
Impact of the Alien Aquatic Plant Ludwigia hexapetala on the Native Utricularia australis: Evidence from an Indoor Experiment

Abstract
:1. Introduction
2. Results and Discussion
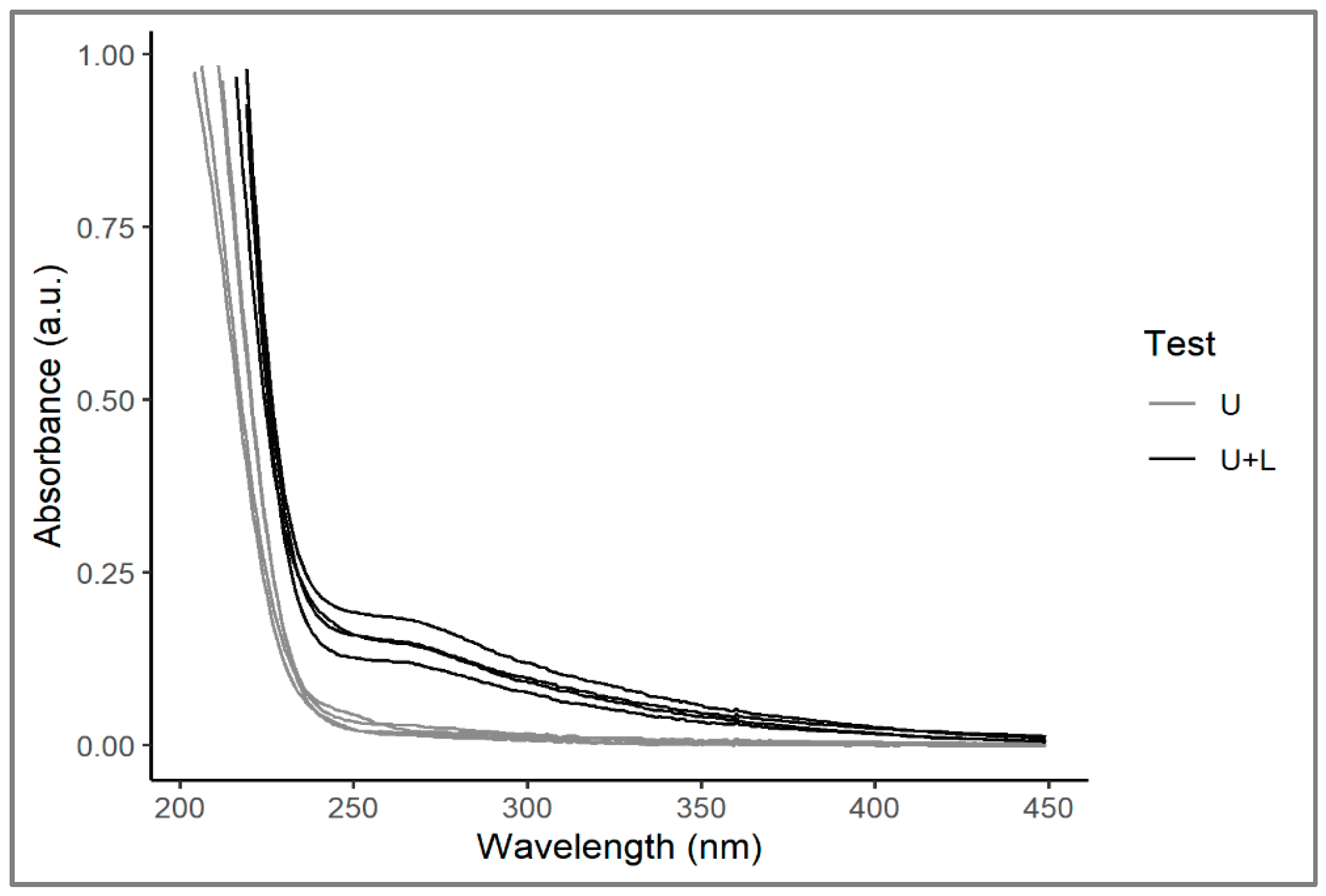
2.1. Water Chemical and Physical Variations
2.2. Plant Parameter Analyses
3. Materials and Methods
3.1. Plant Material
3.2. Experimental Design
3.3. Water Chemical and Physical Analyses
3.4. Plant Parameter Measures
3.5. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seebens, H.; Blackburn, T.M.; Dyer, E.E.; Genovesi, P.; Hulme, P.E.; Jeschke, J.M.; Pagad, S.; Pysek, P.; Winter, M.; Arianoutsou, M.; et al. No saturation in the accumulation of alien species worldwide. Nat. Commun. 2017, 8, 14435. [Google Scholar] [CrossRef]
- Pysek, P.; Jarosík, V.; Hulme, P.E.; Pergl, J.; Hejda, M.; Schaffner, U.; Vilà, M. A global assessment of invasive plant impacts on resident species, communities and ecosystems: The interaction of impact measures, invading species’ traits and environment. Glob. Chang. Biol. 2012, 18, 1725–1737. [Google Scholar] [CrossRef]
- Gigante, D.; Acosta, A.T.R.; Agrillo, E.; Armiraglio, S.; Assini, S.; Attorre, F.; Bagella, S.; Buffa, G.; Casella, L.; Giancola, C.; et al. Habitat conservation in Italy: The state of the art in the light of the first European red list of terrestrial and freshwater habitats. Rend. Lincei Sci. Fis. Nat. 2018, 29, 251–265. [Google Scholar] [CrossRef]
- Viciani, D.; Vidali, M.; Gigante, D.; Bolpagni, R.; Villani, M.; Acosta, A.T.R.; Adorni, M.; Aleffi, M.; Allegrezza, M.; Angiolini, C.; et al. A first checklist of the alien-dominated vegetation in Italy. Plant Sociol. 2020, 57, 29–54. [Google Scholar] [CrossRef]
- Dobson, A.; Lodge, D.; Alder, J.; Cumming, G.S.; Keymer, J.; McGlade, J.; Mooney, H.; Rusak, J.A.; Sala, O.; Wolters, V.; et al. Habitat loss, trophic collapse, and the decline of ecosystem services. Ecology 2006, 87, 1915–1924. [Google Scholar] [CrossRef]
- Maes, J. A Model for the Assessment of Habitat Conservation Status in the EU; European Commission Joint Research Centre, Institute for Environment and Sustainability: Luxembourg, 2013.
- Lazzaro, L.; Bolpagni, R.; Buffa, G.; Gentili, R.; Lonati, M.; Stinca, A.; Acosta Rosario, A.T.; Adorni, M.; Aleffi, M.; Allegrezza, M.; et al. Impact of invasive alien plants on native plant communities and Natura 2000 habitats: State of the art, gap analysis and perspectives in Italy. J. Environ. Manag. 2020, 274, 111–140. [Google Scholar] [CrossRef] [PubMed]
- Shea, K.; Chesson, P. Community ecology theory as a framework for biological invasions. Trends Ecol. Evol. 2002, 17, 170–176. [Google Scholar] [CrossRef]
- Anufriieva, E.V.; Shadrin, N.V. Extreme hydrological events destabilize aquatic ecosystems and open doors for alien species. Quarter. Internat. 2018, 475, 11–15. [Google Scholar] [CrossRef]
- Union List. List of Invasive Alien Species of Union Concern. 2016. Available online: https://ec.europa.eu/environment/nature/invasivealien/list/index_en.htm (accessed on 16 December 2022).
- Ruaux, B.; Greulich, S.; Haury, J.; Berton, J.P. Sexual reproduction of two alien invasive Ludwigia (Onagraceae) on the middle Loire River, France. Aquat. Bot. 2009, 90, 143–148. [Google Scholar] [CrossRef]
- Gori, B.; Pezzi, G.; Brundu, G.; Ceschin, S.; Pelella, E.; Alessandrini, A.; Amadei, L.; Andreatta, S.; Ardenghi, N.M.G.; Armiraglio, S.; et al. Invasion trends of aquatic Ludwigia hexapetala and L. peploides subsp. montevidensis (Onagraceae) in Italy based on herbarium records and global datasets. Neobiota 2023. submitted. [Google Scholar]
- Armitage, J.D.; Konyves, K.; Bailey, J.P.; David, J.C.; Culham, A. A molecular, morphological and cytological investigation of the identity of non-native Ludwigia (Onagraceae) populations in Britain. New J. Bot. 2013, 3, 88–95. [Google Scholar] [CrossRef]
- Martins, C. Sur les Racines Aérifères ou Vessies Natatoires des Espèces Aquatiques du Genre Jussiaea L. Mémories Sect. Sc; Académie des Sciences et Lettres de Montpellier: Montpellier, France, 1866; Volume 6, pp. 353–370. [Google Scholar]
- Dandelot, S.; Robles, C.; Pech, N.; Cazaubon, A.; Verlaque, R. Allelopathic potential of two invasive alien Ludwigia spp. Aquat. Bot. 2008, 88, 311–316. [Google Scholar] [CrossRef]
- Di Pietro, R.; Misano, G.; Wagensommer, R.P. Notulae alla checklist della flora vascolare Italiana 4. Inf. Bot. Ital. 2007, 39, 401–435. [Google Scholar]
- Bonali, F.; Giordana, F.; Pellizzari, M.; Merloni, N.; Lazzari, G.; Saiani, D.; Piccoli, F. Notulae alla checklist della flora vascolare italiana 2. Inf. Bot. Ital. 2006, 38, 191–214. [Google Scholar]
- Galasso, G.; Domina, G.; Andreatta, S.; Angiolini, C.; Ardenghi, N.M.G.; Aristarchi, C.; Arnoul, M.; Azzella, M.M.; Bacchetta, G.; Bartolucci, F.; et al. Notulae to the Italian alien vascular flora: 8. Ital. Bot. 2019, 8, 63–93. [Google Scholar] [CrossRef]
- Thiebaut, G.; Thouvenot, L.; Rodriguez-Perez, H. Allelopathic effect of the invasive Ludwigia hexapetala on growth of three macrophyte species. Front. Plant Sci. 2018, 9, 1835. [Google Scholar] [CrossRef] [PubMed]
- Dandelot, S.; Verlaque, R.; Dutartre, A.; Cazaubon, A. Ecological, dynamic and taxonomic problems due to Ludwigia (Onagraceae) in France. Hydrobiologia 2005, 551, 131–136. [Google Scholar] [CrossRef]
- Stiers, I.; Crohain, N.; Josens, G.; Triest, L. Impact of three aquatic invasive species on native plants and macroinvertebrates in temperate ponds. Biol. Invasions 2011, 13, 2715–2726. [Google Scholar] [CrossRef]
- Thouvenot, L.; Haury, J.; Thiebaut, G. A success story: Water primroses, aquatic plant pests. Aquat. Conserv. Mar. Freshw. Ecosyst. 2013, 23, 790–803. [Google Scholar] [CrossRef]
- Wardle, D.A.; Karban, R.; Callaway, R.M. The ecosystem and evolutionary contexts of allelopathy. Trends Ecol. Evol. 2011, 26, 655–662. [Google Scholar]
- Bais, H.P.; Vepachedu, R.; Gilroy, S.; Callaway, R.M.; Vivanco, J.M. Allelopathy and exotic plant invasion: From molecules and genes to species interactions. Science 2003, 301, 1377–1380. [Google Scholar] [CrossRef] [PubMed]
- Rice, E.L. Allelopathy, 2nd ed.; Academic Press: New York, NY, USA, 1984. [Google Scholar]
- Rossi, G.; Orsenigo, S.; Gargano, D.; Montagnani, C.; Peruzzi, L.; Fenu, G.; Abeli, T.; Alessandrini, A.; Astuti, G.; Bacchetta, G.; et al. Lista Rossa della Flora Italiana. 2 Endemiti e Altre Specie Minacciate; Ministero dell’Ambiente e della Tutela del Territorio e del Mare: Rome, Italy, 2020.
- Acta Plantarum. Utricularia australis R. Br.—Scheda IPFI, Acta Plantarum. 2007. Available online: https://www.actaplantarum.org/flora/flora_info.php?id=8058 (accessed on 16 December 2022).
- Ceschin, S.; Bellini, A.; Traversetti, L.; Zuccarello, V.; Ellwood, N.T.W. Ecological study of the aquatic carnivorous plant Utricularia australis R. Br. (Lentibulariaceae). Aq. Ecol. 2020, 54, 295–307. [Google Scholar] [CrossRef]
- Ceschin, S.; Pelella, E.; Azzella, M.; Bellini, A.; Ellwood, N.T.W. Unusual underwater flowering of Utricularia australis populations: A botanical enigma? Aquat. Bot. 2022, 178, 103487. [Google Scholar] [CrossRef]
- Hoch, P.C.; Wagner, W.L.; Raven, P.H. The correct name for a section of Ludwigia, L. (Onagraceae). PhytoKeys 2015, 50, 31–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunt, R. Basic Growth Analysis, 1st ed.; Unwin Hyman: London, UK, 1990. [Google Scholar]
- Adamec, L.; Ondok, J.P. Water alkalization due to photosynthesis of aquatic plants: The dependence on total alkalinity. Aquat. Bot. 1992, 43, 93–98. [Google Scholar] [CrossRef]
- Sand-Jensen, K.; Riis, T.; Martinsen, K.T. Photosynthesis, growth, and distribution of plants in lowland streams—A synthesis and new data analyses of 40 years research. Freshw. Biol. 2022, 67, 1255–1271. [Google Scholar] [CrossRef]
- Wang, W.; Li, S.L.; Zhong, J.; Slowinski, S.; Li, S.; Li, C.; Yi, Y.; Dong, K.; Xu, S.; van Cappellen, P.; et al. Carbonate mineral dissolution and photosynthesis-induced precipitation regulate inorganic carbon cycling along the karst river-reservoir continuum, SW China. J. Hydrol. 2022, 615, 128621. [Google Scholar] [CrossRef]
- Denny, P. Sites of Nutrient Absorption in Aquatic Macrophytes. J. Ecol. 1972, 60, 819–829. [Google Scholar] [CrossRef]
- Liu, Z.H.; Macpherson, G.L.; Groves, C.; Martin, J.B.; Yuan, D.; Zeng, S. Large and active CO2 uptake by coupled carbonate weathering. Earth-Sci. Rev. 2018, 182, 42–49. [Google Scholar] [CrossRef]
- Kleeberg, A. Impact of aquatic macrophyte decomposition on sedimentary nutrient and metal mobilization in the initial stages of ecosystem development. Aquat. Bot. 2013, 105, 41–49. [Google Scholar] [CrossRef]
- Averett, J.E.; Zardini, E.M.; Hoch, P.C. Flavonoid systematics of ten sections of Ludwigia (Onagraceae). Biochem. Syst. Ecol. 1990, 18, 529–532. [Google Scholar] [CrossRef]
- Bazira Makenga, R.; Leroux, G.D.; Simard, R.R. Effect of benzoic acid, cinnamic acids on membrane permeability of soybean roots. J. Chem. Ecol. 1995, 21, 1271–1285. [Google Scholar] [CrossRef] [PubMed]
- Dragoeva, A.P.; Koleva, V.P.; Stoyanovea, Z.D. Allelopathic effects of Adonis vernalis L.: Root growth inhibition and cytogenetic alterations. J. Agric. Chem. Environ. 2015, 4, 48–55. [Google Scholar] [CrossRef]
- George, D.; Sindhu, P.V.; Menon, M.V.; Beena, C.; Reddy, T.V. Allelopathic effect of tulsi (Ocimum tenuiflorum L.) on germination and seedling growth of rice, cowpea and upland weeds. J. Trop. Agric. 2021, 59, 257–265. [Google Scholar]
- Madadi, E.; Fallah, S.; Sadeghpour, A.; Barani-Beiranvand, H. Exploring the use of chamomile (Matricaria chamomilla L.) bioactive compounds to control flixweed (Descurainia sophia L.) in bread wheat (Triticum aestivum L.): Implication for reducing chemical herbicide pollution. Saudi J. Biol. Sci. 2022, 29, 103421. [Google Scholar] [CrossRef]
- Osman, M.F.; Mutwali, E.M. Allelopathic effects of Amaranthus viridis L. on some growth characters of Swiss chard plant (Beta vulgaris var. Cicla). World J. Biol. Pharm. Health Sci. 2022, 9, 1–4. [Google Scholar] [CrossRef]
- Romani, A.; Zuccaccia, C.; Clementi, C. An NMR and UV-visible spectroscopic study of the principal colored component of Stil de Grain Lake. Dyes Pigm. 2006, 71, 218–223. [Google Scholar] [CrossRef]
- Thiébaut, G.; Rodriguez-Perez, H.; Jambon, O. Reciprocal interactions between the native Mentha aquatica and the invasive Ludwigia hexapetala in an outdoor experiment. Aquat Bot. 2019, 157, 17–23. [Google Scholar] [CrossRef]
- Billet, K.; Genitoni, J.; Bozec, M.; Renault, D.; Barloy, D. Aquatic and terrestrial morphotypes of the aquatic invasive plant, Ludwigia grandiflora, show distinct morphological and metabolomic responses. Ecol. Evol. 2018, 8, 2568–2579. [Google Scholar] [CrossRef]
- Yang, H.; Yang, J.; Lv, Y.; He, J. SPAD values and nitrogen nutrition index for the evaluation of rice nitrogen status. Plant Prod. Sci. 2015, 17, 81–92. [Google Scholar] [CrossRef]
- Shu, M.; Zuo, J.; Shen, M.; Yin, P.; Wang, M.; Yang, X.; Tang, J.; Li, B.; Ma, Y. Improving the estimation accuracy of SPAD values for maize leaves by removing UAV hyperspectral image backgrounds. Int. J. Remote Sens. 2021, 42, 5862–5881. [Google Scholar] [CrossRef]
- Cao, Y.; Xu, H.; Song, J.; Yang, Y.; Hu, X.; Wiyao, K.T.; Zhai, Z. Applying spectral fractal dimension index to predict the SPAD value of rice leaves under bacterial blight disease stress. Plant Meth. 2022, 18, 67. [Google Scholar] [CrossRef]
- Lucchese, F. Atlante della flora vascolare del Lazio: Cartografia, ecologia e biogeografia. Parte Gen. Flora Alloctona 2017, 1, 279. [Google Scholar]
- Adamec, L. Rootless aquatic plant Aldrovanda vesiculosa: Physiological polarity, mineral nutrition, and importance of carnivory. Biol. Plant. 2000, 43, 113–119. [Google Scholar] [CrossRef]
- Adamec, L.; Kovářová, M. Field growth characteristics of two aquatic carnivorous plants, Aldrovanda vesiculosa and Utricularia australis. Folia Geobot. 2006, 41, 395–406. [Google Scholar] [CrossRef]
- Radford, P.J. Growth analysis formulae—their use and abuse. Crop. Sci. 1967, 7, 171–175. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Sofw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Barton, K. MuMIn: Multi-Model Inference, R Package Version 1.46.0; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 2 December 2022).
- Wickham, H. Data analysis. In ggplot2: Elegant Graphics for Data Analysis; Springer: Cham, Switzerland, 2016; pp. 189–201. [Google Scholar]
- Pysek, P.; Pysek, A. Invasion by Heracleum mantegazzianum in different habitats in the Czech Republic. J. Veg. Sci. 1995, 6, 711–718. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Marginal R2 | Conditional R2 | Test p Value | Time p Value | Interaction p Value |
|---|---|---|---|---|---|
| Dissolved oxygen | 0.9012 | 0.9648 | 0.0006 * | 5.8 × 10−12 * | 3.3 × 10−7 * |
| Conductivity | 0.9718 | 0.9897 | 2.9 × 10−5 * | 2.2 × 10−16 * | 5.9 × 10−13 * |
| pH | 0.9800 | 0.9804 | 7.6 × 10−6 * | 2.2 × 10−16 * | 3.1 × 10−5 * |
| Utricularia total shoot length | 0.3213 | 0.7991 | 0.3647 | 0.2749 | 0.0003 * |
| Utricularia FW | 0.7906 | 0.8750 | 0.0033 * | 8.5 × 10−5 * | 0.0119 * |
| Utricularia RGR | 0.5035 | 0.5035 | 0.0011 * | 0.3111 | N.S. |
| Utricularia internode number | 0.2605 | 0.8623 | 0.7187 | 0.0001 * | 0.0039 * |
| Utricularia internode length | 0.7890 | 0.7890 | 1.2 × 10−5 * | 1.5 × 10−7 * | 0.2605 |
| Utricularia trap number | 0.4271 | 0.4793 | 0.0283 * | 0.0124 * | N.S. |
| Ludwigia shoot length | 0.5887 | 0.7007 | NA | 0.0033 * | NA |
| Ludwigia FW | 0.5842 | 0.9697 | NA | 3.7 × 10−7 * | NA |
| Ludwigia RGR | 0.0555 | 0.0555 | NA | 0.7317 | NA |
| Ludwigia SPAD | 0.5417 | 0.6707 | NA | 0.0060 * | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pelella, E.; Questino, B.; Ceschin, S. Impact of the Alien Aquatic Plant Ludwigia hexapetala on the Native Utricularia australis: Evidence from an Indoor Experiment. Plants 2023, 12, 811. https://doi.org/10.3390/plants12040811
Pelella E, Questino B, Ceschin S. Impact of the Alien Aquatic Plant Ludwigia hexapetala on the Native Utricularia australis: Evidence from an Indoor Experiment. Plants. 2023; 12(4):811. https://doi.org/10.3390/plants12040811
Chicago/Turabian StylePelella, Emanuele, Beatrice Questino, and Simona Ceschin. 2023. "Impact of the Alien Aquatic Plant Ludwigia hexapetala on the Native Utricularia australis: Evidence from an Indoor Experiment" Plants 12, no. 4: 811. https://doi.org/10.3390/plants12040811