
Niche Filling Dynamics of Ragweed (Ambrosia artemisiifolia L.) during Global Invasion

Abstract
:1. Introduction
2. Results
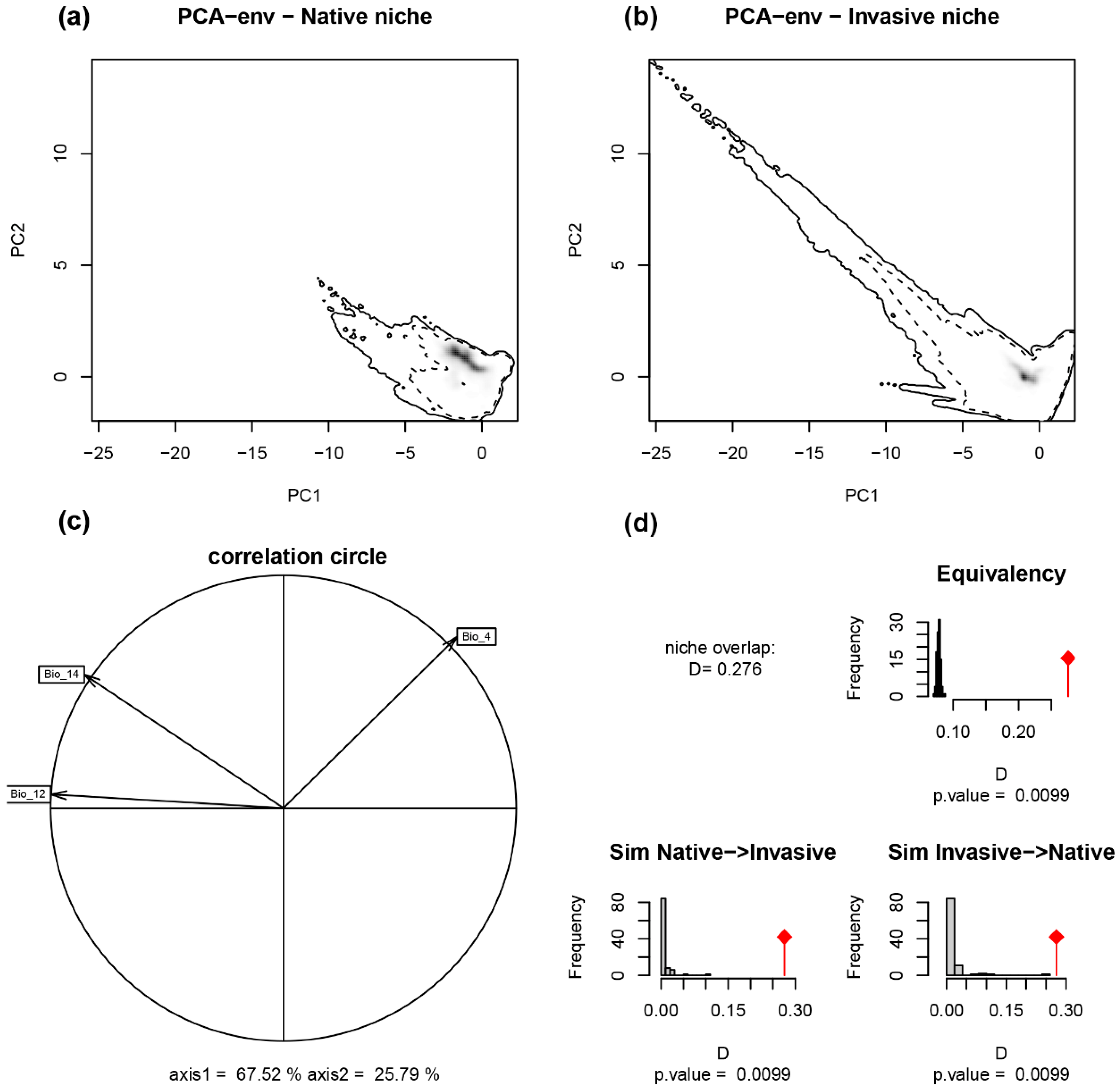
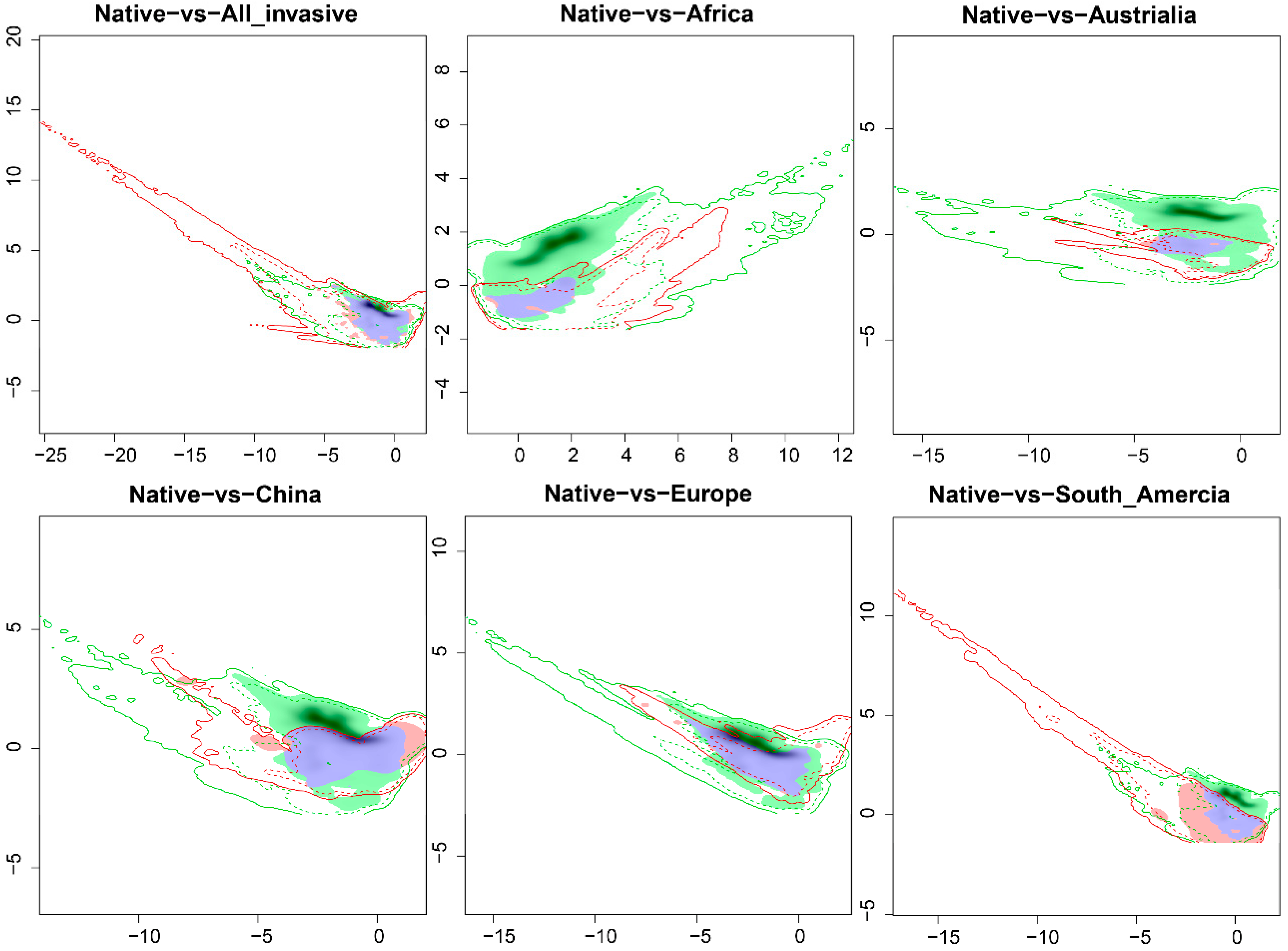
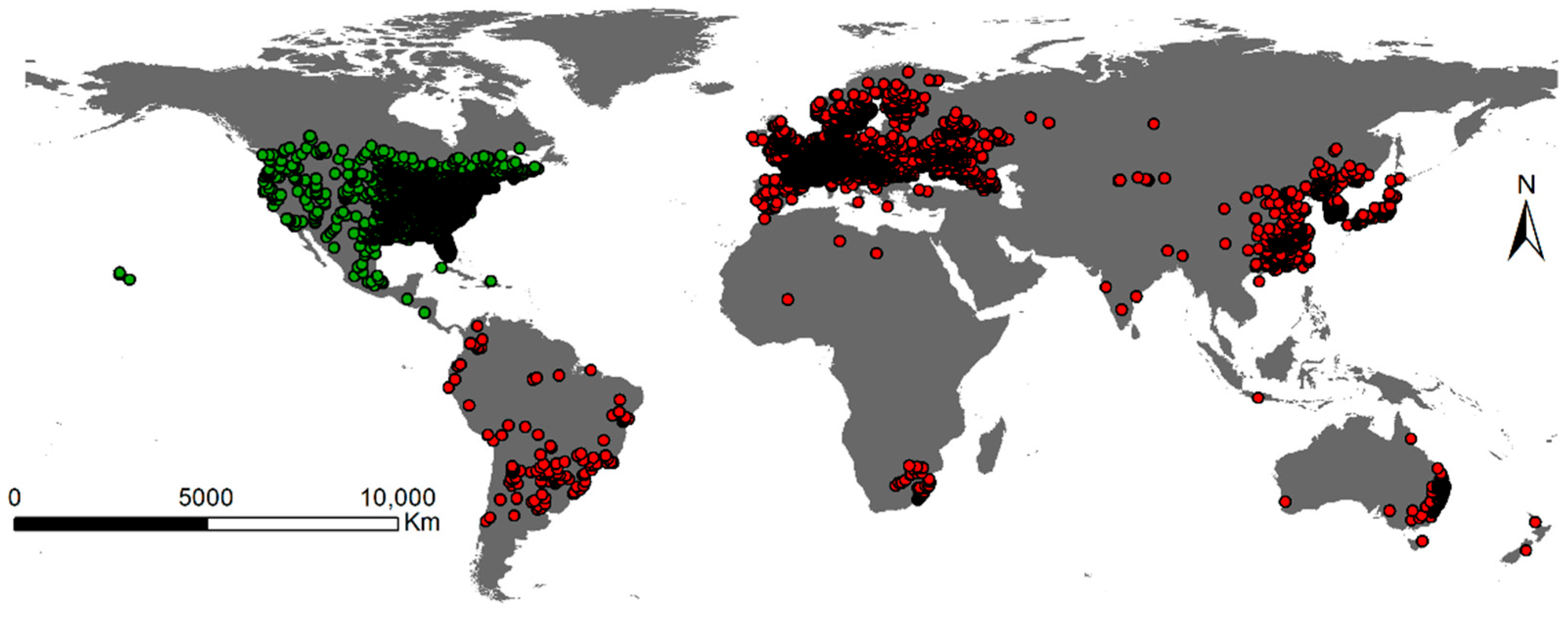
2.1. Niche Shift of A. artemisiifolia in Its Native and Invasive Range
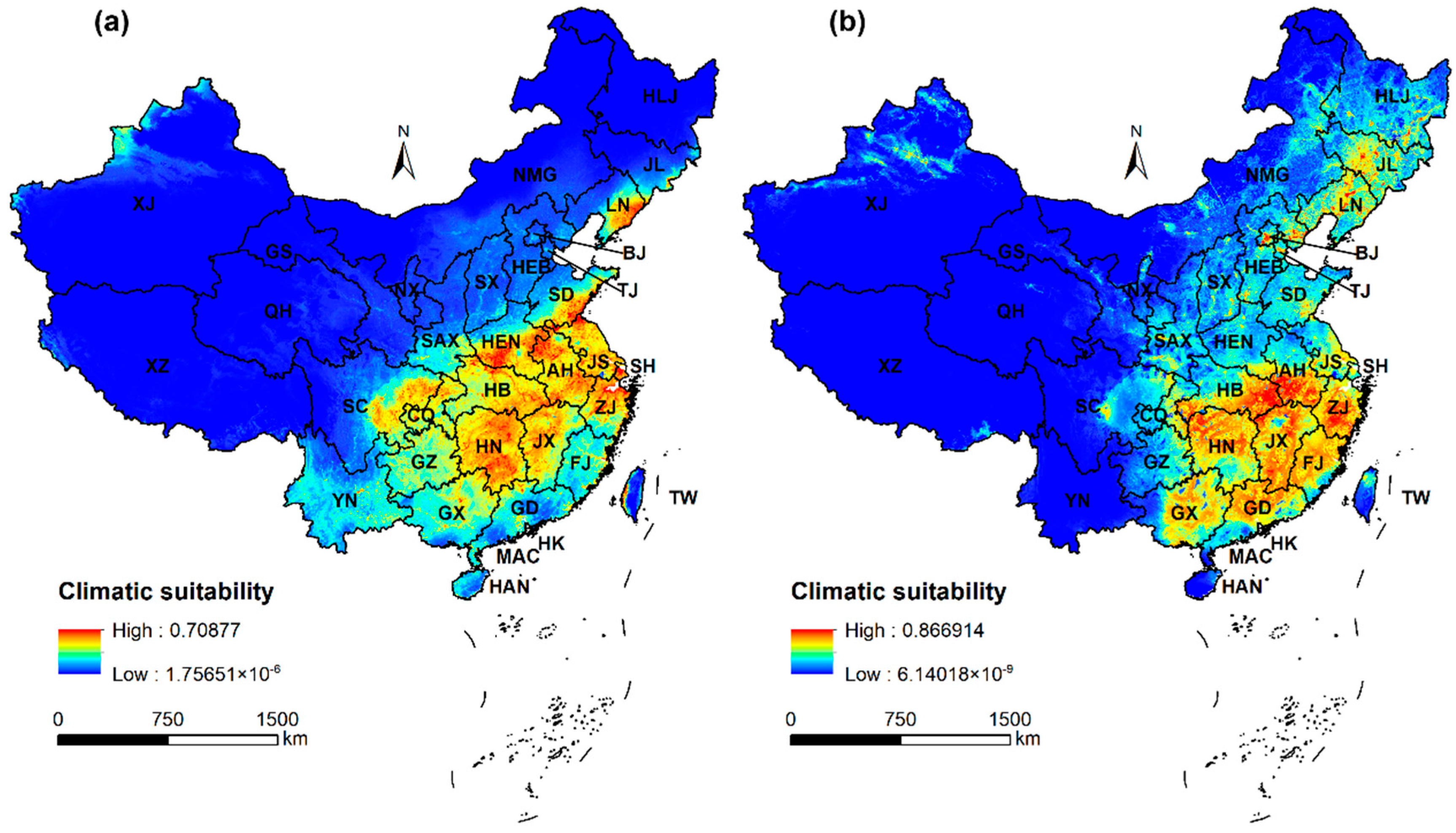
2.2. Predicted Potential Distribution of A. artemisiifolia in China
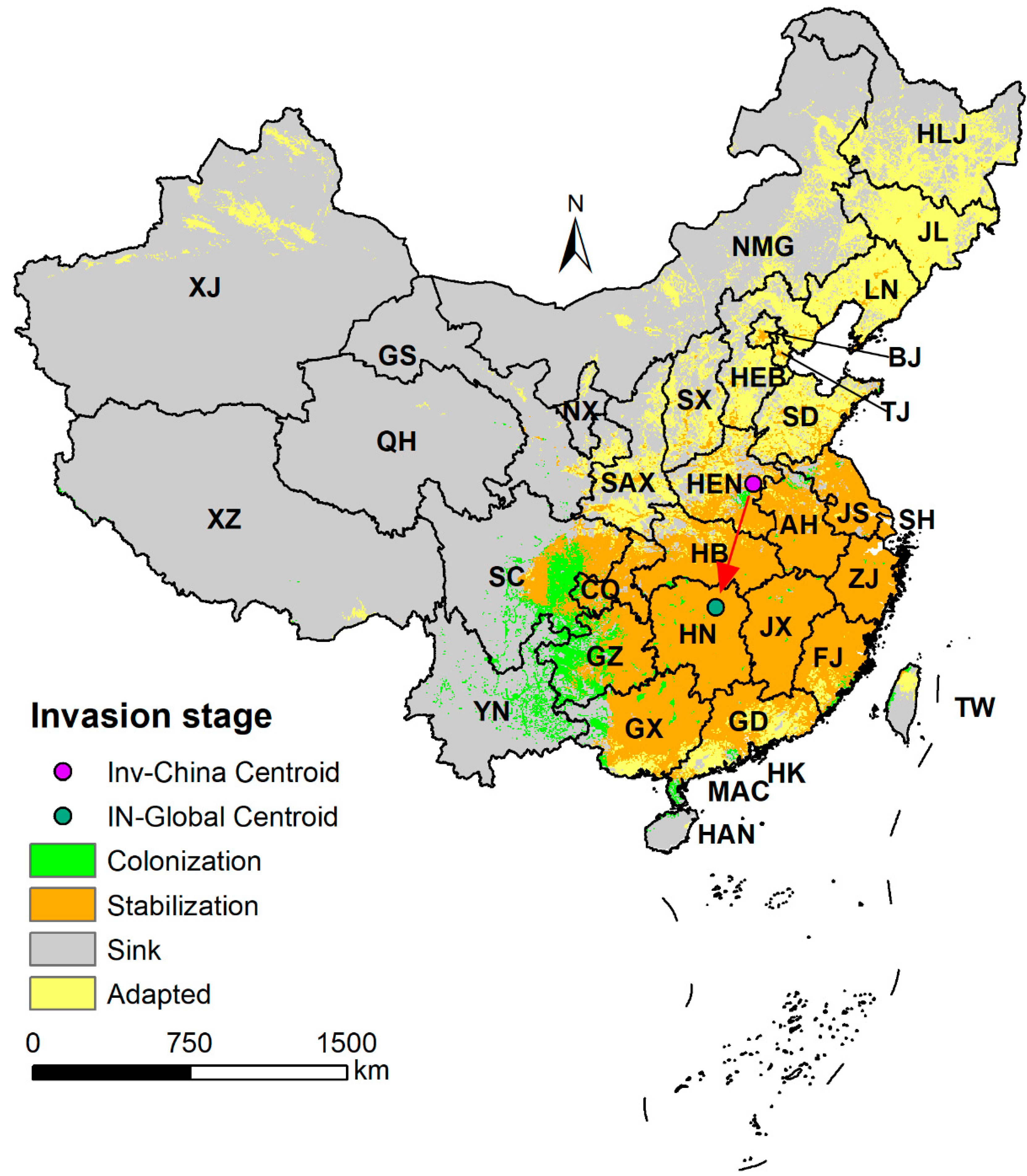
2.3. Invasion Stage Analysis
3. Discussion

4. Methods
4.1. Species Occurrence Data
4.2. Environmental Data
4.3. Ecological Niche Modeling
4.4. Climatic Niche Space Comparison: “COUE” Framework
4.5. Invasion Stage Analysis in China
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hall, C.M. Tourism and biological exchange and invasions: A missing dimension in sustainable tourism? Tour. Recreat. Res. 2015, 40, 81–94. [Google Scholar] [CrossRef]
- Rodríguez-Labajos, B.; Binimelis, R.; Monterroso, I. Multi-level driving forces of biological invasions. Ecol. Econ. 2009, 69, 63–75. [Google Scholar] [CrossRef]
- Meyerson, L.A.; Reaser, J.K. Biosecurity: Moving toward a comprehensive approach: A comprehensive approach to biosecurity is necessary to minimize the risk of harm caused by non-native organisms to agriculture, the economy, the environment, and human health. BioScience 2002, 52, 593–600. [Google Scholar] [CrossRef] [Green Version]
- Rockwell-Postel, M.; Laginhas, B.B.; Bradley, B.A. Supporting proactive management in the context of climate change: Prioritizing range-shifting invasive plants based on impact. Biol. Invasions 2020, 22, 2371–2383. [Google Scholar] [CrossRef]
- Xu, H.; Ding, H.; Li, M.; Qiang, S.; Guo, J.; Han, Z.; Huang, Z.; Sun, H.; He, S.; Wu, H. The distribution and economic losses of alien species invasion to China. Biol. Invasions 2006, 8, 1495–1500. [Google Scholar] [CrossRef]
- Bauer, J.T. Invasive species:“back-seat drivers” of ecosystem change? Biol. Invasions 2012, 14, 1295–1304. [Google Scholar] [CrossRef]
- Crowl, T.A.; Crist, T.O.; Parmenter, R.R.; Belovsky, G.; Lugo, A.E. The spread of invasive species and infectious disease as drivers of ecosystem change. Front. Ecol. Environ. 2008, 6, 238–246. [Google Scholar] [CrossRef]
- Doherty, T.S.; Glen, A.S.; Nimmo, D.G.; Ritchie, E.G.; Dickman, C.R. Invasive predators and global biodiversity loss. Proc. Natl. Acad. Sci. USA 2016, 113, 11261–11265. [Google Scholar] [CrossRef] [Green Version]
- Mainka, S.A.; Howard, G.W. Climate change and invasive species: Double jeopardy. Integr. Zool. 2010, 5, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.M.; Edwards, P.J. Limits to the niche and range margins of alien species. Oikos 2010, 119, 1377–1386. [Google Scholar] [CrossRef]
- Colautti, R.I.; Lau, J.A. Contemporary evolution during invasion: Evidence for differentiation, natural selection, and local adaptation. Mol. Ecol. 2015, 24, 1999–2017. [Google Scholar] [CrossRef] [PubMed]
- Shea, K.; Chesson, P. Community ecology theory as a framework for biological invasions. Trends Ecol. Evol. 2002, 17, 170–176. [Google Scholar] [CrossRef]
- González-Moreno, P.; Diez, J.M.; Richardson, D.M.; Vilà, M. Beyond climate: Disturbance niche shifts in invasive species. Glob. Ecol. Biogeogr. 2015, 24, 360–370. [Google Scholar] [CrossRef] [Green Version]
- Pili, A.N.; Tingley, R.; Sy, E.Y.; Diesmos, M.L.L.; Diesmos, A.C. Niche shifts and environmental non-equilibrium undermine the usefulness of ecological niche models for invasion risk assessments. Sci. Rep. 2020, 10, 7972. [Google Scholar] [CrossRef]
- Broennimann, O.; Treier, U.A.; Müller-Schärer, H.; Thuiller, W.; Peterson, A.; Guisan, A. Evidence of climatic niche shift during biological invasion. Ecol. Lett. 2007, 10, 701–709. [Google Scholar] [CrossRef] [Green Version]
- Peterson, A.T.; Vieglais, D.A. Predicting Species Invasions Using Ecological Niche Modeling: New Approaches from Bioinformatics Attack a Pressing Problem: A new approach to ecological niche modeling, based on new tools drawn from biodiversity informatics, is applied to the challenge of predicting potential species’ invasions. BioScience 2001, 51, 363–371. [Google Scholar] [CrossRef]
- Thuiller, W.; Richardson, D.M.; Pyšek, P.; Midgley, G.F.; Hughes, G.O.; Rouget, M. Niche-based modelling as a tool for predicting the risk of alien plant invasions at a global scale. Glob. Chang. Biol. 2005, 11, 2234–2250. [Google Scholar] [CrossRef]
- Oliveira, B.F.; Costa, G.C.; Fonseca, C.R. Niche dynamics of two cryptic Prosopis invading South American drylands. Biol. Invasions 2018, 20, 181–194. [Google Scholar] [CrossRef]
- Callen, S.T.; Miller, A.J. Signatures of niche conservatism and niche shift in the North American kudzu (Pueraria montana) invasion. Divers. Distrib. 2015, 21, 853–863. [Google Scholar] [CrossRef]
- Liu, C.; Wolter, C.; Xian, W.; Jeschke, J.M. Most invasive species largely conserve their climatic niche. Proc. Natl. Acad. Sci. USA 2020, 117, 23643–23651. [Google Scholar] [CrossRef]
- Petitpierre, B.; Kueffer, C.; Broennimann, O.; Randin, C.; Daehler, C.; Guisan, A. Climatic niche shifts are rare among terrestrial plant invaders. Science 2012, 335, 1344–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peterson, A.T. Ecological niche conservatism: A time-structured review of evidence. J. Biogeogr. 2011, 38, 817–827. [Google Scholar] [CrossRef]
- Williamson, M. Explaining and predicting the success of invading species at different stages of invasion. Biol. Invasions 2006, 8, 1561–1568. [Google Scholar] [CrossRef]
- Beaumont, L.J.; Gallagher, R.V.; Thuiller, W.; Downey, P.O.; Leishman, M.R.; Hughes, L. Different climatic envelopes among invasive populations may lead to underestimations of current and future biological invasions. Divers. Distrib. 2009, 15, 409–420. [Google Scholar] [CrossRef]
- Fitzpatrick, M.C.; Dunn, R.R.; Sanders, N.J. Data sets matter, but so do evolution and ecology. Glob. Ecol. Biogeogr. 2008, 17, 562–565. [Google Scholar] [CrossRef]
- Fitzpatrick, M.C.; Weltzin, J.F.; Sanders, N.J.; Dunn, R.R. The biogeography of prediction error: Why does the introduced range of the fire ant over-predict its native range? Glob. Ecol. Biogeogr. 2007, 16, 24–33. [Google Scholar] [CrossRef]
- Gallaher, T.; Merlin, M. Biology and impacts of Pacific island invasive species. 6. Prosopis pallida and Prosopis juliflora (Algarroba, Mesquite, Kiawe)(Fabaceae). Pac. Sci. 2010, 64, 489–526. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; LeBrun, E.G.; Stohlgren, T.J.; Stabach, J.A.; McDonald, D.L.; Oi, D.H.; LaPolla, J.S. Evidence of niche shift and global invasion potential of the Tawny Crazy ant, Nylanderia fulva. Ecol. Evol. 2015, 5, 4628–4641. [Google Scholar] [CrossRef]
- Medley, K.A. Niche shifts during the global invasion of the Asian tiger mosquito, Aedes albopictus Skuse (Culicidae), revealed by reciprocal distribution models. Glob. Ecol. Biogeogr. 2010, 19, 122–133. [Google Scholar] [CrossRef]
- Petersen, M.J. Evidence of a climatic niche shift following North American introductions of two crane flies (Diptera; genus Tipula). Biol. Invasions 2013, 15, 885–897. [Google Scholar] [CrossRef]
- Tingley, R.; Vallinoto, M.; Sequeira, F.; Kearney, M.R. Realized niche shift during a global biological invasion. Proc. Natl. Acad. Sci. USA 2014, 111, 10233–10238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Václavík, T.; Meentemeyer, R.K. Equilibrium or not? Modelling potential distribution of invasive species in different stages of invasion. Divers. Distrib. 2012, 18, 73–83. [Google Scholar] [CrossRef]
- Guisan, A.; Petitpierre, B.; Broennimann, O.; Daehler, C.; Kueffer, C. Unifying niche shift studies: Insights from biological invasions. Trends Ecol. Evol. 2014, 29, 260–269. [Google Scholar] [CrossRef] [Green Version]
- Chauvel, B.; Dessaint, F.; Cardinal-Legrand, C.; Bretagnolle, F. The historical spread of Ambrosia artemisiifolia L. in France from herbarium records. J. Biogeogr. 2006, 33, 665–673. [Google Scholar] [CrossRef]
- Bass, D.J.; Delpech, V.; Beard, J.; Bass, P.; Walls, R.S. Ragweed in Australia. Aerobiologia 2000, 16, 107–111. [Google Scholar] [CrossRef]
- Guo, J.-Y.; Zhou, Z.-S.; Zheng, X.-W.; Chen, H.-S.; Wan, F.-H.; Luo, Y.-H. Control efficiency of leaf beetle, Ophraella communa, on the invasive common ragweed, Ambrosia artemisiifolia, at different growing stages. Biocontrol Sci. Technol. 2011, 21, 1049–1063. [Google Scholar] [CrossRef]
- Darlington, H.T. Dr. WJ Beal’s seed-viability experiment. Am. J. Bot. 1922, 9, 266–269. [Google Scholar] [CrossRef]
- Cao, Z.; Wang, H.; Meng, L.; Li, B. Risk to nontarget plants from Ophraella communa (Coleoptera: Chrysomelidae), a potential biological control agent of alien invasive weed Ambrosia artemisiifolia (Asteraceae) in China. Appl. Entomol. Zool. 2011, 46, 375–381. [Google Scholar] [CrossRef]
- Liu, J.-X.; Wang, M.; Chen, B.-X.; Jin, P.; Li, J.-Y.; Zeng, K. Microsporogenesis, microgametogenesis, and pollen morphology of Ambrosia artemisiifolia L. in China. Plant Syst. Evol. 2012, 298, 43–50. [Google Scholar] [CrossRef]
- Sang, W.; Liu, X.; Axmacher, J.C. Germination and emergence of Ambrosia artemisiifolia L. under changing environmental conditions in China. Plant Species Biol. 2011, 26, 125–133. [Google Scholar] [CrossRef]
- Peterson, A.T. Predicting the geography of species’ invasions via ecological niche modeling. Q. Rev. Biol. 2003, 78, 419–433. [Google Scholar] [CrossRef] [Green Version]
- Chapman, D.S.; Scalone, R.; Štefanić, E.; Bullock, J.M. Mechanistic species distribution modeling reveals a niche shift during invasion. Ecology 2017, 98, 1671–1680. [Google Scholar] [CrossRef] [Green Version]
- Cunze, S.; Leiblein, M.C.; Tackenberg, O. Range expansion of Ambrosia artemisiifolia in Europe is promoted by climate change. Int. Sch. Res. Not. 2013, 2013, 610126. [Google Scholar] [CrossRef] [Green Version]
- Prentis, P.J.; Wilson, J.R.; Dormontt, E.E.; Richardson, D.M.; Lowe, A.J. Adaptive evolution in invasive species. Trends Plant Sci. 2008, 13, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.E. Evolutionary genetics of invasive species. Trends Ecol. Evol. 2002, 17, 386–391. [Google Scholar] [CrossRef]
- Atwater, D.Z.; Ervine, C.; Barney, J.N. Climatic niche shifts are common in introduced plants. Nat. Ecol. Evol. 2018, 2, 34–43. [Google Scholar] [CrossRef] [Green Version]
- Strubbe, D.; Beauchard, O.; Matthysen, E. Niche conservatism among non-native vertebrates in Europe and North America. Ecography 2015, 38, 321–329. [Google Scholar] [CrossRef]
- Strubbe, D.; Broennimann, O.; Chiron, F.; Matthysen, E. Niche conservatism in non-native birds in Europe: Niche unfilling rather than niche expansion. Glob. Ecol. Biogeogr. 2013, 22, 962–970. [Google Scholar] [CrossRef]
- Gerber, E.; Schaffner, U.; Gassmann, A.; Hinz, H.; Seier, M.; MÜLLER-SCHÄRER, H. Prospects for biological control of Ambrosia artemisiifolia in Europe: Learning from the past. Weed Res. 2011, 51, 559–573. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.-S.; Chen, H.-S.; Zheng, X.-W.; Guo, J.-Y.; Guo, W.; Li, M.; Luo, M.; Wan, F.-H. Control of the invasive weed Ambrosia artemisiifolia with Ophraella communa and Epiblema strenuana. Biocontrol Sci. Technol. 2014, 24, 950–964. [Google Scholar] [CrossRef]
- Zhou, Z.-S.; Guo, J.-Y.; Chen, H.-S.; Wan, F.-H. Effects of temperature on survival, development, longevity, and fecundity of Ophraella communa (Coleoptera: Chrysomelidae), a potential biological control agent against Ambrosia artemisiifolia (Asterales: Asteraceae). Environ. Entomol. 2010, 39, 1021–1027. [Google Scholar] [CrossRef] [Green Version]
- Brandes, D.; Nitzsche, J. Biology, introduction, dispersal, and distribution of common ragweed (Ambrosia artemisiifolia L.) with special regard to Germany. Nachr.-Dtsch. Pflanzenschutzd. Braunschw. 2006, 58, 286. [Google Scholar] [CrossRef]
- Gaudeul, M.; Giraud, T.; Kiss, L.; Shykoff, J.A. Nuclear and chloroplast microsatellites show multiple introductions in the worldwide invasion history of common ragweed, Ambrosia artemisiifolia. PLoS ONE 2011, 6, e17658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maron, J.L.; Vila, M. When do herbivores affect plant invasion? Evidence for the natural enemies and biotic resistance hypotheses. Oikos 2001, 95, 361–373. [Google Scholar] [CrossRef] [Green Version]
- Cartwright, R.; Templeton, G. Biological limitations of Protomyces gravidus as a mycoherbicide for giant ragweed, Ambrosia trifida. Plant Dis. 1988, 72, 580–582. [Google Scholar] [CrossRef]
- Hartmann, H.; Watson, A. Damage to common ragweed (Ambrosia artemisiifolia) caused by the white rust fungus (Albugo tragopogi). Weed Sci. 1980, 28, 632–635. [Google Scholar] [CrossRef]
- Iannella, M.; De Simone, W.; D’Alessandro, P.; Console, G.; Biondi, M. Investigating the current and future co-occurrence of Ambrosia artemisiifolia and Ophraella communa in Europe through ecological modelling and remote sensing data analysis. Int. J. Environ. Res. Public Health 2019, 16, 3416. [Google Scholar] [CrossRef] [Green Version]
- Leiblein-Wild, M.C.; Kaviani, R.; Tackenberg, O. Germination and seedling frost tolerance differ between the native and invasive range in common ragweed. Oecologia 2014, 174, 739–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallien, L.; Thuiller, W.; Fort, N.; Boleda, M.; Alberto, F.J.; Rioux, D.; Laine, J.; Lavergne, S. Is there any evidence for rapid, genetically-based, climatic niche expansion in the invasive common ragweed? PLoS ONE 2016, 11, e0152867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.-M.; She, D.-Y.; Zhang, D.-Y.; Liao, W.-J. Life history trait differentiation and local adaptation in invasive populations of Ambrosia artemisiifolia in China. Oecologia 2015, 177, 669–677. [Google Scholar] [CrossRef]
- Li, F.; van Kleunen, M.; Li, J.; Liu, X.; Gao, K.; Zhu, J.; Zhao, X.; Zhao, C.; Li, J. Patterns of genetic variation reflect multiple introductions and pre-admixture sources of common ragweed (Ambrosia artemisiifolia) in China. Biol. Invasions 2019, 21, 2191–2209. [Google Scholar] [CrossRef]
- Li, X.-M.; Liao, W.-J.; Wolfe, L.M.; Zhang, D.-Y. No evolutionary shift in the mating system of North American Ambrosia artemisiifolia (Asteraceae) following its introduction to China. PLoS ONE 2012, 7, e31935. [Google Scholar] [CrossRef]
- Sun, Y.; Roderick, G.K. Rapid evolution of invasive traits facilitates the invasion of common ragweed, Ambrosia artemisiifolia. J. Ecol. 2019, 107, 2673–2687. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.-L.; Li, H.-Q.; Wang, J.-H.; Sun, X.-P.; Fu, Y.-Y.; Xing, L.-G. The current and future potential geographical distribution of common ragweed, Ambrosia artemisiifolia in China. Pak. J. Bot 2021, 53, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, J.; Zhao, C.; Quan, Z.; Zhao, X.; Gong, L. Prediction of potential suitable area of Ambrosia artemisiifolia L. in China based on MAXENT and ArcGIS. Acta Phytophyl. Sin. 2016, 43, 1041–1048. [Google Scholar] [CrossRef]
- Chen, H.; Chen, L.; Albright, T.P. Predicting the potential distribution of invasive exotic species using GIS and information-theoretic approaches: A case of ragweed (Ambrosia artemisiifolia L.) distribution in China. Chin. Sci. Bull. 2007, 52, 1223–1230. [Google Scholar] [CrossRef]
- Chapple, D.G.; Whitaker, A.H.; Chapple, S.N.; Miller, K.A.; Thompson, M.B. Biosecurity interceptions of an invasive lizard: Origin of stowaways and human-assisted spread within New Zealand. Evol. Appl. 2013, 6, 324–339. [Google Scholar] [CrossRef] [PubMed]
- Tingley, R.; Thompson, M.B.; Hartley, S.; Chapple, D.G. Patterns of niche filling and expansion across the invaded ranges of an Australian lizard. Ecography 2016, 39, 270–280. [Google Scholar] [CrossRef]
- Chen, J.; Ma, F.; Zhang, Y.; Wang, C.; Xu, H. Spatial distribution patterns of invasive alien species in China. Glob. Ecol. Conserv. 2021, 26, e01432. [Google Scholar] [CrossRef]
- Fonseca, É.; Both, C.; Cechin, S.Z. Introduction pathways and socio-economic variables drive the distribution of alien amphibians and reptiles in a megadiverse country. Divers. Distrib. 2019, 25, 1130–1141. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Bian, Z.; Ren, W.; Wu, J.; Liu, J.; Shrestha, N. Spatial patterns and hotspots of plant invasion in China. Glob. Ecol. Conserv. 2023, 43, e02424. [Google Scholar] [CrossRef]
- Bertelsmeier, C.; Ollier, S.; Liebhold, A.; Keller, L. Recent human history governs global ant invasion dynamics. Nat. Ecol. Evol. 2017, 1, 0184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dyer, E.E.; Cassey, P.; Redding, D.W.; Collen, B.; Franks, V.; Gaston, K.J.; Jones, K.E.; Kark, S.; Orme, C.D.L.; Blackburn, T.M. The global distribution and drivers of alien bird species richness. PLoS Biol. 2017, 15, e2000942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blackburn, T.M.; Delean, S.; Pyšek, P.; Cassey, P. On the island biogeography of aliens: A global analysis of the richness of plant and bird species on oceanic islands. Glob. Ecol. Biogeogr. 2016, 25, 859–868. [Google Scholar] [CrossRef] [Green Version]
- Simberloff, D. The role of propagule pressure in biological invasions. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 81–102. [Google Scholar] [CrossRef]
- Beans, C.M.; Kilkenny, F.F.; Galloway, L.F. Climate suitability and human influences combined explain the range expansion of an invasive horticultural plant. Biol. Invasions 2012, 14, 2067–2078. [Google Scholar] [CrossRef] [Green Version]
- Bucklin, D.N.; Basille, M.; Benscoter, A.M.; Brandt, L.A.; Mazzotti, F.J.; Romanach, S.S.; Speroterra, C.; Watling, J.I. Comparing species distribution models constructed with different subsets of environmental predictors. Divers. Distrib. 2015, 21, 23–35. [Google Scholar] [CrossRef]
- Zhu, G.; Li, H.; Zhao, L. Incorporating anthropogenic variables into ecological niche modeling to predict areas of invasion of Popillia japonica. J. Pest Sci. 2017, 90, 151–160. [Google Scholar] [CrossRef]
- Xian, X.; Zhao, H.; Wang, R.; Huang, H.; Chen, B.; Zhang, G.; Liu, W.; Wan, F. Climate change has increased the global threats posed by three ragweeds (Ambrosia L.) in the Anthropocene. Sci. Total Environ. 2023, 859, 160252. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef] [Green Version]
- Chapman, D.S.; Makra, L.; Albertini, R.; Bonini, M.; Páldy, A.; Rodinkova, V.; Šikoparija, B.; Weryszko-Chmielewska, E.; Bullock, J.M. Modelling the introduction and spread of non-native species: International trade and climate change drive ragweed invasion. Glob. Chang. Biol. 2016, 22, 3067–3079. [Google Scholar] [CrossRef] [Green Version]
- Catford, J.A.; Daehler, C.C.; Murphy, H.T.; Sheppard, A.W.; Hardesty, B.D.; Westcott, D.A.; Rejmánek, M.; Bellingham, P.J.; Pergl, J.; Horvitz, C.C. The intermediate disturbance hypothesis and plant invasions: Implications for species richness and management. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 231–241. [Google Scholar] [CrossRef]
- With, K.A. The landscape ecology of invasive spread. Conserv. Biol. 2002, 16, 1192–1203. [Google Scholar] [CrossRef] [Green Version]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Graham, C.H.; Master, L.L.; Albert, D.L. The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 2006, 29, 773–785. [Google Scholar] [CrossRef]
- Liu, X.; Liu, H.; Gong, H.; Lin, Z.; Lv, S. Appling the one-class classification method of maxent to detect an invasive plant Spartina alterniflora with time-series analysis. Remote Sens. 2017, 9, 1120. [Google Scholar] [CrossRef] [Green Version]
- Guisan, A.; Zimmermann, N.E.; Elith, J.; Graham, C.H.; Phillips, S.; Peterson, A.T. What matters for predicting the occurrences of trees: Techniques, data, or species’ characteristics? Ecol. Monogr. 2007, 77, 615–630. [Google Scholar] [CrossRef] [Green Version]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Syfert, M.M.; Smith, M.J.; Coomes, D.A. The effects of sampling bias and model complexity on the predictive performance of MaxEnt species distribution models. PLoS ONE 2013, 8, e55158. [Google Scholar] [CrossRef]
- GBIF. Global Biodiversity Information Facility (GBIF) (23 August 2021) GBIF Occurrence Download. 2021. Available online: https://www.gbif.org/ (accessed on 23 August 2021).
- Early, R.; Bradley, B.A.; Dukes, J.S.; Lawler, J.J.; Olden, J.D.; Blumenthal, D.M.; Gonzalez, P.; Grosholz, E.D.; Ibañez, I.; Miller, L.P. Global threats from invasive alien species in the twenty-first century and national response capacities. Nat. Commun. 2016, 7, 12485. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Wildlife Conservation Society—WCS, Center for International Earth Science Information Network—CIESIN—Columbia University. Last of the Wild Project, Version 2, 2005 (LWP-2): Global Human Footprint Dataset (Geographic). 2005. Available online: https://doi.org/10.7927/H4BP00QC (accessed on 27 July 2021).
- Warren, D.L.; Glor, R.E.; Turelli, M. ENMTools: A toolbox for comparative studies of environmental niche models. Ecography 2010, 33, 607–611. [Google Scholar] [CrossRef]
- Tang, X.; Yuan, Y.; Liu, X.; Zhang, J. Potential range expansion and niche shift of the invasive Hyphantria cunea between native and invasive countries. Ecol. Entomol. 2021, 46, 910–925. [Google Scholar] [CrossRef]
- Yan, H.; Feng, L.; Zhao, Y.; Feng, L.; Wu, D.; Zhu, C. Prediction of the spatial distribution of Alternanthera philoxeroides in China based on ArcGIS and MaxEnt. Glob. Ecol. Conserv. 2020, 21, e00856. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Modell. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Cobos, M.E.; Peterson, A.T.; Barve, N.; Osorio-Olvera, L. kuenm: An R package for detailed development of ecological niche models using Maxent. PeerJ 2019, 7, e6281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broennimann, O.; Fitzpatrick, M.C.; Pearman, P.B.; Petitpierre, B.; Pellissier, L.; Yoccoz, N.G.; Thuiller, W.; Fortin, M.J.; Randin, C.; Zimmermann, N.E. Measuring ecological niche overlap from occurrence and spatial environmental data. Glob. Ecol. Biogeogr. 2012, 21, 481–497. [Google Scholar] [CrossRef] [Green Version]
- Schoener, T.W. Nonsynchronous spatial overlap of lizards in patchy habitats. Ecology 1970, 51, 408–418. [Google Scholar] [CrossRef] [Green Version]
- Warren, D.L.; Glor, R.E.; Turelli, M. Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evol. Int. J. Org. Evol. 2008, 62, 2868–2883. [Google Scholar] [CrossRef]
- Di Cola, V.; Broennimann, O.; Petitpierre, B.; Breiner, F.T.; d’Amen, M.; Randin, C.; Engler, R.; Pottier, J.; Pio, D.; Dubuis, A. ecospat: An R package to support spatial analyses and modeling of species niches and distributions. Ecography 2017, 40, 774–787. [Google Scholar] [CrossRef]
- Gallien, L.; Douzet, R.; Pratte, S.; Zimmermann, N.E.; Thuiller, W. Invasive species distribution models–how violating the equilibrium assumption can create new insights. Glob. Ecol. Biogeogr. 2012, 21, 1126–1136. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Invasive Region | Niche Overlap | Niche Equivalency | Niche Similarity | Unfilling | Stability | Expansion | |
|---|---|---|---|---|---|---|---|
| D | p-Value | Native > Invasive p-Value | Invasive > Native p-Value | ||||
| All Invasive | 0.276 | 0.0099 | 0.0099 | 0.0099 | 0.276 | 0.992 | 0.008 |
| Africa | 0.045 | 0.0003 | 0.0396 | 0.0792 | 0.979 | 0.947 | 0.053 |
| Australia | 0.210 | 0.3267 | 0.0099 | 0.0099 | 0.964 | 0.975 | 0.025 |
| China | 0.122 | 0.0099 | 0.0198 | 0.0099 | 0.663 | 0.952 | 0.048 |
| Europe | 0.226 | 0.0099 | 0.0099 | 0.0099 | 0.559 | 0.999 | 0.001 |
| South America | 0.068 | 0.0099 | 0.0396 | 0.0495 | 0.846 | 0.593 | 0.407 |
| Environmental Variable | Native | Inv-China | IN-Global | |||
|---|---|---|---|---|---|---|
| Percent Contribution | Permutation Importance | Percent Contribution | Permutation Importance | Percent Contribution | Permutation Importance | |
| Human influence index (HII) | 35.8 | 37.8 | 43.1 | 32.9 | 40.3 | 46.1 |
| Temperature seasonality (BIO_4, standard deviation × 100) | 14.7 | 35.7 | 23.9 | 39.1 | 38.7 | 34.6 |
| Annual precipitation (BIO_12, mm) | 10.2 | 17.1 | 21.8 | 20 | 4.1 | 8.9 |
| Precipitation of driest month (BIO_14, mm) | 39.2 | 9.3 | 11.2 | 8 | 16.8 | 10.3 |
| Variables | Description | Units |
|---|---|---|
| Bio_1 | Annual mean temperature | °C |
| Bio_2 | Mean diurnal range | °C |
| Bio_3 | Isothermality | / |
| Bio_4 | Temperature seasonality | / |
| Bio_5 | Max temperature of warmest month | °C |
| Bio_6 | Min temperature of coldest month | °C |
| Bio_7 | Temperature annual range | °C |
| Bio_8 | Mean temperature of wettest quarter | °C |
| Bio_9 | Mean temperature of driest quarter | °C |
| Bio_10 | Mean temperature of warmest quarter | °C |
| Bio_11 | Mean temperature of coldest quarter | °C |
| Bio_12 | Annual precipitation | mm |
| Bio_13 | Precipitation of wettest month | mm |
| Bio_14 | Precipitation of driest month | mm |
| Bio_15 | Precipitation seasonality | / |
| Bio_16 | Precipitation of wettest quarter | mm |
| Bio_17 | Precipitation of driest quarter | mm |
| Bio_18 | Precipitation of warmest quarter | mm |
| Bio_19 | Precipitation of coldest quarter | mm |
| Alt | Elevation | m |
| HII | Human Influence Index | / |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, X.-J.; Liu, G.; Qian, Z.-Q.; Zhu, Z.-H. Niche Filling Dynamics of Ragweed (Ambrosia artemisiifolia L.) during Global Invasion. Plants 2023, 12, 1313. https://doi.org/10.3390/plants12061313
Song X-J, Liu G, Qian Z-Q, Zhu Z-H. Niche Filling Dynamics of Ragweed (Ambrosia artemisiifolia L.) during Global Invasion. Plants. 2023; 12(6):1313. https://doi.org/10.3390/plants12061313
Chicago/Turabian StyleSong, Xing-Jiang, Gang Liu, Zeng-Qiang Qian, and Zhi-Hong Zhu. 2023. "Niche Filling Dynamics of Ragweed (Ambrosia artemisiifolia L.) during Global Invasion" Plants 12, no. 6: 1313. https://doi.org/10.3390/plants12061313