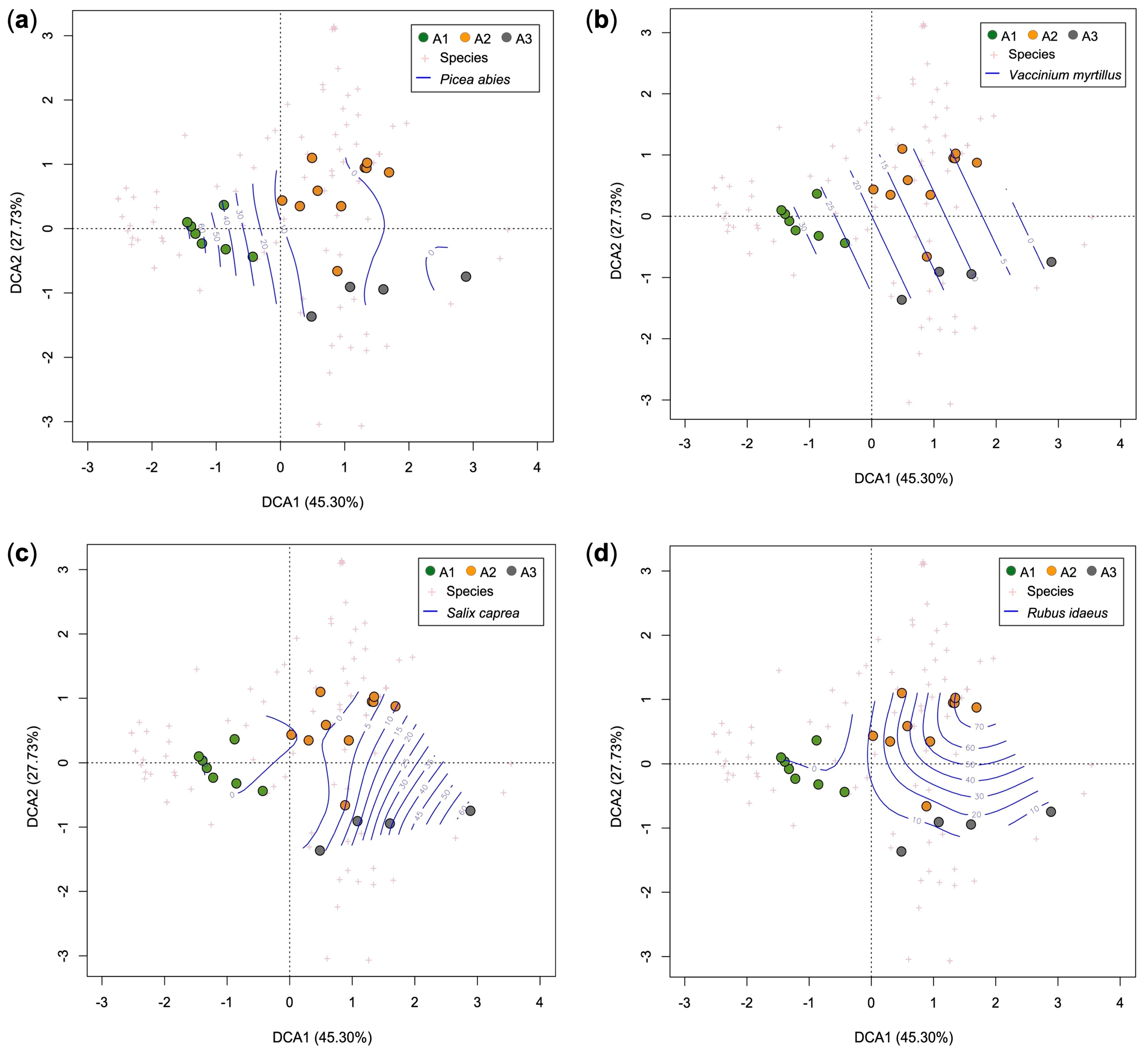
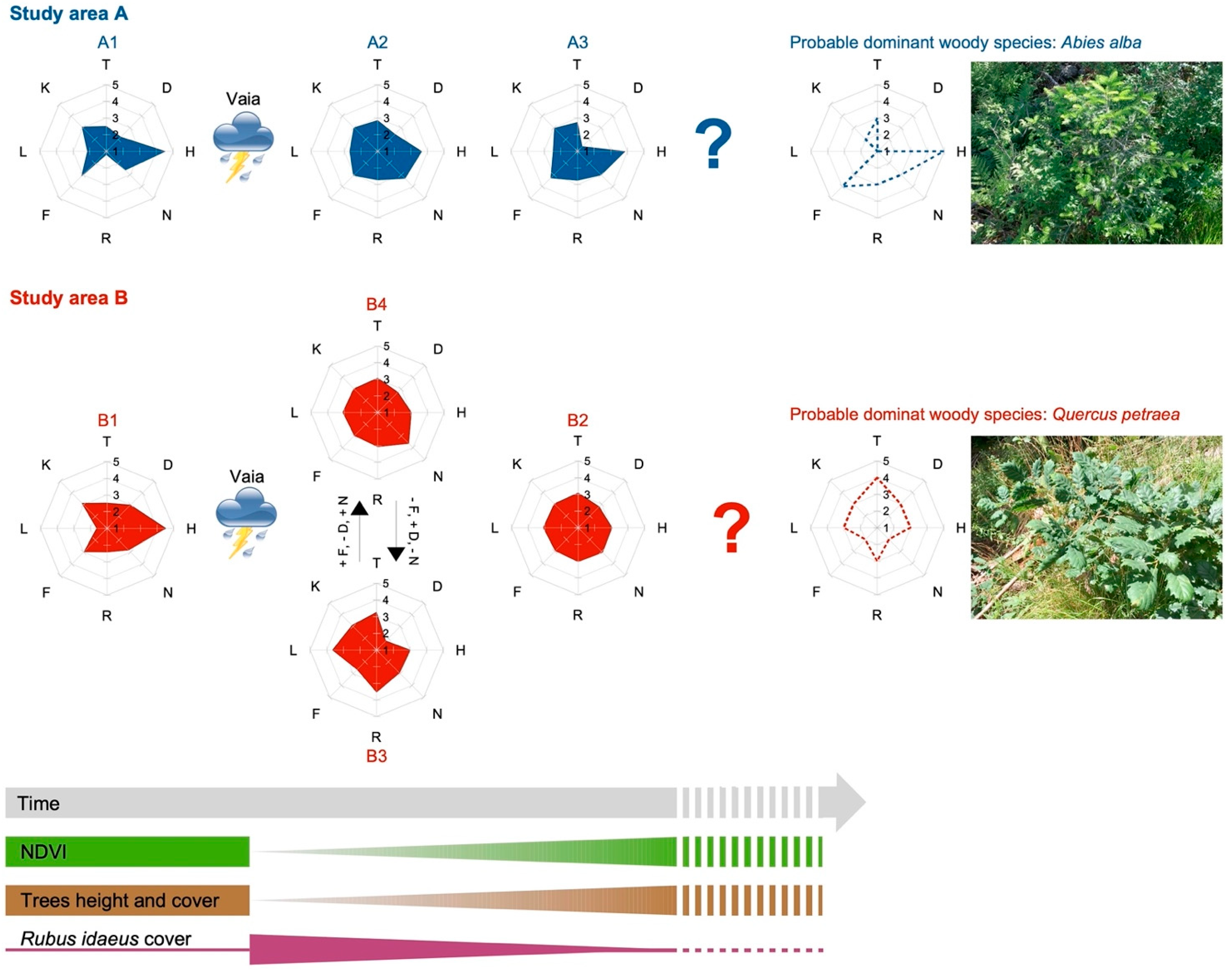
3.2. Current Vegetation of Area A
- A1, Picea abies forest (Calamagrostio arundinaceae-Piceetum): vegetation present at the edges of the forest destroyed by the Vaia storm (control), consisting of spruce (Picea abies), which is the dominant tree, and other diagnostic species of the Vaccinio-Piceetea class, (Table 1) including Larix decidua, Vaccinium myrtillus, Arctostaphylos uva-ursi, and Oxalis acetosella.
- A2, Rubetum idaei: shrubland dominated by Rubus idaeus, which is a diagnostic species of this cluster together with Luzula sylvatica (Table 1). In this community, there are also herbaceous species of forest edges (such as Epilobium angustifolium) and young woody plants such as Sorbus aucuparia, Sambucus racemosa, Salix caprea, and Abies alba (silver fir). Juvenile spruces are scarce. Rubetum idaei is a secondary vegetation type of clearing, forests opening, and degraded forest. The height of this vegetation is generally less than 1.5 m.
- A3, Piceo abietis-Sorbetum aucupariae: community with Sorbus aucuparia (growing mostly as a shrub) accompanied by other woody plants and shrubs of the Sambuco-Salicion capreae alliance (Robinietea class) as Salix caprea (diagnostic species of this cluster), Sambucus racemosa, Betula pendula, and Rubus idaeus. Calamagrostis villosa and Abies alba, two species that characterize the silver fir woods of Lombardy (Calamagrostio villosae-Abietetum), also grow in this plant community. This is a short-living community occurring on forest clearings, where it is gradually replaced by the vegetation of mature forests. Vegetation height: 1–3.5 m.
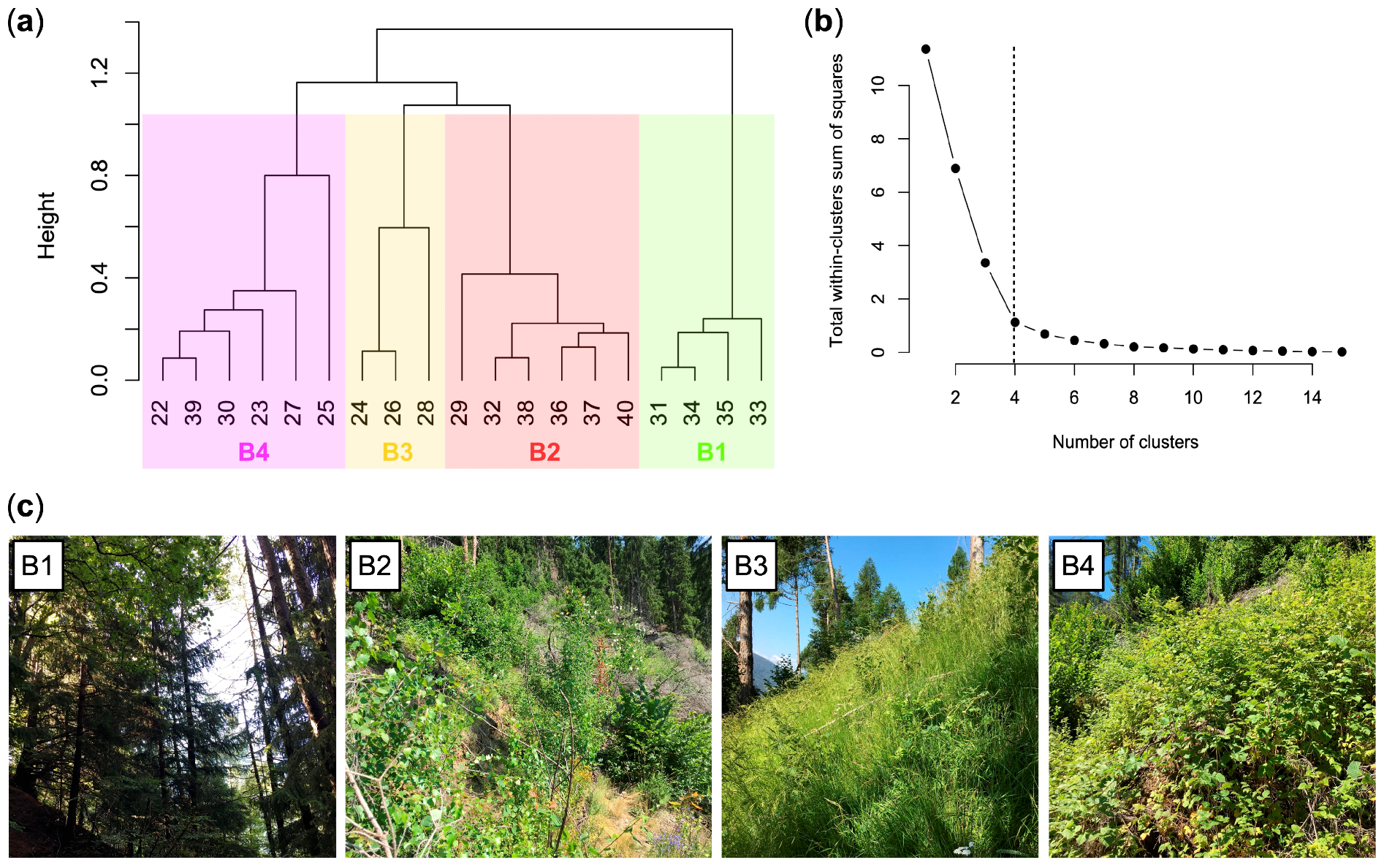
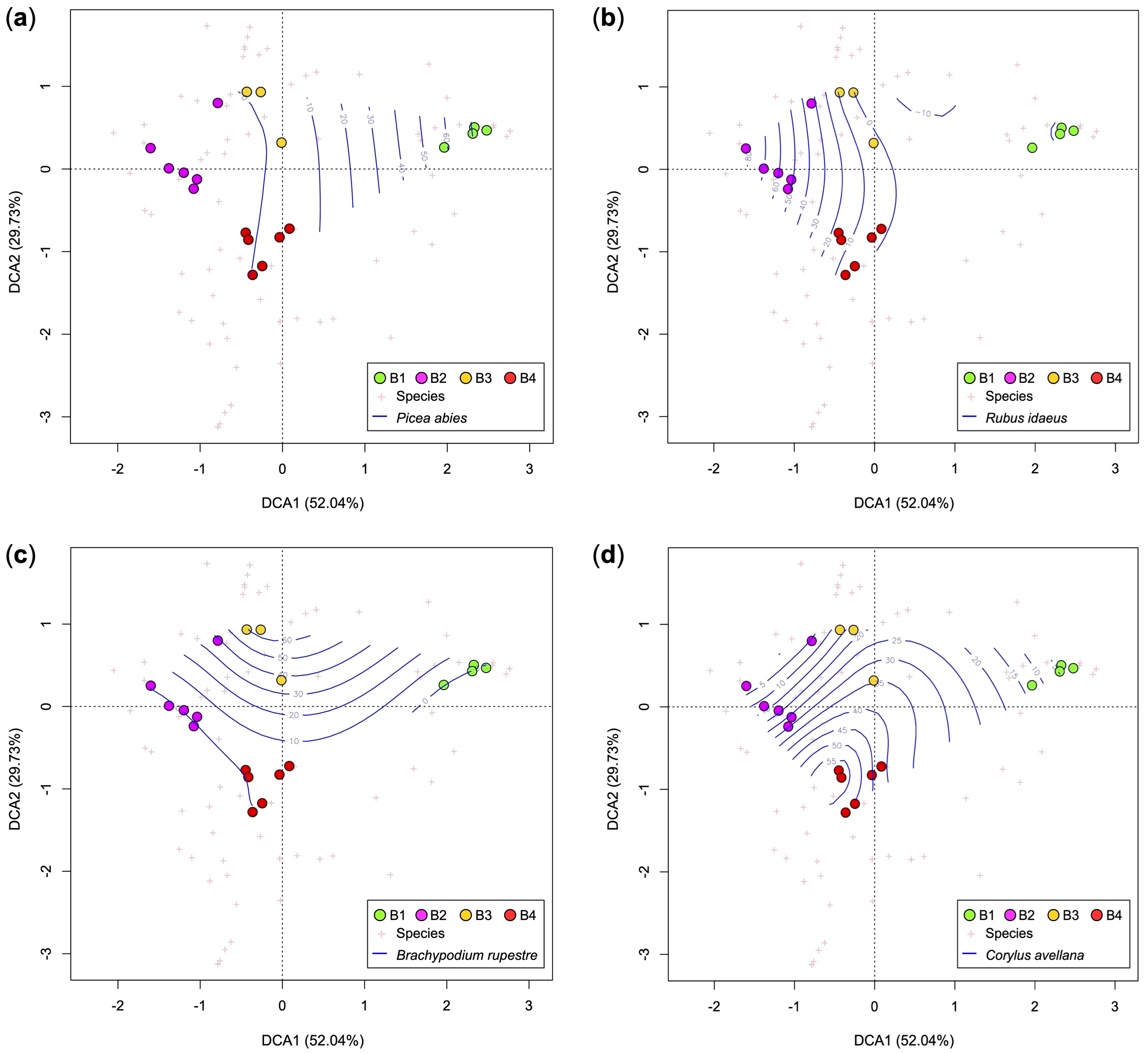
3.3. Current Vegetation of Area B
- B1, Picea abies forest (Calamagrostio arundinaceae-Piceetum): this vegetation is physiognomically similar to A1 but with some species of the mountain-hilly belt, typical of broad-leaved woods, such as Quercus petraea (sessile oak), Luzula nivea, Veronica officinalis, Campanula rotundifolia, Corylus avellana, and Prunus avium (Table 1 and Table S2). This is the vegetation present at the edges of the forest destroyed by Vaia (control).
- B2, community of Astrantio-Corylion avellanae alliance: shrubland with species of the Crataego-Prunetea class such as Corylus avellana (diagnostic species of this cluster), Rosa canina, and Prunus spinosa, and other pioneer shrubs and trees such as Populus tremula, Salix caprea, Buddleja davidii (exotic species), Betula pendula, and Sambucus nigra. Spruce is scarce while Quercus petraea is fairly abundant. The height of the trees/shrubs is about 1.5–3.0 m.
- B3, Brachypodium rupestre-Astragalus glycyphyllos community: grassland dominated by Bachypodium rupestre with other herbaceous diagnostic species of this cluster (Achillea millefolium, Lotus corniculatus, Astragalus glycyphyllos and Festuca valesiaca) and young shrubs (Corylus avellana, Rosa canina, Rubus idaeus, Salix caprea and Prunus avium). In this community, there are larches over 10 m high that survived Vaia.
- B4, Rubetum idaei: plant community dominated by Rubus idaeus, unique diagnostic species of this cluster (Table 1). In this plant community, there are also young shrubs/trees (maximum plant height: 1.5 m) such as Corylus avellana, Salix caprea, Betula pendula, Fraxinus excelsior, and Quercus petraea. No young/juvenile spruce was detected.
3.4. Plant Succession Models of the Two Study Areas
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Usbeck, T.; Wohlgemuth, T.; Dobbertin, M.; Pfister, C.; Bürgi, A.; Rebetez, M. Increasing storm damage to forests in Switzerland from 1858 to 2007. Agric. For. Meteorol. 2010, 150, 47–55. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.J.; Rammer, W.; Verkerk, P.J. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Change 2014, 4, 806–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saad, C.; Boulanger, Y.; Beaudet, M.; Gachon, P.; Ruel, J.C.; Gauthier, S. Potential impact of climate change on the risk of windthrow in eastern Canada’s forests. Clim. Change 2017, 143, 487–501. [Google Scholar] [CrossRef]
- Motta, R.; Ascoli, D.; Corona, P.; Marchetti, M.; Vacchiano, G. Selvicoltura e schianti da vento. Il caso della “tempesta Vaia”. Forest@ 2018, 15, 94–98. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.; Allen, C.D.; Anderson-Teixeira, K.; Aukema, B.H.; Bond-Lamberty, B.; Chini, L.; Clark, J.S.; Dietze, M.; Grossiord, C.; Hanbury-Brown, A.; et al. Pervasive shifts in forest dynamics in a changing world. Science 2020, 368, 964. [Google Scholar] [CrossRef]
- Gardiner, B.; Blennow, K.; Carnus, J.M.; Fleischer, P.; Ingemarsson, F.; Landmann, G.; Lindner, M.; Marzano, M.; Nicoll, B.; Orazio, C.; et al. Destructive Storms in European Forests: Past and Forthcoming Impacts; European Forest Institute, Efiatlantic: Joensuu, Finland, 2010. [Google Scholar]
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliott, K.; Ford, C.R.; Foster, D.R.; Kloeppel, B.D.; Knoepp, J.D.; Lovett, G.M.; et al. Loss of foundation species: Consequences for the structure and dynamics of forested ecosystems. Front. Ecol. Environ. 2005, 3, 479–486. [Google Scholar] [CrossRef]
- Duelli, P.; Wermelinger, B.; Moretti, M.; Obrist, M. Fire and windthrow in forests: Winners and losers in Neuropterida and Mecoptera. Alp. Entomol. 2019, 3, 39–50. [Google Scholar] [CrossRef]
- Giupponi, L.; Bischetti, G.B.; Giorgi, A. Vegetation analysis and estimation of forest reconstitution time in protected areas of Val Camonica (Southern Alps) where a commercial mixture of seeds was sown. J. Prot. Mt. Areas Res. Manag. 2017, 9, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Bordon, N.G.; Nogueira, A.; Filho, N.L.; Higuchi, N. Blowdown disturbance effect on the density, richness and species composition of the seed bank in Central Amazonia. For. Ecol. Manag. 2019, 453, 117633. [Google Scholar] [CrossRef]
- Walker, L.R.; del Moral, R. Lessons from primary succession for restoration of severely damaged habitats. Appl. Veg. Sci. 2008, 12, 56–67. [Google Scholar] [CrossRef]
- Giupponi, L.; Borgonovo, G.; Giorgi, A.; Bischetti, G.B. How to renew soil bioengineering for slope stabilization: Some proposals. Landsc. Ecol. Eng. 2019, 15, 37–50. [Google Scholar] [CrossRef]
- Gardiner, B.; Schuck, A.; Schelhaas, M.-J.; Orazio, C.; Blennow, K.; Nicoli, B. Living with Storm Damage to Forests. What Science Can Tell Us 3; European Forest Institute: Joensuu, Finland, 2013. [Google Scholar]
- Chirici, G.; Giannetti, F.; Travaglini, D.; Nocentini, S.; Francini, S.; D’Amico, G.; Calvo, E.; Fasolini, D.; Broll, M.; Maistrelli, F.; et al. Stima dei danni della tempesta “Vaia” alle foreste in Italia. Forest@ 2019, 16, 3–9. [Google Scholar] [CrossRef] [Green Version]
- Laurin, G.V.; Francini, S.; Luti, T.; Chirici, G.; Pirotti, F.; Papale, D. Satellite open data to monitor forest damage caused by extreme climate-induced events: A case study of the Vaia storm in Northern Italy. Forestry 2021, 94, 407–416. [Google Scholar] [CrossRef]
- Rapella, A. La Tempesta Vaia e le Risorse Messe in Campo in Lombardia. Available online: https://eventi.regione.lombardia.it/it/la-foresta-ritorner (accessed on 7 March 2023).
- Andreis, A.; Armiraglio, S.; Caccianiga, M.; Cerabolini, B. La vegetazione forestale dell’ordine Piceetalia excelsae Pawl. in Pawl. et al. 1928 nelle Alpi Lombarde. Plant Sociol. 2009, 46, 49–74. [Google Scholar]
- Schönenberger, W. Post windthrow stand regeneration in Swiss mountain forests: The first ten years after the 1990 storm Vivian. For. Snow Landsc. Res. 2002, 77, 61–80. [Google Scholar]
- Wohlgemuth, T.; Kull, P.; Wüthrich, H. Disturbance of microsites and early tree regeneration after windthrow in Swiss mountain forests due to the winter storm Vivian 1990. For. Snow Landsc. Res. 2002, 77, 17–47. [Google Scholar]
- Mitchell, S.J. Wind as a natural disturbance agent in forests: A synthesis. Forestry 2012, 86, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Ducoli, A. Interventi di Bonifica in Soprassuoli Colpiti Dalla Tempesta Vaia in Valle Camonica. Report Finale: Primi Risultati e Proposte Programmatiche; Comunità Montana di Valle Camonica: Breno, Italy, 2019. [Google Scholar]
- Blasi, C.; Capotorti, G.; Copiz, R.; Guida, D.; Mollo, B.; Smiraglia, D.; Zavattero, L. Classification and mapping of the ecoregions of Italy. Plant Biosyst. 2014, 148, 1255–1345. [Google Scholar] [CrossRef]
- Verde, S.; Assini, S.; Andreis, C. Le serie di vegetazione della regione Lombardia. In La vegetazione d’Italia; Blasi, C., Ed.; Palombi and Partner S.r.l.: Rome, Italy, 2010; pp. 181–203. [Google Scholar]
- Del Favero, R. I Tipi Forestali Della Lombardia; Cierre: Verona, Italy, 2002. [Google Scholar]
- Del Favero, R. I Boschi Delle Regioni Alpine Italiane; Cleup: Padova, Italy, 2004. [Google Scholar]
- Braun-Blanquet, J. Pflanzensoziologie, 3rd ed.; Springer: Wien, Austria, 1964. [Google Scholar]
- Pignatti, S. Flora d’Italia, 2nd ed.; Edagricole: Bologn, Italy, 2017. [Google Scholar]
- Canullo, R.; Allegrini, M.C.; Campetella, G. Reference field manual for vegetation surveys on the CONECOFOR LII network, Italy (National Programme of Forest Ecosystems Control-UNECE, ICP Forests). Braun-Blanquetia 2012, 48, 5–65. [Google Scholar]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Chytrý, M.; Tichý, L.; Holt, J.; Botta-Dukát, Z. Determination of diagnostic species with statistical fidelity measures. J. Veg. Sci. 2002, 13, 79–90. [Google Scholar] [CrossRef]
- Tichý, L.; Chytrý, M. Statistical determination of diagnostic species for site groups of unequal size. J. Veg. Sci. 2006, 17, 809–818. [Google Scholar] [CrossRef]
- Dixon, P. Vegan, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Wien, Austria, 2021; Available online: http://www.R-project.org/ (accessed on 17 October 2022).
- De Cáceres, M. How to Use Indicspecies Package (Ver. 1.7.1); Centre Tecnologic Forestal de Catalunya: Solsona, Spain, 2013. [Google Scholar]
- Landolt, E.; Bäumler, B.; Erhardt, A.; Hegg, O.; Klötzli, F.; Lämmle, W.; Nobis, M.; Rudmann-Maurer, K.; Schweingruber, F.H.; Theurillat, J.-P.; et al. Flora Indicativa. Ecological Indicator Values and Biological Attributes of the Flora of Switzerland and the Alps; Haupt Verlag: Bern, Switzerland, 2010. [Google Scholar]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; García, R.G.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19 (Suppl. 1), 3–264. [Google Scholar] [CrossRef]
- Ubaldi, D. La Vegetazione Boschiva d’Italia: Manuale di Fitosociologia Forestale; Clueb: Bologna, Italy, 2008. [Google Scholar]
- Biondi, E.; Blasi, C. Prodromo Della Vegetazione d’Italia. 2015. Available online: https://www.prodromo-vegetazione-italia.org (accessed on 5 July 2022).
- Chytrý, M.; Danihelka, J.; Kaplan, Z.; Wild, J.; Holubová, D.; Novotný, P.; Řezníčková, M.; Rohn, M.; Dřevojan, P.; Grulich, V.; et al. Pladias Database of the Czech Flora and Vegetation. Preslia 2021, 93, 1–87. [Google Scholar] [CrossRef]
- Preislerová, Z.; Jiménez-Alfaro, B.; Mucina, L.; Berg, C.; Bonari, G.; Kuzemko, A.; Landucci, F.; Marcenò, C.; Monteiro-Henriques, T.; Novák, P.; et al. Distribution maps of vegetation alliances in Europe. Appl. Veg. Sci. 2022, 25, e12642. [Google Scholar] [CrossRef]
- Suvanto, S.; Peltoniemi, M.; Tuominen, S.; Strandström, M.; Lehtonen, A. High-resolution mapping of forest vulnerability to wind for disturbance-aware forestry. For. Ecol. Manag. 2019, 453, 117619. [Google Scholar] [CrossRef]
- Karaer, A.; Chen, M.; Gazzea, M.; Ghorbanzadeh, M.; Abichou, T.; Arghandeh, R.; Ozguven, E.E. Remote sensing-based comparative damage assessment of historical storms and hurricanes in Northwestern Florida. Int. J. Disaster Risk Reduct. 2022, 72, 102857. [Google Scholar] [CrossRef]
- Ryu, J.-H.; Han, K.-S.; Hong, S.; Park, N.-W.; Lee, Y.-W.; Cho, J. Satellite-Based Evaluation of the Post-Fire Recovery Process from the Worst Forest Fire Case in South Korea. Remote Sens. 2018, 10, 918. [Google Scholar] [CrossRef] [Green Version]
- Reisigl, H.; Keller, C. Alpenpflanzen im lebensraum; Gustav Fischer Verlag: Stuttgart, Germany; New York, NY, USA, 1987. [Google Scholar]
- Körner, C. Alpine Plant Life, 3rd ed.; Springer Nature: Cham, Switzerland, 2021. [Google Scholar]
- Silva, G.F.C.; Goncalves, A.C.A.; Junior, C.A.S.; Nanni, M.R.; Facco, C.U.; Cezar, E.; da Silva, A.A. NDVI Response to Water Stress in Different Phenological Stages in Culture Bean. J. Agron. 2016, 15, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Sarti, M.; Vaccari, F.P.; Calfapietra, C.; Brugnoli, E.; Scartazza, A. A statistical approach to detect land cover changes in mediterranean ecosystems using multi-temporal landsat data: The case study of Pianosa Island, Italy. Forests 2020, 11, 334. [Google Scholar] [CrossRef] [Green Version]
- Barni, E.; Siniscalco, C. Vegetation dynamics and arbuscular mycorrhiza in old-field successions of the western Italian Alps. Mycorrhiza 1999, 10, 63–72. [Google Scholar] [CrossRef]
- Shen, X.; Liu, B.; Henderson, M.; Wang, L.; Jiang, M.; Lu, X. Vegetation Greening, Extended Growing Seasons, and Temperature Feedbacks in Warming Temperate Grasslands of China. J. Clim. 2022, 35, 5103–5117. [Google Scholar] [CrossRef]
- Sun, Z.; Chang, N.-B.; Opp, C.; Hennig, T. Evaluation of ecological restoration through vegetation patterns in the lower Tarim River, China with MODIS NDVI data. Ecol. Inform. 2011, 6, 156–163. [Google Scholar] [CrossRef]
- Rao, Y.; Zhu, X.; Chen, J.; Wang, J. An Improved Method for Producing High Spatial-Resolution NDVI Time Series Datasets with Multi-Temporal MODIS NDVI Data and Landsat TM/ETM+ Images. Remote Sens. 2015, 7, 7865–7891. [Google Scholar] [CrossRef] [Green Version]
- Valtonen, A.; Korkiatupa, E.; Holm, S.; Malinga, G.M.; Nakadai, R. Remotely sensed vegetation greening along a restoration gradient of a tropical forest, Kibale National Park, Uganda. Land. Degrad. Dev. 2021, 32, 5166–5177. [Google Scholar] [CrossRef]
- Giupponi, L.; Bischetti, G.B.; Giorgi, A. A proposal for assessing the success of soil bioengineering work by analysing vegetation: Results of two case studies in the Italian Alps. Landsc. Ecol. Eng. 2017, 13, 305–318. [Google Scholar] [CrossRef]
- Pesaresi, S.; Mancini, A.; Casavecchia, S. Recognition and characterization of forest plant communities through remote-sensing NDVI time series. Diversity 2020, 12, 313. [Google Scholar] [CrossRef]
- Pott, R. Phytosociology: A modern geobotanical method. Plant Biosyst. 2011, 145, 9–18. [Google Scholar] [CrossRef]
- Łaska, G. Ecological consequences of deforestation and afforestation on a post-arable land: Changes in the composition and structure of plant communities and transformations of oak-hornbeam habitats and soil. Ecol. Quest. 2014, 20, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Tampucci, D.; Digiovinazzo, P.; Andreis, C. Serie dinamica del bosco di rovere: Il caso del Parco Nazionale Val Grande (VCO). Nat. Brescia. 2013, 38, 93–99. [Google Scholar]
- Pignatti, S. I boschi d’Italia; UTET: Torino, Italy, 1998. [Google Scholar]
- Lenoir, J.; Gégout, J.C.; Marquet, P.A.; de Ruffray, P.; Brisse, H. A significant upward shift in plant species optimum elevation during the 20th century. Science 2008, 320, 1768–1771. [Google Scholar] [CrossRef]
- Rumpf, S.B.; Hülber, K.; Wessely, J.; Willner, W.; Moser, D.; Gattringer, A.; Klonner, G.; Zimmermann, N.E.; Dullinger, S. Extinction debts and colonization credits of non-forest plants in the European Alps. Nat. Commun. 2019, 10, 4293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, L.; Liu, Y.; Zhang, M.; An, C. Response of altitudinal vegetation belts of the Tianshan Mountains in northwestern China to climate change during 1989–2015. Sci. Rep. 2021, 11, 4870. [Google Scholar] [CrossRef] [PubMed]
- Zschg, A.P.; Frehner, M.; Gubelmann, P.; Augustin, S.; Brang, P.; Huber, B. Participatory modelling of upward shifts of altitudinal vegetation belts for assessing site type transformation in Swiss forests due to climate change. Appl. Veg. Sci. 2021, 24, e12621. [Google Scholar] [CrossRef]
- Blasi, C. La Vegetazione d’Italia, Carta delle Serie di Vegetazione, Scala 1:500 000; Palombi & Partner s.r.l.: Rome, Italy, 2010. [Google Scholar]
- Peterson, C.J. Catastrophic wind damage to North American forests and the potential impact of climate change. Sci. Total Environ. 2000, 262, 287–311. [Google Scholar] [CrossRef]
- Uriarte, M.; Canham, C.D.; Thompson, J.; Zimmerman, J.K.; Brokaw, N. Seedling recruitment in a hurricane-driven tropical forest: Light limitation, density-dependence and the spatial distribution of parent trees. J. Ecol. 2005, 93, 291–304. [Google Scholar] [CrossRef]
- Shiels, A.B.; Zimmerman, J.K.; Garcıa-Montiel, D.C.; Jonckheere, I.; Holm, J.; Horton, D.; Brokaw, N. Plant responses to simulated hurricane impacts in a subtropical wet forest, Puerto Rico. J. Ecol. 2010, 98, 659–673. [Google Scholar] [CrossRef]
- Canadell, J.; Jackson, R.B.; Ehleringer, J.R.; Mooney, H.A.; Sala, O.E.; Schulze, E.D. Maximum rooting depth of vegetation types at the global scale. Oecologia 1996, 108, 583–595. [Google Scholar] [CrossRef]
- Kutschera, L.; Lichtenegger, E. Wurzelatlas Mitteleuropäischer Waldbäume und Sträucher; Leopold Stocker Verlag: Graz, Austria; Stuttgart, Germany, 2002. [Google Scholar]
- Touliatos, P.; Roth, E. Hurricanes and trees: Ten lessons from Camille. J. For. 1971, 69, 285–289. [Google Scholar]
- Foster, D.R. Species and stand response to catastrophic wind in central New England, USA. J. Ecol. 1988, 76, 135–151. [Google Scholar] [CrossRef]
- Eriksson, M.; Pouttu, A.; Roininen, H. The influence of windthrow area and timber characteristics on colonization of wind-felled spruces by Ips typographus (L.). For. Ecol. Manag. 2005, 216, 105–116. [Google Scholar] [CrossRef]
- Schroeder, L.M.; Lindelöw, Å. Attacks on living spruce trees by the bark beetle Ips typographus (Col. Scolytidae) following a storm-felling: A comparison between stands with and without removal of wind-felled trees. Agric. For. Entomol. 2002, 4, 47–56. [Google Scholar] [CrossRef]
- Lindelöw, Å.; Risberg, B.; Sjödin, K. Attraction during flight of scolytids and other bark-and wood-dwelling beetles to volatiles from fresh and stored spruce wood. Can. J. For. Res. 1992, 22, 224–228. [Google Scholar] [CrossRef]
- Austarå, Ø.; Bakke, A.; Midtgaard, F. Response in Ips typographus to logging waste odours and synthetic pheromones. J. Appl. Entomol. 1986, 101, 194–198. [Google Scholar] [CrossRef]
- Hedgren, P.O.; Schroeder, L.M.; Weslien, J. Tree killing by Ips typographus (Coleoptera: Scolytidae) at stand edges with and without colonized felled spruce trees. Agric. For. Entomol. 2003, 5, 67–74. [Google Scholar] [CrossRef]
- Mulock, P.; Christiansen, E. The threshold of successful attack by Ips typographus on Picea abies: A field experiment. For. Ecol. Manag. 1986, 14, 125–132. [Google Scholar] [CrossRef]
- Byers, J.A.; Zhang, Q.H.; Schlyter, F.; Birgersson, G. Volatiles from nonhost birch trees inhibit pheromone response in spruce bark beetles. Sci. Nat. 1998, 85, 557–561. [Google Scholar] [CrossRef]
- Nieto, A.; Alexander, K.N.A. European Red List of Saproxylic Beetles; Publications Office of the European Union: Luxembourg, 2010. [Google Scholar]
- Leoni, V.; Giupponi, L.; Pavlovic, R.; Gianoncelli, C.; Cecati, F.; Ranzato, E.; Martinotti, S.; Pedrali, D.; Giorgi, A.; Panseri, S. Multidisciplinary analysis of Italian Alpine wildflower honey reveals criticalities, diversity and value. Sci. Rep. 2021, 11, 19316. [Google Scholar] [CrossRef] [PubMed]
- Steinert, M.; Eldegard, K.; Sydenham, M.A.; Moe, S.R. Bumble bee communities in power-line clearings: Effects of experimental management practices. Insect Conserv. Divers. 2021, 14, 377–392. [Google Scholar] [CrossRef]
- Svensson, B.; Lagerlöf, J.; Svensson, B.G. Habitat preferences of nest-seeking bumble bees (Hymenoptera: Apidae) in an agricultural landscape. Agric. Ecosyst. Environ. 2000, 77, 247–255. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Area | Cluster | Species | Phytosociological Class | Φ | p | |
|---|---|---|---|---|---|---|
| A | A1 | Picea abies | PIC | 0.89 | 0.01 | ** |
| Larix decidua | PIC | 0.74 | 0.01 | ** | ||
| Vaccinium myrtillus | PIC, LOI, QUE, NAR | 0.70 | 0.01 | ** | ||
| Arctostaphylos uva-ursi | PIC, LOI, | 0.69 | 0.01 | ** | ||
| Oxalis acetosella | PIC, FAG | 0.65 | 0.01 | ** | ||
| Viola biflora | VIR, MUL, MON, ASP | 0.55 | 0.03 | * | ||
| Rhododendron ferrugineum | LOI | 0.54 | 0.02 | * | ||
| Imperatoria ostruthium | MUL | 0.47 | 0.05 | * | ||
| A2 | Rubus idaeus | ROB | 0.66 | 0.01 | ** | |
| Luzula sylvatica | PIC, FAG, QUE, | 0.56 | 0.05 | * | ||
| A3 | Salix caprea | ROB | 0.61 | 0.01 | ** | |
| Lonicera alpigena | FAG | 0.53 | 0.04 | * | ||
| Petasites albus | MUL, EPI | 0.45 | 0.04 | * | ||
| B | B1 | Picea abies | PIC | 0.98 | 0.01 | ** |
| Luzula nivea | FAG | 0.72 | 0.03 | * | ||
| Veronica officinalis | QUE, NAR | 0.68 | 0.02 | * | ||
| B2 | Corylus avellana | RHA | 0.79 | 0.01 | ** | |
| B3 | Brachypodium rupestre | ERI, FES | 0.92 | 0.01 | ** | |
| Lotus corniculatus | MOL | 0.90 | 0.01 | ** | ||
| Achillea millefolium | MOL, ART | 0.83 | 0.01 | ** | ||
| Acer pseudoplatanus | FAG | 0.78 | 0.01 | ** | ||
| Astragalus glycyphyllos | GER | 0.69 | 0.01 | ** | ||
| Festuca valesiaca | FES | 0.63 | 0.05 | * | ||
| Fraxinus excelsior | FAG | 0.58 | 0.04 | * | ||
| B4 | Rubus idaeus | ROB | 0.93 | 0.01 | ** |
| Study Area | Vegetation Type | Richness | HS | T [1-5] | K [1-5] | L [1-5] | F [1-5] | R [1-5] | N [1-5] | H [1-5] | D [1-5] |
|---|---|---|---|---|---|---|---|---|---|---|---|
| A | A1 | 25.14 ± 6.67 | 1.90 ± 0.09 | 2.50 ± 0.13 | 3.07 ± 0.17 | 2.14 ± 0.25 | 3.11 ± 0.07 | 1.20 ± 0.22 | 2.60 ± 0.18 | 4.54 ± 0.29 | 2.21 ± 0.31 |
| A2 | 23.30 ± 7.93 | 1.72 ± 0.79 | 2.85 ± 0.12 | 3.01 ± 0.12 | 2.68 ± 0.27 | 3.05 ± 0.11 | 2.73 ± 0.23 | 3.32 ± 0.40 | 3.65 ± 0.50 | 2.46 ± 0.38 | |
| A3 | 19.25 ± 0.96 | 1.81 ± 0.61 | 2.74 ± 0.22 | 2.96 ± 0.03 | 2.46 ± 0.14 | 3.27 ± 0.19 | 2.75 ± 0.29 | 3.02 ± 0.39 | 3.86 ± 0.68 | 1.41 ± 0.22 | |
| B | B1 | 15.00 ± 3.83 | 0.90 ± 0.28 | 2.52 ± 0.06 | 3.14 ± 0.08 | 1.62 ± 0.30 | 2.97 ± 0.03 | 2.41 ± 0.16 | 2.83 ± 0.08 | 4.54 ± 0.25 | 2.98 ± 0.02 |
| B2 | 26.67 ± 6.71 | 1.54 ± 0.46 | 3.10 ± 0.05 | 3.02 ± 0.04 | 3.09 ± 0.09 | 2.94 ± 0.05 | 3.03 ± 0.02 | 3.09 ± 0.17 | 3.04 ± 0.08 | 2.79 ± 0.10 | |
| B3 | 26.00 ± 2.64 | 1.56 ± 0.25 | 3.27 ± 0.17 | 3.10 ± 0.07 | 3.66 ± 0.14 | 2.66 ± 0.11 | 3.54 ± 0.26 | 2.95 ± 0.06 | 3.02 ± 0.02 | 1.78 ± 0.34 | |
| B4 | 19.17 ± 5.56 | 1.17 ± 0.41 | 3.07 ± 0.09 | 3.02 ± 0.02 | 3.09 ± 0.18 | 2.97 ± 0.08 | 3.06 ± 0.16 | 3.65 ± 0.22 | 3.03 ± 0.02 | 2.72 ± 0.32 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giupponi, L.; Leoni, V.; Pedrali, D.; Giorgi, A. Restoration of Vegetation Greenness and Possible Changes in Mature Forest Communities in Two Forests Damaged by the Vaia Storm in Northern Italy. Plants 2023, 12, 1369. https://doi.org/10.3390/plants12061369
Giupponi L, Leoni V, Pedrali D, Giorgi A. Restoration of Vegetation Greenness and Possible Changes in Mature Forest Communities in Two Forests Damaged by the Vaia Storm in Northern Italy. Plants. 2023; 12(6):1369. https://doi.org/10.3390/plants12061369
Chicago/Turabian StyleGiupponi, Luca, Valeria Leoni, Davide Pedrali, and Annamaria Giorgi. 2023. "Restoration of Vegetation Greenness and Possible Changes in Mature Forest Communities in Two Forests Damaged by the Vaia Storm in Northern Italy" Plants 12, no. 6: 1369. https://doi.org/10.3390/plants12061369