
Transgenerational Herbivory Effects on Performance of Clonal Offspring of the Invasive Plant Alternanthera philoxeroides
 and
and
Abstract
:1. Introduction
2. Results
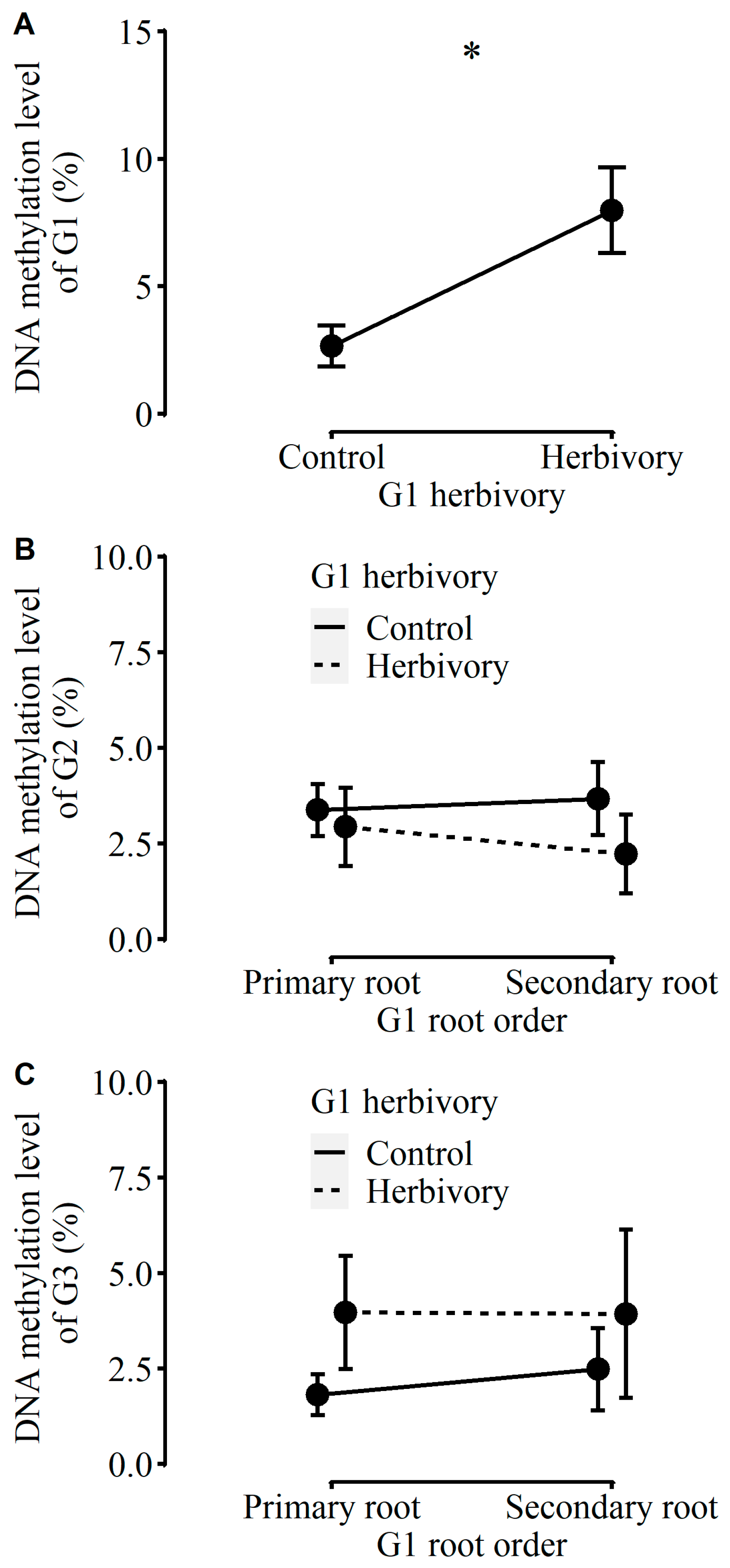
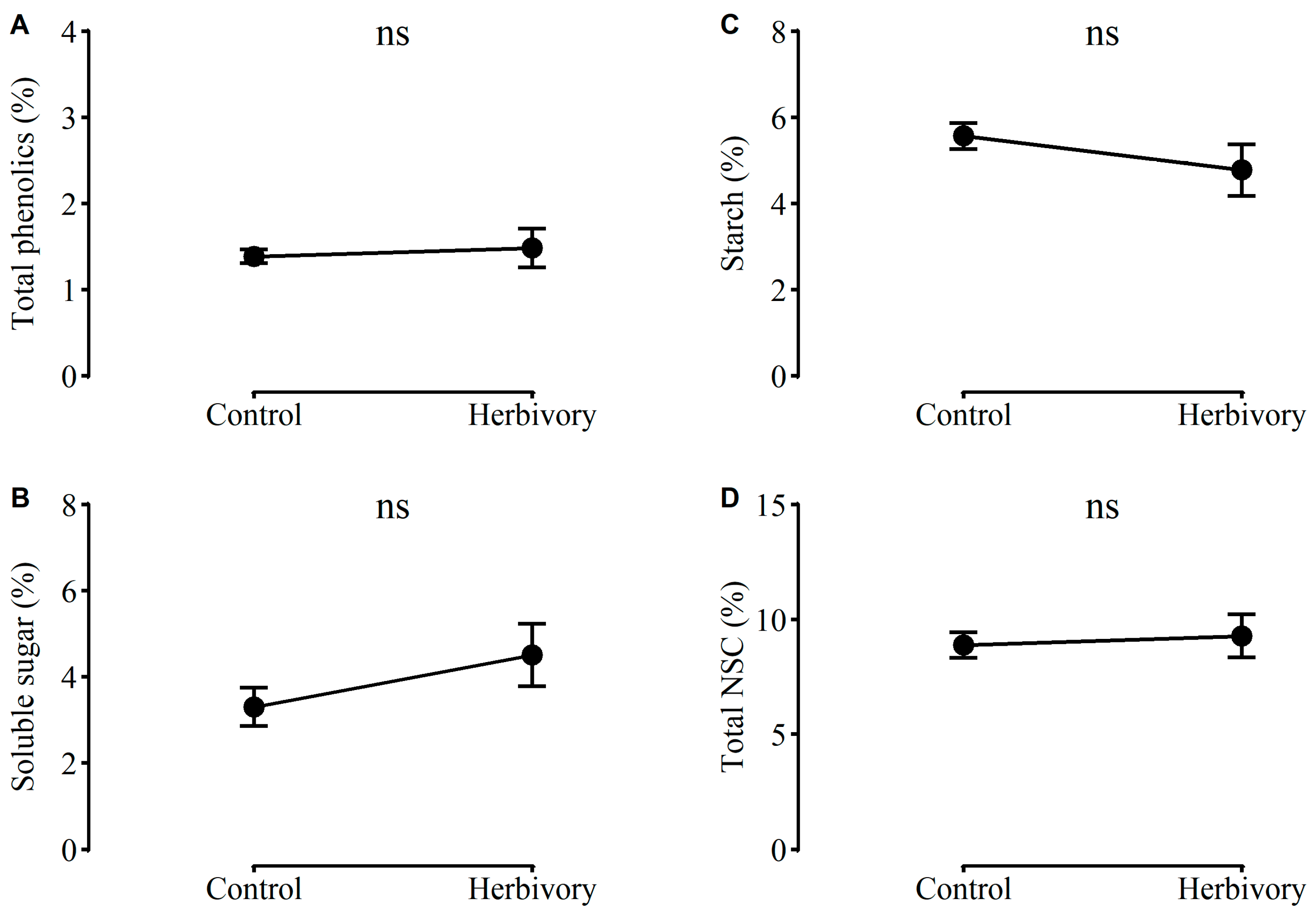
2.1. Performance of Plants in G1
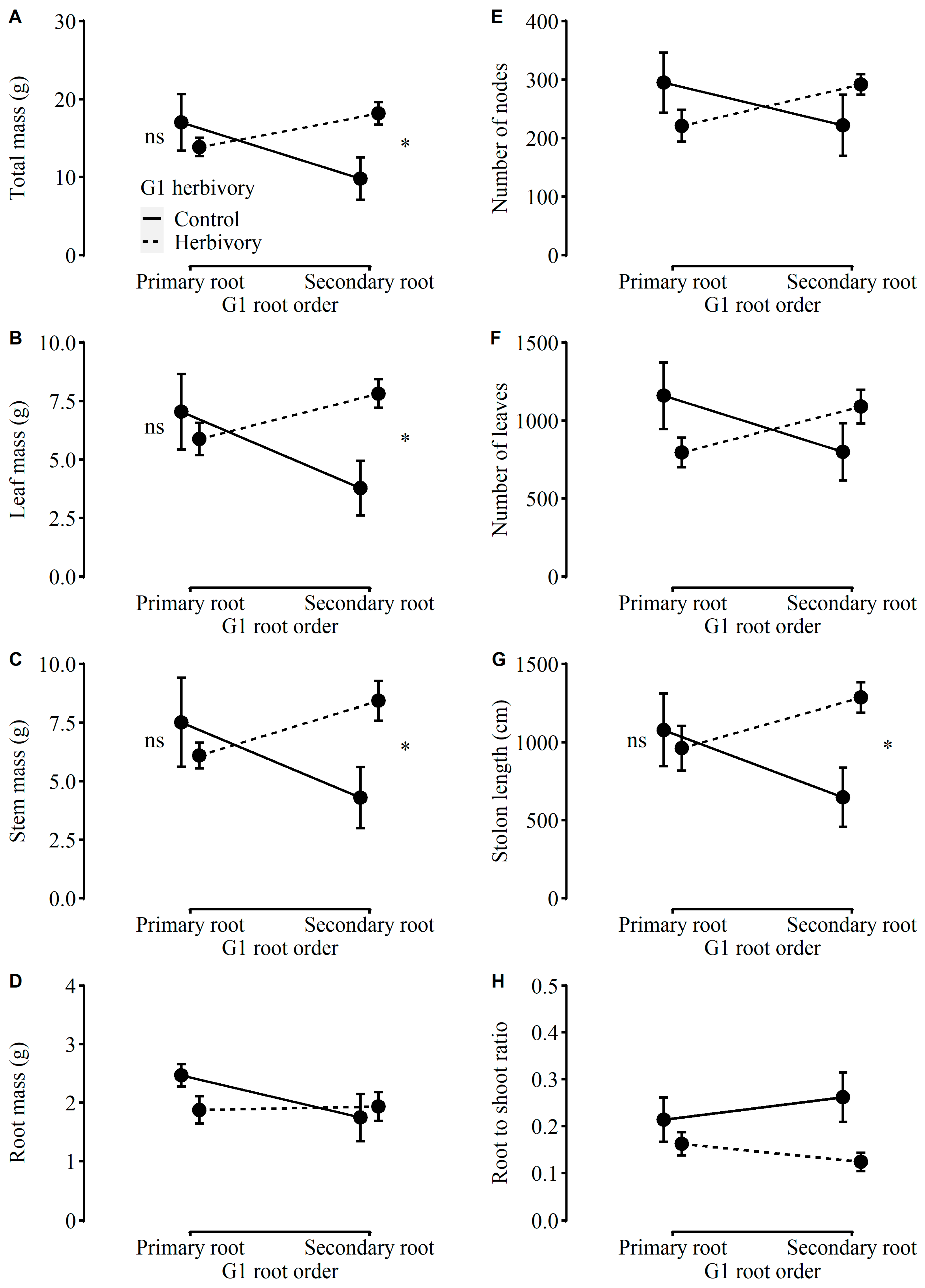
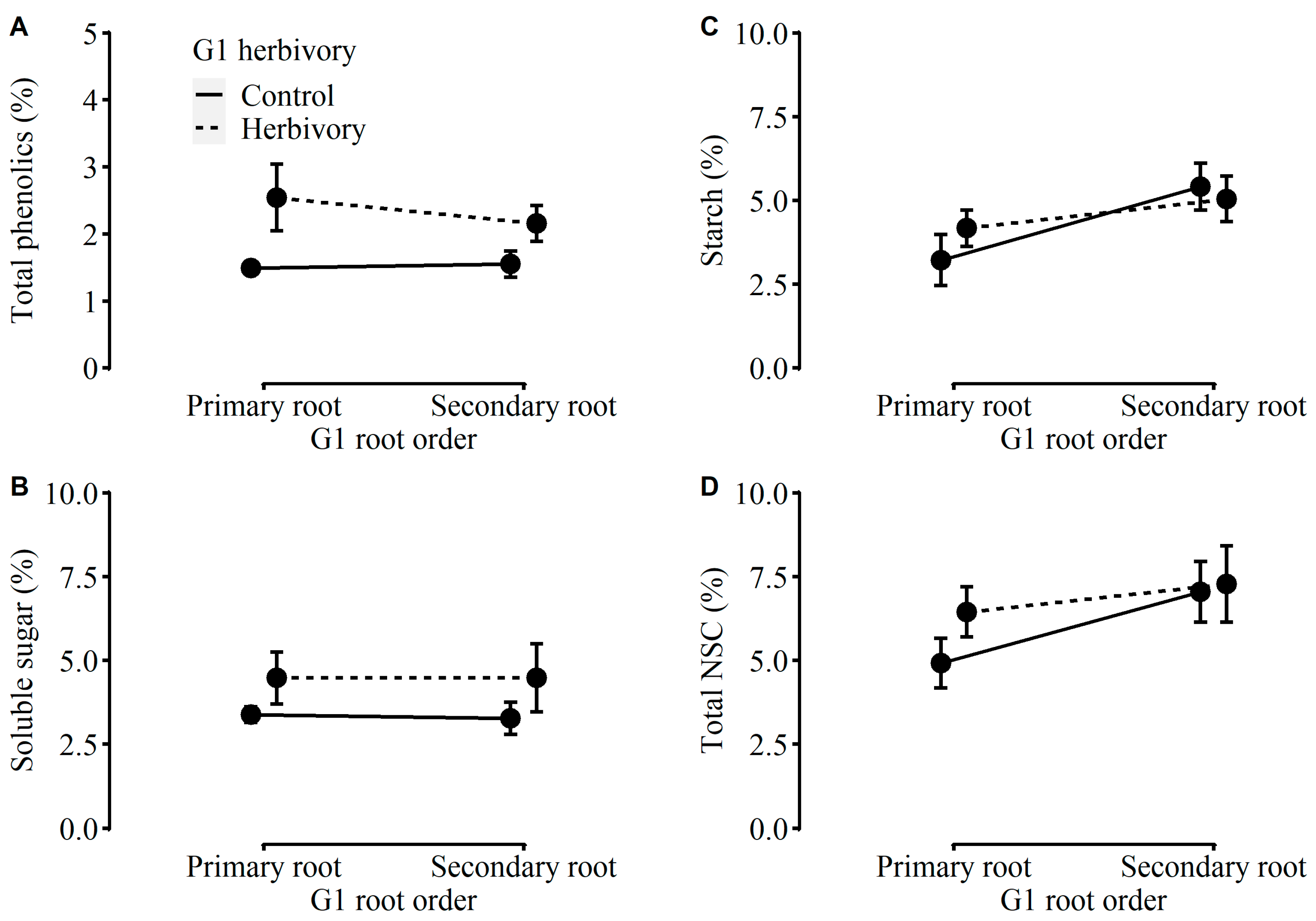
2.2. Performance of Plants in G2
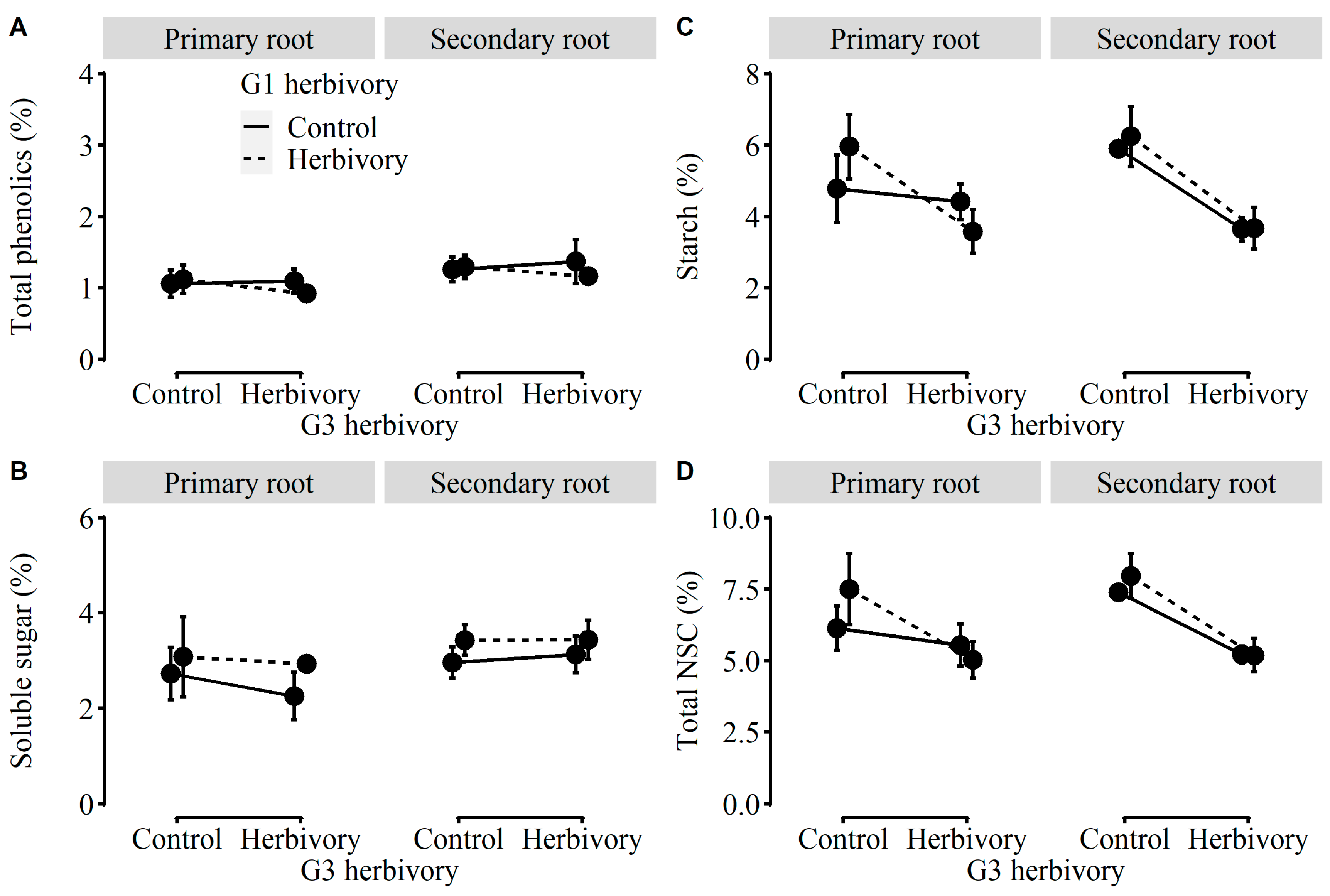
2.3. Performance of Plants in G3
3. Discussion
3.1. Direct Herbivory Effects in G1 and G3
3.2. Transgenerational Effects in G2 and G3
3.3. Role of DNA Methylation
4. Materials and Methods
4.1. Study Species
4.2. Experimental Design
4.3. Measurements
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Traits | Physiological Traits | |||
|---|---|---|---|---|
| F | p | F | p | |
| G1 herbivory (G1H) | 5.91 | 0.006 | 1.41 | 0.305 |
| G1 root order (GR) | 1.80 | 0.188 | 2.91 | 0.084 |
| G1H × GR | 1.57 | 0.244 | 0.82 | 0.546 |
| Growth Traits | Physiological Traits | |||
|---|---|---|---|---|
| F | p | F | p | |
| G1 herbivory (G1H) | 1.13 | 0.364 | 0.67 | 0.616 |
| G1 root order (GR) | 0.99 | 0.446 | 0.99 | 0.431 |
| G3 herbivory (G3H) | 17.77 | <0.001 | 4.44 | 0.006 |
| G1H × GR | 0.90 | 0.507 | 0.33 | 0.855 |
| G1H × G3H | 0.85 | 0.536 | 0.93 | 0.463 |
| GR × G3H | 1.69 | 0.148 | 0.73 | 0.582 |
| G1H × GR × G3H | 0.61 | 0.723 | 0.43 | 0.783 |



References
- Mitchell, C.E.; Agrawal, A.A.; Bever, J.D.; Gilbert, G.S.; Hufbauer, R.A.; Klironomos, J.N.; Maron, J.L.; Morris, W.F.; Parker, I.M.; Power, A.G.; et al. Biotic Interactions and Plant Invasions. Ecol. Lett. 2006, 9, 726–740. [Google Scholar] [CrossRef]
- Dai, Z.-C.; Wan, L.-Y.; Qi, S.-S.; Rutherford, S.; Ren, G.-Q.; Wan, J.S.H.; Du, D.-L. Synergy among Hypotheses in the Invasion Process of Alien Plants: A Road Map within a Timeline. Perspect. Plant Ecol. Evol. Syst. 2020, 47, 125575. [Google Scholar] [CrossRef]
- Enders, M.; Havemann, F.; Ruland, F.; Bernard-Verdier, M.; Catford, J.A.; Gomez-Aparicio, L.; Haider, S.; Heger, T.; Kueffer, C.; Kuehn, I.; et al. A Conceptual Map of Invasion Biology: Integrating Hypotheses into a Consensus Network. Glob. Ecol. Biogeogr. 2020, 29, 978–991. [Google Scholar] [CrossRef] [Green Version]
- Keane, R.M.; Crawley, M.J. Exotic Plant Invasions and the Enemy Release Hypothesis. Trends Ecol. Evol. 2002, 17, 164–170. [Google Scholar] [CrossRef]
- Costan, C.-A.; Godsoe, W.K.; Bufford, J.L.; Marris, J.W.M.; Hulme, P.E. Can the Enemy Release Hypothesis Explain the Success of Rumex (Polygonaceae) Species in an Introduced Range? Biol. Invasions 2022, 24, 2665–2677. [Google Scholar] [CrossRef]
- Lucero, J.E.; Schaffner, U.; Asadi, G.; Bagheri, A.; Rajabov, T.; Callaway, R.M. Enemy Release from the Effects of Generalist Granivores Can Facilitate Bromus Tectorum Invasion in the Great Basin Desert. Ecol. Evol. 2019, 9, 8490–8499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Mu, X.-D.; Zhang, S.; Dick, J.T.A.; Zhu, B.-T.; Gu, D.-G.; Yang, Y.-X.; Luo, D.; Hu, Y.-C. A Global Analysis of Enemy Release and Its Variation with Latitude. Glob. Ecol. Biogeogr. 2021, 30, 277–288. [Google Scholar] [CrossRef]
- Colautti, R.I.; Ricciardi, A.; Grigorovich, I.A.; MacIsaac, H.J. Is Invasion Success Explained by the Enemy Release Hypothesis? Ecol. Lett. 2004, 7, 721–733. [Google Scholar] [CrossRef]
- Liu, H.; Stiling, P. Testing the Enemy Release Hypothesis: A Review and Meta-Analysis. Biol. Invasions 2006, 8, 1535–1545. [Google Scholar] [CrossRef]
- Meijer, K.; Schilthuizen, M.; Beukeboom, L.; Smit, C. A Review and Meta-Analysis of the Enemy Release Hypothesis in Plant–Herbivorous Insect Systems. PeerJ 2016, 4, e2778. [Google Scholar] [CrossRef] [Green Version]
- Chun, Y.-J.; van Kleunen, M.; Dawson, W. The Role of Enemy Release, Tolerance and Resistance in Plant Invasions: Linking Damage to Performance. Ecol. Lett. 2010, 13, 937–946. [Google Scholar] [CrossRef]
- Schoonhoven, L.M.; van Loon, J.J.A.; Dicke, M. Insect-Plant Biology, 2nd ed.; Oxford University Press: Oxford, UK, 2005; ISBN 978-0-19-852595-0. [Google Scholar]
- Strauss, S.Y.; Agrawal, A.A. The Ecology and Evolution of Plant Tolerance to Herbivory. Trends Ecol. Evol. 1999, 14, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Stowe, K.A.; Marquis, R.; Hochwender, C.; Simms, E. The Evolutionary Ecology of Tolerance to Consumer Damage. Annu. Rev. Ecol. Evol. Syst. 2000, 31, 565–595. [Google Scholar] [CrossRef]
- Boege, K.; Marquis, R.J. Facing Herbivory as You Grow up: The Ontogeny of Resistance in Plants. Trends Ecol. Evol. 2005, 20, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.-C.; Wang, M.-Z.; Liu, R.-H.; Luo, F.-L.; Li, H.-L.; Yu, F.-H. Direct and Legacy Effects of Herbivory on Growth and Physiology of a Clonal Plant. Biol. Invasions 2018, 20, 3631–3645. [Google Scholar] [CrossRef]
- Dong, B.-C.; Alpert, P.; Yu, F.-H. Transgenerational Effects of Herbivory and Soil Nutrients Transmitted via Vegetative Reproduction in the Clonal Plant Alternanthera philoxeroides. Perspect. Plant Ecol. Evol. Syst. 2019, 41, 125498. [Google Scholar] [CrossRef]
- Agrawal, A.A.; Weber, M.G. On the Study of Plant Defence and Herbivory Using Comparative Approaches: How Important Are Secondary Plant Compounds. Ecol. Lett. 2015, 18, 985–991. [Google Scholar] [CrossRef]
- War, A.R.; Taggar, G.K.; Hussain, B.; Taggar, M.S.; Nair, R.M.; Sharma, H.C. Plant Defense against Herbivory and Insect Adaptations. AoB Plants 2018, 10, ply037. [Google Scholar] [CrossRef]
- Pearse, I.S.; Hipp, A.L. Phylogenetic and Trait Similarity to a Native Species Predict Herbivory on Non-Native Oaks. Proc. Natl. Acad. Sci. USA 2009, 106, 18097–18102. [Google Scholar] [CrossRef] [Green Version]
- Tanentzap, A.J.; Lee, W.G.; Dugdale, J.S.; Patrick, B.P.; Fenner, M.; Walker, S.; Coomes, D.A. Differential Responses of Vertebrate and Invertebrate Herbivores to Traits of New Zealand Subalpine Shrubs. Ecology 2011, 92, 994–999. [Google Scholar] [CrossRef]
- Huang, W.; Siemann, E.; Xiao, L.; Yang, X.-F.; Ding, J.-Q. Species-Specific Defence Responses Facilitate Conspecifics and Inhibit Heterospecifics in above–Belowground Herbivore Interactions. Nat. Commun. 2014, 5, 4851. [Google Scholar] [CrossRef] [Green Version]
- Steets, J.A.; Ashman, T. Maternal Effects of Herbivory in Impatiens capensis. Int. J. Plant Sci. 2010, 171, 509–518. [Google Scholar] [CrossRef]
- Rasmann, S.; De Vos, M.; Casteel, C.L.; Tian, D.; Halitschke, R.; Sun, J.Y.; Agrawal, A.A.; Felton, G.W.; Jander, G. Herbivory in the Previous Generation Primes Plants for Enhanced Insect Resistance. Plant Physiol. 2012, 158, 854–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, B.-C.; Fu, T.; Luo, F.-L.; Yu, F.-H. Herbivory-Induced Maternal Effects on Growth and Defense Traits in the Clonal Species Alternanthera philoxeroides. Sci. Total Environ. 2017, 605, 114–123. [Google Scholar] [CrossRef]
- Kellenberger, R.T.; Desurmont, G.A.; Schlüter, P.M.; Schiestl, F.P. Trans-Generational Inheritance of Herbivory-Induced Phenotypic Changes in Brassica rapa. Sci. Rep. 2018, 8, 3536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kafle, D.; Wurst, S. Legacy Effects of Herbivory Enhance Performance and Resistance of Progeny Plants. J. Ecol. 2019, 107, 58–68. [Google Scholar] [CrossRef]
- Nihranz, C.T.; Kolstrom, R.L.; Kariyat, R.R.; Mescher, M.C.; De Moraes, C.M.; Stephenson, A.G. Herbivory and Inbreeding Affect Growth, Reproduction, and Resistance in the Rhizomatous Offshoots of Solanum Carolinense (Solanaceae). Evol. Ecol. 2019, 33, 499–520. [Google Scholar] [CrossRef] [Green Version]
- Nihranz, C.T.; Walker, W.S.; Brown, S.J.; Mescher, M.C.; De Moraes, C.M.; Stephenson, A.G. Transgenerational Impacts of Herbivory and Inbreeding on Reproductive Output in Solanum carolinense. Am. J. Bot. 2020, 107, 286–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobral, M.; Sampedro, L.; Neylan, I.; Siemens, D.; Dirzo, R. Phenotypic Plasticity in Plant Defense across Life Stages: Inducibility, Transgenerational Induction, and Transgenerational Priming in Wild Radish. Proc. Natl. Acad. Sci. USA 2021, 118, e2005865118. [Google Scholar] [CrossRef] [PubMed]
- Uller, T.; Nakagawa, S.; English, S. Weak Evidence for Anticipatory Parental Effects in Plants and Animals. J. Evol. Biol. 2013, 26, 2161–2170. [Google Scholar] [CrossRef]
- Engqvist, L.; Reinhold, K. Adaptive Trans-Generational Phenotypic Plasticity and the Lack of an Experimental Control in Reciprocal Match/Mismatch Experiments. Methods Ecol. Evol. 2016, 7, 1482–1488. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.-J.; Zhou, M.; Lin, Z.; Li, Q.-S.Q.; Zhang, Y.-Y. Transgenerational Effects Benefit Offspring across Diverse Environments: A Meta-Analysis in Plants and Animals. Ecol. Lett. 2019, 22, 1976–1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, A.M.; Hellmann, J.K. An Integrative Framework for Understanding the Mechanisms and Multigenerational Consequences of Transgenerational Plasticity. Annu. Rev. Ecol. Evol. Syst. 2019, 50, 97–118. [Google Scholar] [CrossRef]
- Richards, C.L.; Alonso, C.; Becker, C.; Bossdorf, O.; Bucher, E.; Colomé-Tatché, M.; Durka, W.; Engelhardt, J.; Gaspar, B.; Gogol-Döring, A.; et al. Ecological Plant Epigenetics: Evidence from Model and Non-Model Species, and the Way Forward. Ecol. Lett. 2017, 20, 1576–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, C.; Ramos-Cruz, D.; Becker, C. The Role of Plant Epigenetics in Biotic Interactions. New Phytol. 2019, 221, 731–737. [Google Scholar] [CrossRef]
- Richards, E.J. Inherited Epigenetic Variation—Revisiting Soft Inheritance. Nat. Rev. Genet. 2006, 7, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Bossdorf, O.; Richards, C.L.; Pigliucci, M. Epigenetics for Ecologists. Ecol. Lett. 2008, 11, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Jablonka, E.; Raz, G. Transgenerational Epigenetic Inheritance: Prevalence, Mechanisms, and Implications for the Study of Heredity and Evolution. Q. Rev. Biol. 2009, 84, 131–176. [Google Scholar] [CrossRef] [Green Version]
- Herman, J.J.; Spencer, H.G.; Donohue, K.; Sultan, S.E. How Stable ‘Should’ Epigenetic Modifications Be? Insights from Adaptive Plasticity and Bet Hedging. Evolution 2014, 68, 632–643. [Google Scholar] [CrossRef] [Green Version]
- Thiebaut, F.; Hemerly, A.S.; Ferreira, P.C.G. A Role for Epigenetic Regulation in the Adaptation and Stress Responses of Non-Model Plants. Front. Plant Sci. 2019, 10, 246. [Google Scholar] [CrossRef] [Green Version]
- Herrera, C.M.; Bazaga, P. Untangling Individual Variation in Natural Populations: Ecological, Genetic and Epigenetic Correlates of Long-Term Inequality in Herbivory. Mol. Ecol. 2011, 20, 1675–1688. [Google Scholar] [CrossRef]
- Kellenberger, R.T.; Schlüter, P.M.; Schiestl, F.P. Herbivore-Induced DNA Demethylation Changes Floral Signalling and Attractiveness to Pollinators in Brassica rapa. PLoS ONE 2016, 11, e0166646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mounger, J.; Ainouche, M.L.; Bossdorf, O.; Cavé-Radet, A.; Li, B.; Parepa, M.; Salmon, A.; Yang, J.; Richards, C.L. Epigenetics and the Success of Invasive Plants. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20200117. [Google Scholar] [CrossRef]
- Latzel, V.; Klimešová, J. Transgenerational Plasticity in Clonal Plants. Evol. Ecol. 2010, 24, 1537–1543. [Google Scholar] [CrossRef]
- Douhovnikoff, V.; Dodd, R.S. Epigenetics: A Potential Mechanism for Clonal Plant Success. Plant Ecol. 2015, 216, 227–233. [Google Scholar] [CrossRef]
- Klimešová, J.; Klimes, L. Clonal Growth Diversity and Bud Banks of Plants in the Czech Flora: An Evaluation Using the CLO-PLA3 Database. Preslia 2008, 80, 255–275. [Google Scholar]
- Klimešová, J.; Martínková, J.; Herben, T. Horizontal Growth: An Overlooked Dimension in Plant Trait Space. Perspect. Plant Ecol. Evol. Syst. 2018, 32, 18–21. [Google Scholar] [CrossRef]
- Ott, J.P.; Klimešová, J.; Hartnett, D.C. The Ecology and Significance of Below-Ground Bud Banks in Plants. Ann. Bot. 2019, 123, 1099–1118. [Google Scholar] [CrossRef]
- Martínková, J.; Klimešová, J. Enforced Clonality Confers a Fitness Advantage. Front. Plant Sci. 2016, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.-L.; Helmisaari, H.-S.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B.; et al. Redefining Fine Roots Improves Understanding of Below-Ground Contributions to Terrestrial Biosphere Processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef]
- Kong, D.; Wang, J.; Valverde-Barrantes, O.J.; Kardol, P. A Framework to Assess the Carbon Supply–Consumption Balance in Plant Roots. New Phytol. 2021, 229, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Guo, D.-L.; Xia, M.-X.; Wei, X.; Chang, W.-J.; Liu, Y.; Wang, Z.-Q. Anatomical Traits Associated with Absorption and Mycorrhizal Colonization Are Linked to Root Branch Order in Twenty-Three Chinese Temperate Tree Species. New Phytol. 2008, 180, 673–683. [Google Scholar] [CrossRef]
- Yu, H.; Fan, S. Differences in Physiological Traits and Resistances of Alternanthera philoxeroides after Herbivory by Generalists and Specialists. Aquat. Ecol. 2018, 52, 323–332. [Google Scholar] [CrossRef]
- Hu, X.-T.; Dong, B.-C. Herbivory and Nitrogen Availability Affect Performance of an Invader Alternanthera philoxeroides and Its Native Congener A. sessilis. Flora 2019, 257, 151412. [Google Scholar] [CrossRef]
- Wilson, J.R.U.; Yeates, A.; Schooler, S.; Julien, M.H. Rapid Response to Shoot Removal by the Invasive Wetland Plant, Alligator Weed (Alternanthera philoxeroides). Environ. Exp. Bot. 2007, 60, 20–25. [Google Scholar] [CrossRef]
- Jia, X.; Pan, X.-Y.; Li, B.; Chen, J.-K.; Yang, X.-Z. Allometric Growth, Disturbance Regime, and Dilemmas of Controlling Invasive Plants: A Model Analysis. Biol. Invasions 2009, 11, 743–752. [Google Scholar] [CrossRef]
- Heinze, J. Herbivory by Aboveground Insects Impacts Plant Root Morphological Traits. Plant Ecol. 2020, 221, 725–732. [Google Scholar] [CrossRef]
- Ashton, I.W.; Lerdau, M.T. Tolerance to Herbivory, and Not Resistance, May Explain Differential Success of Invasive, Naturalized, and Native North American Temperate Vines. Divers. Distrib. 2008, 14, 169–178. [Google Scholar] [CrossRef]
- Huang, W.; Siemann, E.; Wheeler, G.S.; Zou, J.-W.; Carrillo, J.-L.; Ding, J.-Q. Resource Allocation to Defence and Growth Are Driven by Different Responses to Generalist and Specialist Herbivory in an Invasive Plant. J. Ecol. 2010, 98, 1157–1167. [Google Scholar] [CrossRef]
- Liu, M.; Pan, X.-Y.; Zhang, Z.-J.; van Kleunen, M.; Li, B. Testing the Shifting Defense Hypothesis for Constitutive and Induced Resistance and Tolerance. J. Pest Sci. 2020, 93, 355–364. [Google Scholar] [CrossRef]
- Li, X.; Png, G.K.; Sun, S.; Shi, H.; Jin, K.; Li, Y. Positive Microbial Legacy and Short-Term Clonal Plasticity Aid Grazing Tolerance of a Widespread Grass Species. Plant Soil 2022, 473, 291–303. [Google Scholar] [CrossRef]
- Geng, Y.-P.; Pan, X.-Y.; Xu, C.-Y.; Zhang, W.-J.; Li, B.; Chen, J.-K.; Lu, B.-R.; Song, Z.-P. Phenotypic Plasticity Rather than Locally Adapted Ecotypes Allows the Invasive Alligator Weed to Colonize a Wide Range of Habitats. Biol. Invasions 2007, 9, 245–256. [Google Scholar] [CrossRef]
- Rodríguez, J.; Lorenzo, P.; González, L. Phenotypic Plasticity of Invasive Carpobrotus edulis Modulates Tolerance against Herbivores. Biol. Invasions 2021, 23, 1859–1875. [Google Scholar] [CrossRef]
- Campoy, J.G.; Sobral, M.; Carro, B.; Lema, M.; Barreiro, R.; Retuerto, R. Epigenetic and Phenotypic Responses to Experimental Climate Change of Native and Invasive Carpobrotus edulis. Front. Plant Sci. 2022, 13, 888391. [Google Scholar] [CrossRef]
- Karban, R. The Ecology and Evolution of Induced Resistance against Herbivores. Funct. Ecol. 2011, 25, 339–347. [Google Scholar] [CrossRef]
- Gao, L.; Geng, Y.; Li, B.; Chen, J.; Yang, J. Genome-Wide DNA Methylation Alterations of Alternanthera philoxeroides in Natural and Manipulated Habitats: Implications for Epigenetic Regulation of Rapid Responses to Environmental Fluctuation and Phenotypic Variation: Environmental Epigenetics of A. Philoxeroides. Plant Cell Environ. 2010, 33, 1820–1827. [Google Scholar] [CrossRef]
- Shi, W.; Chen, X.; Gao, L.; Xu, C.-Y.; Ou, X.; Bossdorf, O.; Yang, J.; Geng, Y. Transient Stability of Epigenetic Population Differentiation in a Clonal Invader. Front. Plant Sci. 2019, 9, 1851. [Google Scholar] [CrossRef] [Green Version]
- Schmid, R.; Holm, L.; Doll, J.; Holm, E.; Pancho, J.; Herberger, J. World Weeds: Natural Histories and Distribution. Taxon 1998, 47, 209. [Google Scholar] [CrossRef]
- Lu, X.-M.; Siemann, E.; He, M.-Y.; Wei, H.; Shao, X.; Ding, J.-Q. Climate Warming Increases Biological Control Agent Impact on a Non-Target Species. Ecol. Lett. 2015, 18, 48–56. [Google Scholar] [CrossRef] [Green Version]
- Ye, W.-H.; Li, J.; Cao, H.-L.; Ge, X.-J. Genetic Uniformity of Alternanthera philoxeroides in South China. Weed Res. 2003, 43, 297–302. [Google Scholar] [CrossRef]
- Wang, B.; Li, W.; Wang, J. Genetic Diversity of Alternanthera philoxeroides in China. Aquat. Bot. 2005, 81, 277–283. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, C.-J.; Ma, L.; Zhou, X.-M.; Sun, X.; Ding, J.-Q. Elevated Temperature Decreases Preferences of Native Herbivores to an Invasive Plant. Entomol. Gen. 2021, 41, 137–146. [Google Scholar] [CrossRef]
- Puy, J.; Dvořáková, H.; Carmona, C.P.; de Bello, F.; Hiiesalu, I.; Latzel, V. Improved Demethylation in Ecological Epigenetic Experiments: Testing a Simple and Harmless Foliar Demethylation Application. Methods Ecol. Evol. 2018, 9, 744–753. [Google Scholar] [CrossRef]
- Atighi, M.R.; Verstraeten, B.; De Meyer, T.; Kyndt, T. Genome-wide DNA Hypomethylation Shapes Nematode Pattern-triggered Immunity in Plants. New Phytol. 2020, 227, 545–558. [Google Scholar] [CrossRef]
- Ruocco, M.; Entrambasaguas, L.; Dattolo, E.; Milito, A.; Marín-Guirao, L.; Procaccini, G. A King and Vassals’ Tale: Molecular Signatures of Clonal Integration in Posidonia oceanica under Chronic Light Shortage. J. Ecol. 2021, 109, 294–312. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]






| Variables | G1 Herbivory (G1H) | G1 Root Order (GR) | G1H × GR | |||
|---|---|---|---|---|---|---|
| F | p | F | p | F | p | |
| Growth | ||||||
| Total mass | 1.13 | 0.303 | 0.35 | 0.565 | 5.53 | 0.032 |
| Leaf mass | 1.74 | 0.206 | 0.37 | 0.553 | 5.65 | 0.030 |
| Stem mass | 1.17 | 0.295 | 0.13 | 0.727 | 4.86 | 0.043 |
| Root mass | 0.50 | 0.488 | 1.37 | 0.259 | 1.91 | 0.185 |
| Number of nodes | <0.01 | 0.963 | <0.01 | 0.978 | 3.23 | 0.091 |
| Number of leaves | 0.05 | 0.818 | 0.04 | 0.837 | 4.31 | 0.054 |
| Solon length | 2.28 | 0.151 | 0.10 | 0.761 | 4.77 | 0.044 |
| Biomass allocation | ||||||
| Root-to-shoot ratio | 5.99 | 0.026 | 0.01 | 0.905 | 1.25 | 0.280 |
| Physiology | ||||||
| Total phenolic | 7.50 | 0.018 | 0.29 | 0.600 | 0.54 | 0.477 |
| Water-soluble sugar | 2.75 | 0.123 | 0.01 | 0.939 | 0.01 | 0.936 |
| Starch | 0.19 | 0.674 | 5.13 | 0.043 | 0.95 | 0.349 |
| Total NSC | 0.95 | 0.348 | 2.68 | 0.127 | 0.52 | 0.486 |
| DNA methylation | 1.02 | 0.332 | 0.05 | 0.826 | 0.30 | 0.597 |
| Variable | G1 Herbivory (G1H) | G1 Root Order (GR) | G3 Herbivory (G3H) | G1H × GR | G1H × G3H | GR × G3H | G1H × GR × G3H | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | F | p | F | p | ||
| Growth | |||||||||||||||
| Total mass | 3.18 | 0.081 | 0.26 | 0.614 | 25.94 | <0.001 | 2.96 | 0.092 | 2.78 | 0.102 | 0.79 | 0.380 | 0.93 | 0.339 | |
| Leaf mass | 3.12 | 0.084 | 0.51 | 0.479 | 49.52 | <0.001 | 1.85 | 0.180 | 2.08 | 0.156 | 0.95 | 0.334 | 0.46 | 0.502 | |
| Stem mass | 1.52 | 0.223 | 0.19 | 0.665 | 26.46 | <0.001 | 1.99 | 0.165 | 3.50 | 0.067 | 1.53 | 0.222 | 0.55 | 0.462 | |
| Root mass | 5.10 | 0.029 | 0.26 | 0.610 | 1.40 | 0.243 | 4.30 | 0.044 | 2.14 | 0.150 | 0.05 | 0.816 | 2.20 | 0.144 | |
| Number of nodes | 0.80 | 0.376 | 0.08 | 0.776 | 41.13 | <0.001 | 1.22 | 0.275 | 1.85 | 0.180 | 0.22 | 0.645 | 0.20 | 0.655 | |
| Number of leaves | 1.96 | 0.168 | 0.02 | 0.896 | 38.56 | <0.001 | 1.72 | 0.195 | 2.24 | 0.141 | 0.93 | 0.340 | 0.26 | 0.610 | |
| Stolon length | 1.16 | 0.288 | 0.33 | 0.567 | 27.90 | <0.001 | 0.75 | 0.392 | 3.27 | 0.077 | 0.14 | 0.713 | 0.59 | 0.447 | |
| Biomass allocation | |||||||||||||||
| Root-to-shoot ratio | 1.37 | 0.248 | 0.05 | 0.833 | 28.50 | <0.001 | 0.10 | 0.756 | 1.85 | 0.180 | 0.28 | 0.602 | 0.15 | 0.705 | |
| Physiology | |||||||||||||||
| Total phenolic | 0.27 | 0.606 | 2.78 | 0.105 | 0.11 | 0.737 | 0.01 | 0.911 | 0.79 | 0.381 | 0.07 | 0.791 | <0.01 | 0.996 | |
| water-soluble sugar | 1.79 | 0.191 | 2.13 | 0.154 | 0.12 | 0.736 | 0.04 | 0.850 | 0.01 | 0.908 | 0.35 | 0.556 | 0.13 | 0.722 | |
| Starch | 0.15 | 0.706 | 0.15 | 0.698 | 16.29 | <0.001 | <0.01 | 0.985 | 1.55 | 0.222 | 1.23 | 0.276 | 0.82 | 0.371 | |
| Total NSC | 0.47 | 0.498 | 0.57 | 0.457 | 15.26 | <0.001 | 0.02 | 0.884 | 1.48 | 0.233 | 0.85 | 0.363 | 0.40 | 0.533 | |
| DNA methylation | 1.54 | 0.233 | 0.05 | 0.831 | / | / | 0.06 | 0.813 | / | / | / | / | / | / | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fu, Q.-Y.; Yu, C.-L.; Dong, R.; Shi, J.; Luo, F.-L.; Gao, J.-Q.; Li, H.-L.; Dong, B.-C.; Yu, F.-H. Transgenerational Herbivory Effects on Performance of Clonal Offspring of the Invasive Plant Alternanthera philoxeroides. Plants 2023, 12, 1180. https://doi.org/10.3390/plants12051180
Fu Q-Y, Yu C-L, Dong R, Shi J, Luo F-L, Gao J-Q, Li H-L, Dong B-C, Yu F-H. Transgenerational Herbivory Effects on Performance of Clonal Offspring of the Invasive Plant Alternanthera philoxeroides. Plants. 2023; 12(5):1180. https://doi.org/10.3390/plants12051180
Chicago/Turabian StyleFu, Qiu-Yue, Cheng-Ling Yu, Ran Dong, Juan Shi, Fang-Li Luo, Jun-Qin Gao, Hong-Li Li, Bi-Cheng Dong, and Fei-Hai Yu. 2023. "Transgenerational Herbivory Effects on Performance of Clonal Offspring of the Invasive Plant Alternanthera philoxeroides" Plants 12, no. 5: 1180. https://doi.org/10.3390/plants12051180