
Increased Leaf Bacterial Network Complexity along the Native Plant Diversity Gradient Facilitates Plant Invasion?
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
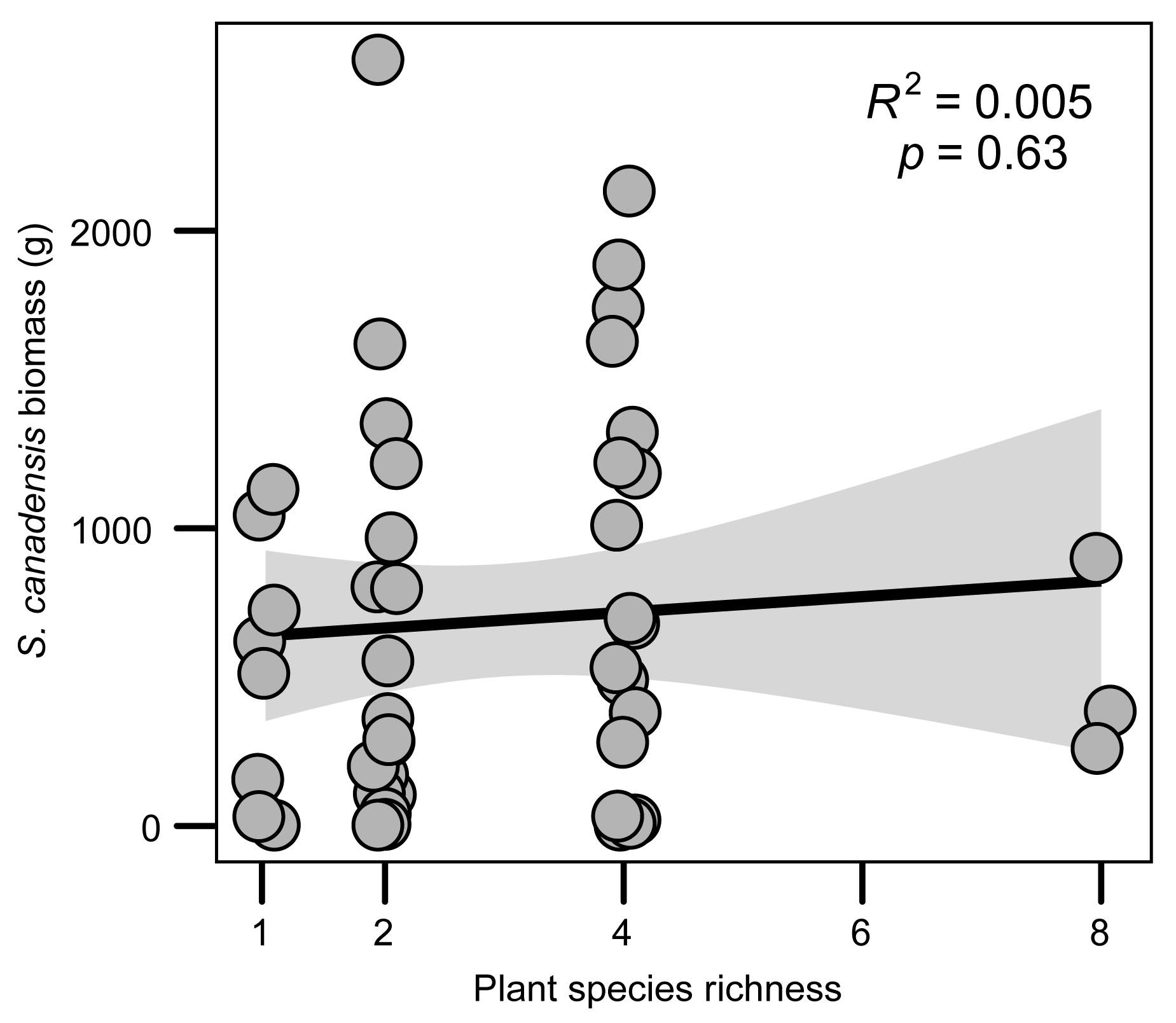
2.1. Plant Richness and Invader Biomass
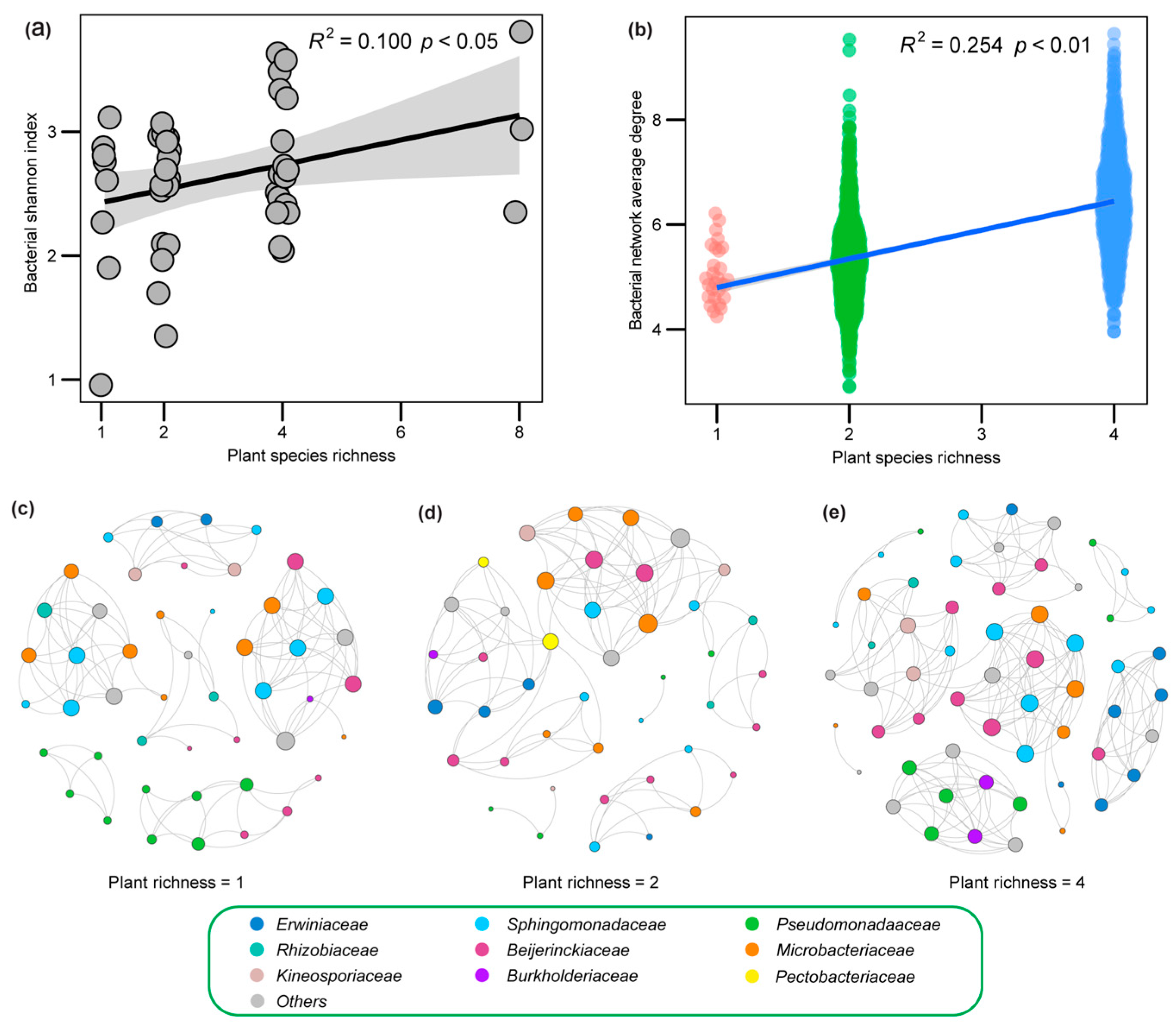
2.2. Plant Richness and Leaf Bacteria Community
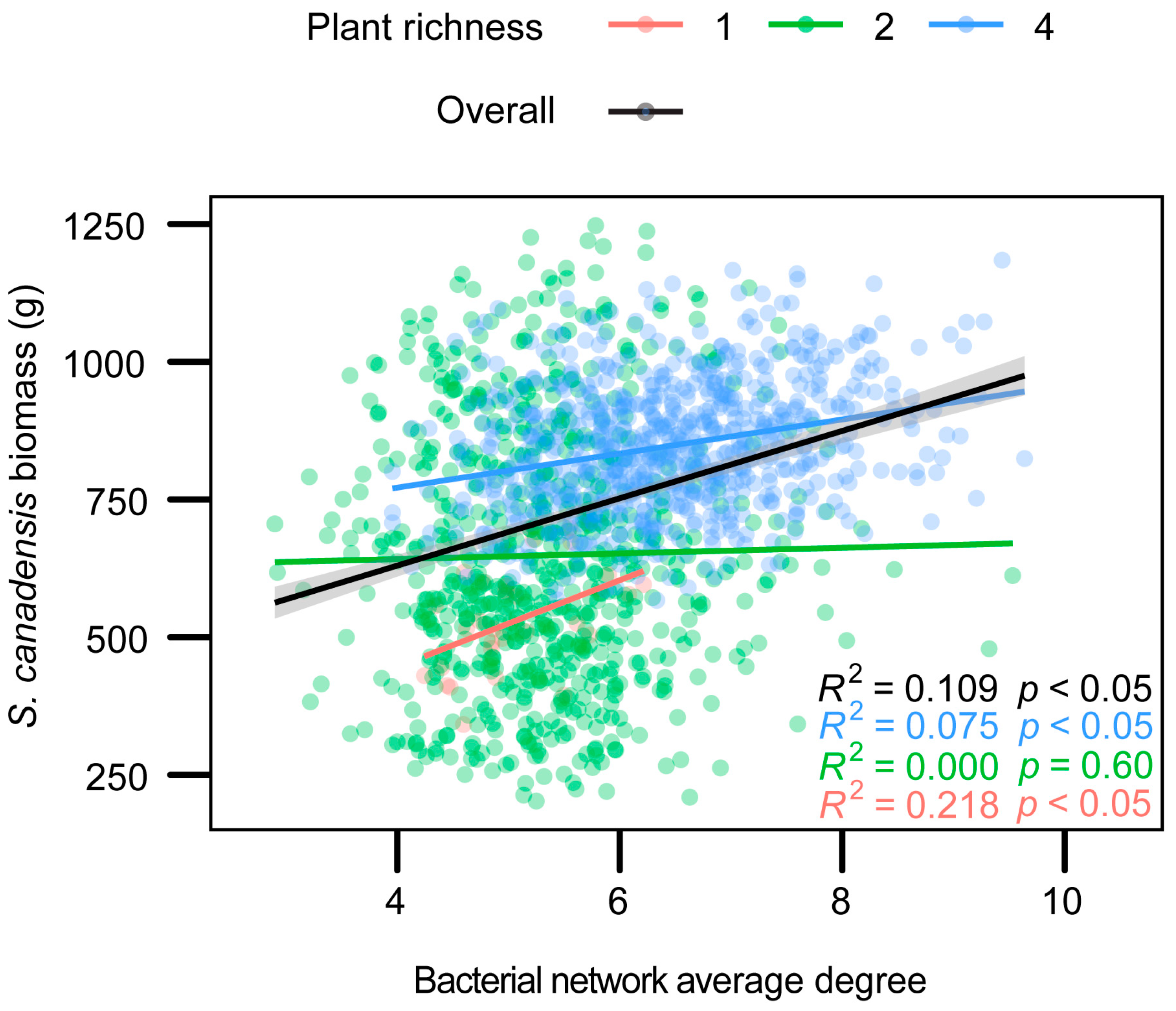
2.3. Leaf Bacteria Community and Invader Biomass
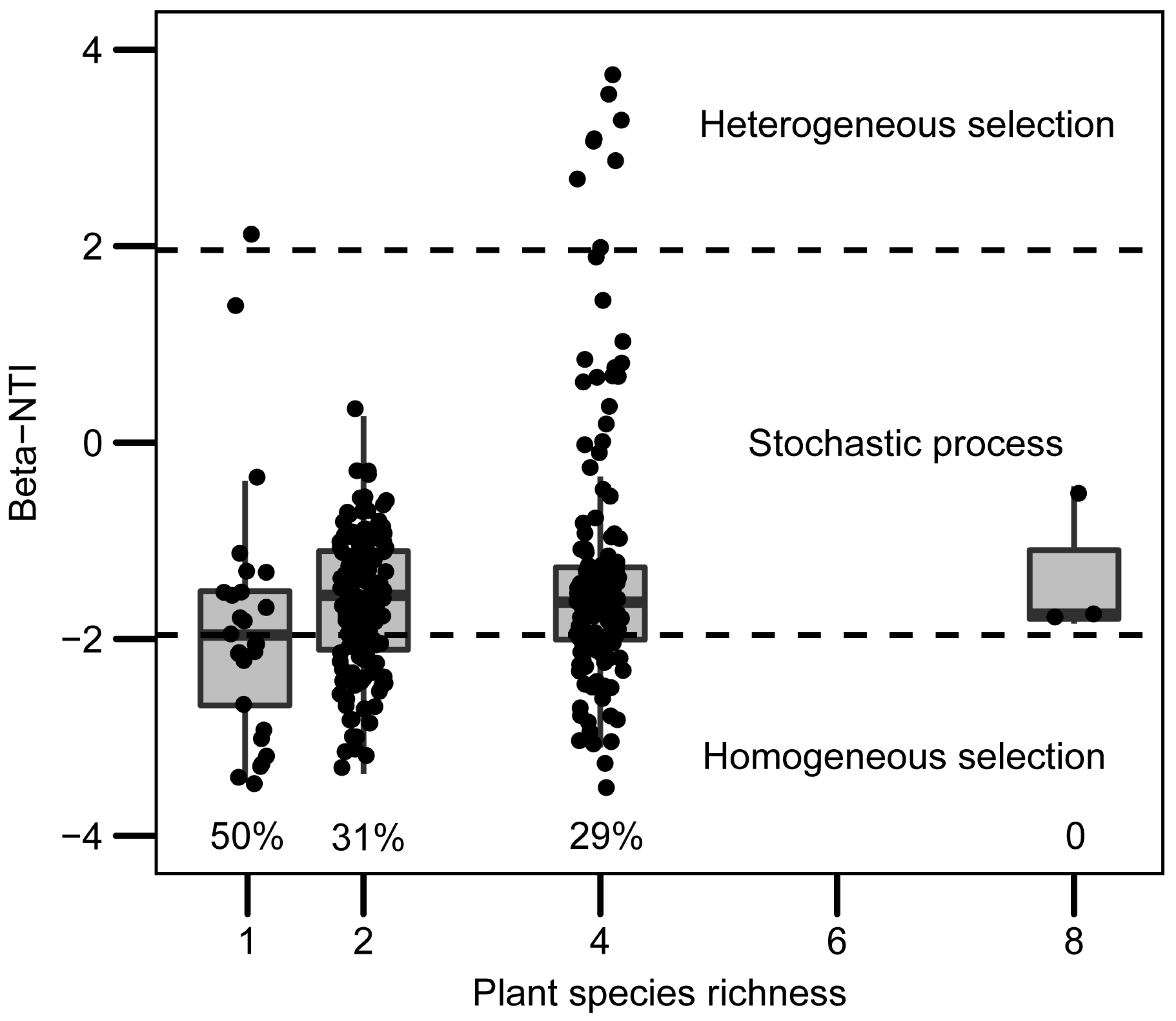
2.4. Community Assembly of Leaf Bacteria of the Invader
3. Discussion
4. Materials and Methods
4.1. Experiment Design
4.2. Sample Collection and Plant Biomass
4.3. Laboratory Analyses
4.4. Sequence Processing
4.5. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Akin-Fajiye, M.; Schmidt, A.C.; Fraser, L.H. Soil Nutrients and Variation in Biomass Rather than Native Species Richness Influence Introduced Plant Richness in a Semi-Arid Grassland. Basic Appl. Ecol. 2021, 53, 62–73. [Google Scholar] [CrossRef]
- Doherty, T.S.; Glen, A.S.; Nimmo, D.G.; Ritchie, E.G.; Dickman, C.R. Invasive Predators and Global Biodiversity Loss. Proc. Natl. Acad. Sci. USA 2016, 113, 11261–11265. [Google Scholar] [CrossRef] [Green Version]
- Tilman, D. The Ecological Consequences of Changes in Biodiversity: A Search for General Principles. Ecology 1999, 80, 1455–1474. [Google Scholar] [CrossRef]
- Zheng, Y.-L.; Burns, J.H.; Liao, Z.-Y.; Li, Y.; Yang, J.; Chen, Y.; Zhang, J.; Zheng, Y. Species Composition, Functional and Phylogenetic Distances Correlate with Success of Invasive Chromolaena Odorata in an Experimental Test. Ecol. Lett. 2018, 21, 1211–1220. [Google Scholar] [CrossRef] [PubMed]
- Beaury, E.M.; Finn, J.T.; Corbin, J.D.; Barr, V.; Bradley, B.A. Biotic Resistance to Invasion Is Ubiquitous across Ecosystems of the United States. Ecol. Lett. 2020, 23, 476–482. [Google Scholar] [CrossRef]
- Meyer, K.M.; Porch, R.; Muscettola, I.E.; Vasconcelos, A.L.S.; Sherman, J.K.; Metcalf, C.J.E.; Lindow, S.E.; Koskella, B. Plant Neighborhood Shapes Diversity and Reduces Interspecific Variation of the Phyllosphere Microbiome. ISME J. 2022, 16, 1376–1387. [Google Scholar] [CrossRef]
- Loreau, M. Biodiversity and Ecosystem Functioning: A Mechanistic Model. Proc. Natl. Acad. Sci. USA 1998, 95, 5632–5636. [Google Scholar] [CrossRef] [Green Version]
- Fargione, J.E.; Tilman, D. Diversity Decreases Invasion via Both Sampling and Complementarity Effects. Ecol. Lett. 2005, 8, 604–611. [Google Scholar] [CrossRef]
- Byun, C.; de Blois, S.; Brisson, J. Plant Functional Group Identity and Diversity Determine Biotic Resistance to Invasion by an Exotic Grass. J. Ecol. 2013, 101, 128–139. [Google Scholar] [CrossRef] [Green Version]
- Wardle, D.A. Experimental Demonstration That Plant Diversity Reduces Invasibility—Evidence of a Biological Mechanism or a Consequence of Sampling Effect? Oikos 2001, 95, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Hector, A.; Dobson, K.; Minns, A.; Bazeley-White, E.; Hartley Lawton, J. Community Diversity and Invasion Resistance: An Experimental Test in a Grassland Ecosystem and a Review of Comparable Studies. Ecol. Res. 2001, 16, 819–831. [Google Scholar] [CrossRef]
- Oakley, C.A.; Knox, J.S. Plant Species Richness Increases Resistance to Invasion by Non-Resident Plant Species during Grassland Restoration. Appl. Veg. Sci. 2013, 16, 21–28. [Google Scholar] [CrossRef]
- Davies, K.F.; Chesson, P.; Harrison, S.; Inouye, B.D.; Melbourne, B.A.; Rice, K.J. Spatial Heterogeneity Explains the Scale Dependence of the Native–Exotic Diversity Relationship. Ecology 2005, 86, 1602–1610. [Google Scholar] [CrossRef]
- Altieri, A.H.; van Wesenbeeck, B.K.; Bertness, M.D.; Silliman, B.R. Facilitation Cascade Drives Positive Relationship between Native Biodiversity and Invasion Success. Ecology 2010, 91, 1269–1275. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Qian, H.; Spyreas, G.; Crossland, M. Native-Exotic Species Richness Relationships across Spatial Scales and Biotic Homogenization in Wetland Plant Communities of Illinois, USA. Divers. Distrib. 2010, 16, 737–743. [Google Scholar] [CrossRef]
- Sax, D.F. Native and Naturalized Plant Diversity Are Positively Correlated in Scrub Communities of California and Chile. Divers. Distrib. 2002, 8, 193–210. [Google Scholar] [CrossRef]
- Zeiter, M.; Stampfli, A. Positive Diversity-Invasibility Relationship in Species-Rich Semi-Natural Grassland at the Neighbourhood Scale. Ann. Bot. 2012, 110, 1385–1393. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Morin, P.J. Productivity Gradients Cause Positive Diversity–Invasibility Relationships in Microbial Communities. Ecol. Lett. 2004, 7, 1047–1057. [Google Scholar] [CrossRef]
- Stohlgren, T.J.; Barnett, D.T.; Kartesz, J.T. The Rich Get Richer: Patterns of Plant Invasions in the United States. Front. Ecol. Environ. 2003, 1, 11–14. [Google Scholar] [CrossRef]
- Mitchell, C.E.; Agrawal, A.A.; Bever, J.D.; Gilbert, G.S.; Hufbauer, R.A.; Klironomos, J.N.; Maron, J.L.; Morris, W.F.; Parker, I.M.; Power, A.G.; et al. Biotic Interactions and Plant Invasions. Ecol. Lett. 2006, 9, 726–740. [Google Scholar] [CrossRef]
- Richardson, D.M.; Allsopp, N.; D’antonio, C.M.; Milton, S.J.; Rejmánek, M. Plant Invasions-the Role of Mutualisms. Biol. Rev. 2000, 75, 65–93. [Google Scholar] [CrossRef] [PubMed]
- Fridley, J.D.; Stachowicz, J.J.; Naeem, S.; Sax, D.F.; Seabloom, E.W.; Smith, M.D.; Stohlgren, T.J.; Tilman, D.; Holle, B.V. The Invasion Paradox: Reconciling Pattern and Process in Species Invasions. Ecology 2007, 88, 3–17. [Google Scholar] [CrossRef] [Green Version]
- Cavieres, L.A. Facilitation and the Invasibility of Plant Communities. J. Ecol. 2021, 109, 2019–2028. [Google Scholar] [CrossRef]
- Dong, L.-J.; Sun, Z.-K.; Gao, Y.; He, W.-M. Two-Year Interactions between Invasive Solidago canadensis and Soil Decrease Its Subsequent Growth and Competitive Ability. J. Plant Ecol. 2015, 8, 617–622. [Google Scholar] [CrossRef] [Green Version]
- Dong, L.-J.; Yu, H.-W.; He, W.-M. What Determines Positive, Neutral and Negative Impacts of Solidago canadensis Invasion on Native Plant Species Richness? Sci. Rep. 2015, 5, 16804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.-Y.; Gao, S.; Chen, T.; Wang, J.; Yu, F.-H. Interactions between Soil Microbes and Native Species Drive a Diversity-Invasibility Relationship. Biol. Invasions 2022, 1–2. [Google Scholar] [CrossRef]
- Bruno, J.F.; Stachowicz, J.J.; Bertness, M.D. Inclusion of Facilitation into Ecological Theory. Trends Ecol. Evol. 2003, 18, 119–125. [Google Scholar] [CrossRef]
- Sax, D.F.; Stachowicz, J.J.; Brown, J.H.; Bruno, J.F.; Dawson, M.N.; Gaines, S.D.; Grosberg, R.K.; Hastings, A.; Holt, R.D.; Mayfield, M.M.; et al. Ecological and Evolutionary Insights from Species Invasions. Trends Ecol. Evol. 2007, 22, 465–471. [Google Scholar] [CrossRef]
- Cavieres, L.A.; Quiroz, C.L.; Molina-Montenegro, M.A.; Muñoz, A.A.; Pauchard, A. Nurse Effect of the Native Cushion Plant Azorella monantha on the Invasive Non-Native Taraxacum officinale in the High-Andes of Central Chile. Perspect. Plant Ecol. Evol. Syst. 2005, 7, 217–226. [Google Scholar] [CrossRef]
- Zanini, L.; Ganade, G.; Hübel, I. Facilitation and Competition Influence Succession in a Subtropical Old Field. Plant Ecol. 2006, 185, 179–190. [Google Scholar] [CrossRef]
- Northfield, T.D.; Laurance, S.G.W.; Mayfield, M.M.; Paini, D.R.; Snyder, W.E.; Stouffer, D.B.; Wright, J.T.; Lach, L. Native Turncoats and Indirect Facilitation of Species Invasions. Proc. R. Soc. B Biol. Sci. 2018, 285, 20171936. [Google Scholar] [CrossRef] [Green Version]
- Sotomayor, D.A.; Lortie, C.J. Indirect Interactions in Terrestrial Plant Communities: Emerging Patterns and Research Gaps. Ecosphere 2015, 6, art103. [Google Scholar] [CrossRef]
- Shen, C.; Wang, J.; He, J.-Z.; Yu, F.-H.; Ge, Y. Plant Diversity Enhances Soil Fungal Diversity and Microbial Resistance to Plant Invasion. Appl. Environ. Microbiol. 2021, 87, e00251-21. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Wang, J.; Jing, Z.; Qiao, N.-H.; Xiong, C.; Ge, Y. Plant Diversity Enhances Soil Fungal Network Stability Indirectly through the Increase of Soil Carbon and Fungal Keystone Taxa Richness. Sci. Total Environ. 2022, 818, 151737. [Google Scholar] [CrossRef]
- Laforest-Lapointe, I.; Paquette, A.; Messier, C.; Kembel, S.W. Leaf Bacterial Diversity Mediates Plant Diversity and Ecosystem Function Relationships. Nature 2017, 546, 145–147. [Google Scholar] [CrossRef]
- Tao, J.; Meng, D.; Qin, C.; Liu, X.; Liang, Y.; Xiao, Y.; Liu, Z.; Gu, Y.; Li, J.; Yin, H. Integrated Network Analysis Reveals the Importance of Microbial Interactions for Maize Growth. Appl. Microbiol. Biotechnol. 2018, 102, 3805–3818. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Walder, F.; Büchi, L.; Meyer, M.; Held, A.Y.; Gattinger, A.; Keller, T.; Charles, R.; van der Heijden, M.G.A. Agricultural Intensification Reduces Microbial Network Complexity and the Abundance of Keystone Taxa in Roots. ISME J. 2019, 13, 1722–1736. [Google Scholar] [CrossRef] [Green Version]
- Xiong, C.; Zhu, Y.-G.; Wang, J.-T.; Singh, B.; Han, L.-L.; Shen, J.-P.; Li, P.-P.; Wang, G.-B.; Wu, C.-F.; Ge, A.-H.; et al. Host Selection Shapes Crop Microbiome Assembly and Network Complexity. New Phytol. 2021, 229, 1091–1104. [Google Scholar] [CrossRef]
- Bodenhausen, N.; Bortfeld-Miller, M.; Ackermann, M.; Vorholt, J.A. A Synthetic Community Approach Reveals Plant Genotypes Affecting the Phyllosphere Microbiota. PLoS Genet. 2014, 10, e1004283. [Google Scholar] [CrossRef] [Green Version]
- Zamioudis, C.; Pieterse, C.M.J. Modulation of Host Immunity by Beneficial Microbes. Mol. Plant-Microbe Interact. MPMI 2012, 25, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Purahong, W.; Orrù, L.; Donati, I.; Perpetuini, G.; Cellini, A.; Lamontanara, A.; Michelotti, V.; Tacconi, G.; Spinelli, F. Plant Microbiome and Its Link to Plant Health: Host Species, Organs and Pseudomonas syringae pv. actinidiae Infection Shaping Bacterial Phyllosphere Communities of Kiwifruit Plants. Front. Plant Sci. 2018, 9, 1563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moyes, A.B.; Kueppers, L.M.; Pett-Ridge, J.; Carper, D.L.; Vandehey, N.; O’Neil, J.; Frank, A.C. Evidence for Foliar Endophytic Nitrogen Fixation in a Widely Distributed Subalpine Conifer. New Phytol. 2016, 210, 657–668. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Stirling, E.; Xie, H.; Li, W.; Lv, X.; Matsumoto, H.; Cheng, H.; Xu, A.; Lai, W.; Wang, Y.; et al. Continental Scale Deciphering of Microbiome Networks Untangles the Phyllosphere Homeostasis in Tea Plant. J. Adv. Res. 2023, 44, 13–22. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, X.; Zhu, Y.; Li, L.; Zhang, Y.; Li, J.; Song, X.; Qiang, S. Biological Control of Solidago canadensis Using a Bioherbicide Isolate of Sclerotium rolfsii SC64 Increased the Biodiversity in Invaded Habitats. Biol. Control 2019, 139, 104093. [Google Scholar] [CrossRef]
- Dong, L.-J.; Ma, L.-N.; He, W.-M. Arbuscular Mycorrhizal Fungi Help Explain Invasion Success of Solidago canadensis. Appl. Soil Ecol. 2021, 157, 103763. [Google Scholar] [CrossRef]
- Wang, C.; Cheng, H.; Wang, S.; Wei, M.; Du, D. Plant Community and the Influence of Plant Taxonomic Diversity on Community Stability and Invasibility: A Case Study Based on Solidago canadensis L. Sci. Total Environ. 2021, 768, 144518. [Google Scholar] [CrossRef]
- Li, S.-P.; Jia, P.; Fan, S.-Y.; Wu, Y.-T.; Liu, X.; Meng, Y.; Li, Y.; Shu, W.; Li, J.; Jiang, L. Functional Traits Explain the Consistent Resistance of Biodiversity to Plant Invasion under Nitrogen Enrichment. Ecol. Lett. 2021, 25, 778–789. [Google Scholar] [CrossRef]
- Kennedy, T.A.; Naeem, S.; Howe, K.M.; Knops, J.M.H.; Tilman, D.; Reich, P. Biodiversity as a Barrier to Ecological Invasion. Nature 2002, 417, 636–638. [Google Scholar] [CrossRef]
- Case, T.J. Invasion Resistance Arises in Strongly Interacting Species-Rich Model Competition Communities. Proc. Natl. Acad. Sci. USA 1990, 87, 9610–9614. [Google Scholar] [CrossRef] [Green Version]
- Stohlgren, T.J.; Binkley, D.; Chong, G.W.; Kalkhan, M.A.; Schell, L.D.; Bull, K.A.; Otsuki, Y.; Newman, G.; Bashkin, M.; Son, Y. Exotic Plant Species Invade Hot Spots of Native Plant Diversity. Ecol. Monogr. 1999, 69, 25–46. [Google Scholar] [CrossRef]
- Brown, R.L.; Peet, R.K. Diversity and Invasibility of Southern Appalachian Plant Communities. Ecology 2003, 84, 32–39. [Google Scholar] [CrossRef]
- Davies, K.F.; Harrison, S.; Safford, H.D.; Viers, J.H. Productivity Alters the Scale Dependence of the Diversity–Invasibility Relationship. Ecology 2007, 88, 1940–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Q. No Consistent Small-Scale Native–Exotic Relationships. Plant Ecol. 2015, 216, 1225–1230. [Google Scholar] [CrossRef]
- Tomasetto, F.; Duncan, R.P.; Hulme, P.E. Resolving the Invasion Paradox: Pervasive Scale and Study Dependence in the Native-Alien Species Richness Relationship. Ecol. Lett. 2019, 22, 1038–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, M.A.; Grime, J.P.; Thompson, K. Fluctuating Resources in Plant Communities: A General Theory of Invasibility. J. Ecol. 2000, 88, 528–534. [Google Scholar] [CrossRef] [Green Version]
- Bulleri, F.; Bruno, J.F.; Benedetti-Cecchi, L. Beyond Competition: Incorporating Positive Interactions between Species to Predict Ecosystem Invasibility. PLoS Biol. 2008, 6, e162. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; van Themaat, E.V.L.; Schulze-Lefert, P. Structure and Functions of the Bacterial Microbiota of Plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [Green Version]
- Horner-Devine, M.C.; Lage, M.; Hughes, J.B.; Bohannan, B.J.M. A Taxa–Area Relationship for Bacteria. Nature 2004, 432, 750–753. [Google Scholar] [CrossRef]
- Bai, Y.; Müller, D.B.; Srinivas, G.; Garrido-Oter, R.; Potthoff, E.; Rott, M.; Dombrowski, N.; Münch, P.C.; Spaepen, S.; Remus-Emsermann, M.; et al. Functional Overlap of the Arabidopsis Leaf and Root Microbiota. Nature 2015, 528, 364–369. [Google Scholar] [CrossRef]
- Simberloff, D.; Von Holle, B. Positive Interactions of Nonindigenous Species: Invasional Meltdown? Biol. Invasions 1999, 1, 21–32. [Google Scholar] [CrossRef]
- Callaway, R.M.; Brooker, R.W.; Choler, P.; Kikvidze, Z.; Lortie, C.J.; Michalet, R.; Paolini, L.; Pugnaire, F.I.; Newingham, B.; Aschehoug, E.T.; et al. Positive Interactions among Alpine Plants Increase with Stress. Nature 2002, 417, 844–848. [Google Scholar] [CrossRef]
- Qi, G.; Ma, G.; Chen, S.; Lin, C.; Zhao, X. Microbial Network and Soil Properties Are Changed in Bacterial Wilt-Susceptible Soil. Appl. Environ. Microbiol. 2019, 85, e00162-19. [Google Scholar] [CrossRef] [Green Version]
- Palmer, M.; Steenkamp, E.T.; Coetzee, M.P.A.; Avontuur, J.R.; Chan, W.-Y.; van Zyl, E.; Blom, J.; Venter, S.N. Mixta Gen. Nov., a New Genus in the Erwiniaceae. Int. J. Syst. Evol. Microbiol. 2018, 68, 1396–1407. [Google Scholar] [CrossRef] [PubMed]
- Romero, F.M.; Marina, M.; Pieckenstain, F.L. The Communities of Tomato (Solanum lycopersicum L.) Leaf Endophytic Bacteria, Analyzed by 16S-Ribosomal RNA Gene Pyrosequencing. FEMS Microbiol. Lett. 2014, 351, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Ottesen, A.R.; González Peña, A.; White, J.R.; Pettengill, J.B.; Li, C.; Allard, S.; Rideout, S.; Allard, M.; Hill, T.; Evans, P.; et al. Baseline Survey of the Anatomical Microbial Ecology of an Important Food Plant: Solanum lycopersicum (Tomato). BMC Microbiol. 2013, 13, 114. [Google Scholar] [CrossRef] [Green Version]
- Cheng, D.; Tian, Z.; Feng, L.; Xu, L.; Wang, H. Diversity Analysis of the Rhizospheric and Endophytic Bacterial Communities of Senecio Vulgaris L. (Asteraceae) in an Invasive Range. PeerJ 2019, 6, e6162. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Wei, Z.; Weidner, S.; Friman, V.-P.; Xu, Y.-C.; Shen, Q.-R.; Jousset, A. Probiotic Pseudomonas Communities Enhance Plant Growth and Nutrient Assimilation via Diversity-Mediated Ecosystem Functioning. Soil Biol. Biochem. 2017, 113, 122–129. [Google Scholar] [CrossRef]
- Santillan, E.; Seshan, H.; Constancias, F.; Drautz-Moses, D.I.; Wuertz, S. Frequency of Disturbance Alters Diversity, Function, and Underlying Assembly Mechanisms of Complex Bacterial Communities. Npj. Biofilms Microbiomes 2019, 5, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shade, A. Diversity Is the Question, Not the Answer. ISME J. 2017, 11, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Stewart, W.D. Biological and Ecological Aspects of Nitrogen Fixation by Free-Living Micro-Organisms. Proc. R. Soc. Lond. B Biol. Sci. 1969, 172, 367–388. [Google Scholar] [CrossRef]
- Bodenhausen, N.; Horton, M.W.; Bergelson, J. Bacterial Communities Associated with the Leaves and the Roots of Arabidopsis thaliana. PLoS ONE 2013, 8, e56329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘Vegan.’ Community Ecology Package, Version. 2013, Volume 2, pp. 1–295. Available online: https://github.com/vegandevs/vegan (accessed on 26 May 2022).
- Fine, P.V.A.; Kembel, S.W. Phylogenetic Community Structure and Phylogenetic Turnover across Space and Edaphic Gradients in Western Amazonian Tree Communities. Ecography 2011, 34, 552–565. [Google Scholar] [CrossRef]
- Stegen, J.C.; Lin, X.; Fredrickson, J.K.; Chen, X.; Kennedy, D.W.; Murray, C.J.; Rockhold, M.L.; Konopka, A. Quantifying Community Assembly Processes and Identifying Features That Impose Them. ISME J. 2013, 7, 2069–2079. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, X.-D.; Wang, J.; Shen, C.; Wang, J.; Jing, Z.; Huang, L.-N.; Luo, Z.-H.; Ge, Y. Increased Leaf Bacterial Network Complexity along the Native Plant Diversity Gradient Facilitates Plant Invasion? Plants 2023, 12, 1406. https://doi.org/10.3390/plants12061406
Du X-D, Wang J, Shen C, Wang J, Jing Z, Huang L-N, Luo Z-H, Ge Y. Increased Leaf Bacterial Network Complexity along the Native Plant Diversity Gradient Facilitates Plant Invasion? Plants. 2023; 12(6):1406. https://doi.org/10.3390/plants12061406
Chicago/Turabian StyleDu, Xiang-Deng, Jiang Wang, Congcong Shen, Jichen Wang, Zhongwang Jing, Li-Nan Huang, Zhen-Hao Luo, and Yuan Ge. 2023. "Increased Leaf Bacterial Network Complexity along the Native Plant Diversity Gradient Facilitates Plant Invasion?" Plants 12, no. 6: 1406. https://doi.org/10.3390/plants12061406