
Antiphotoaging and Skin-Protective Activities of Ardisia silvestris Ethanol Extract in Human Keratinocytes

 , , , and
, , , and 
Abstract
:1. Introduction
2. Results
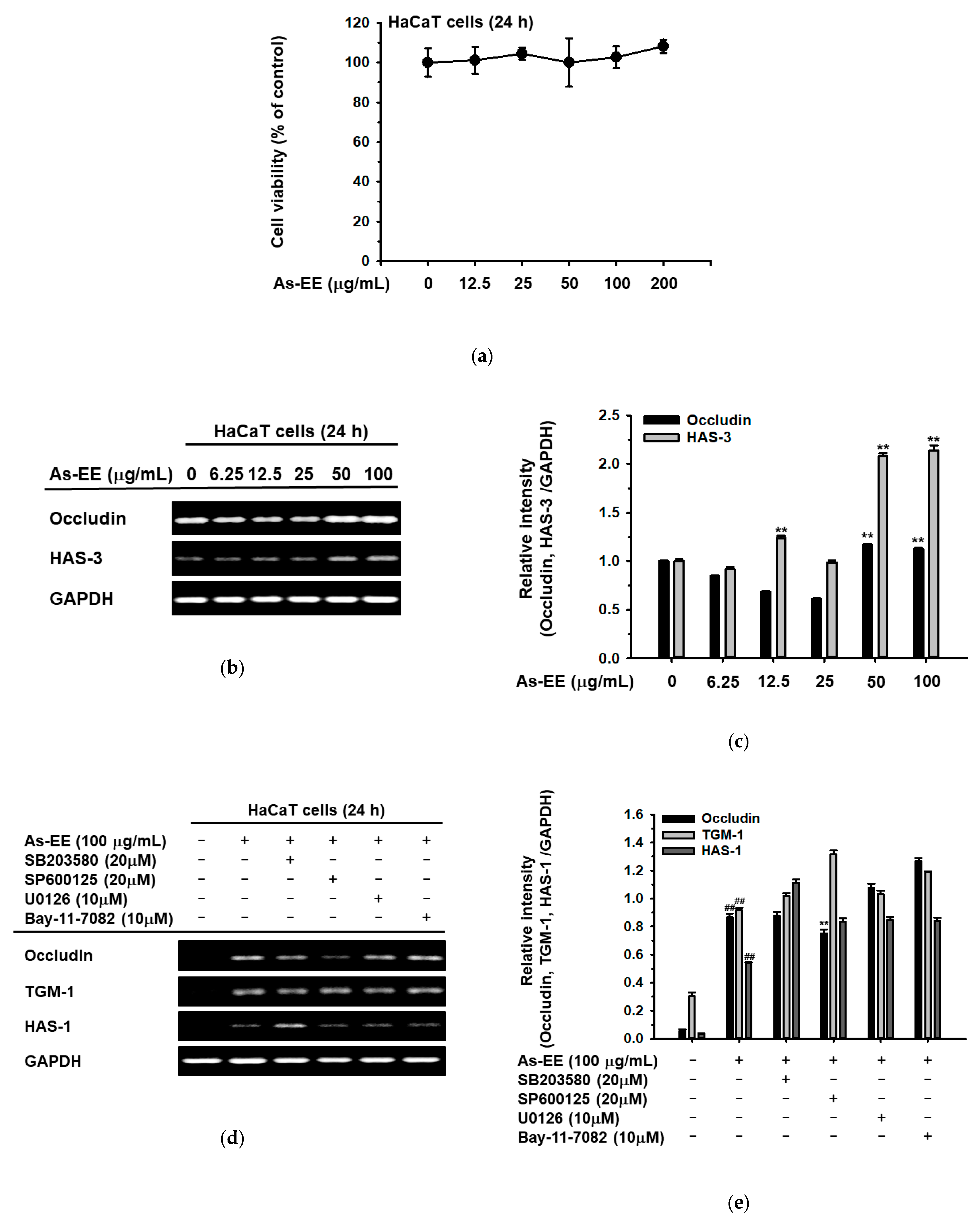
2.1. Effects of As-EE on Antioxidative Capacity
2.2. Effects of As-EE on Cell Viability and Skin Moisture Protection Activity
2.3. Effects of As-EE on MAPK-Mediated AP-1 and CREB Signaling Pathway
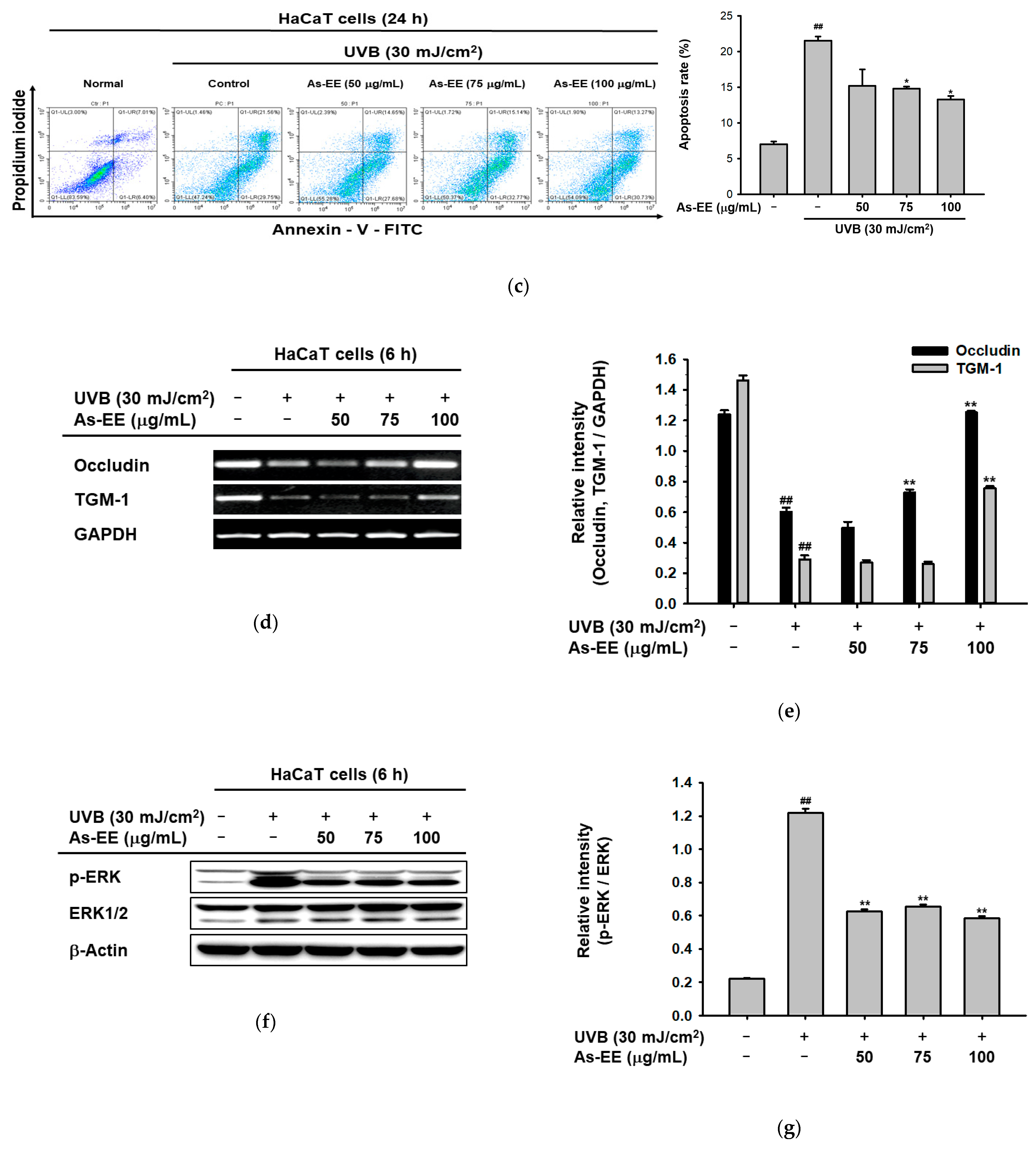
2.4. Moisturizing and Anti-Apoptotic Effect of As-EE in UVB-Irradiated Human Keratinocytes
3. Discussion
4. Materials and Methods
4.1. Materials
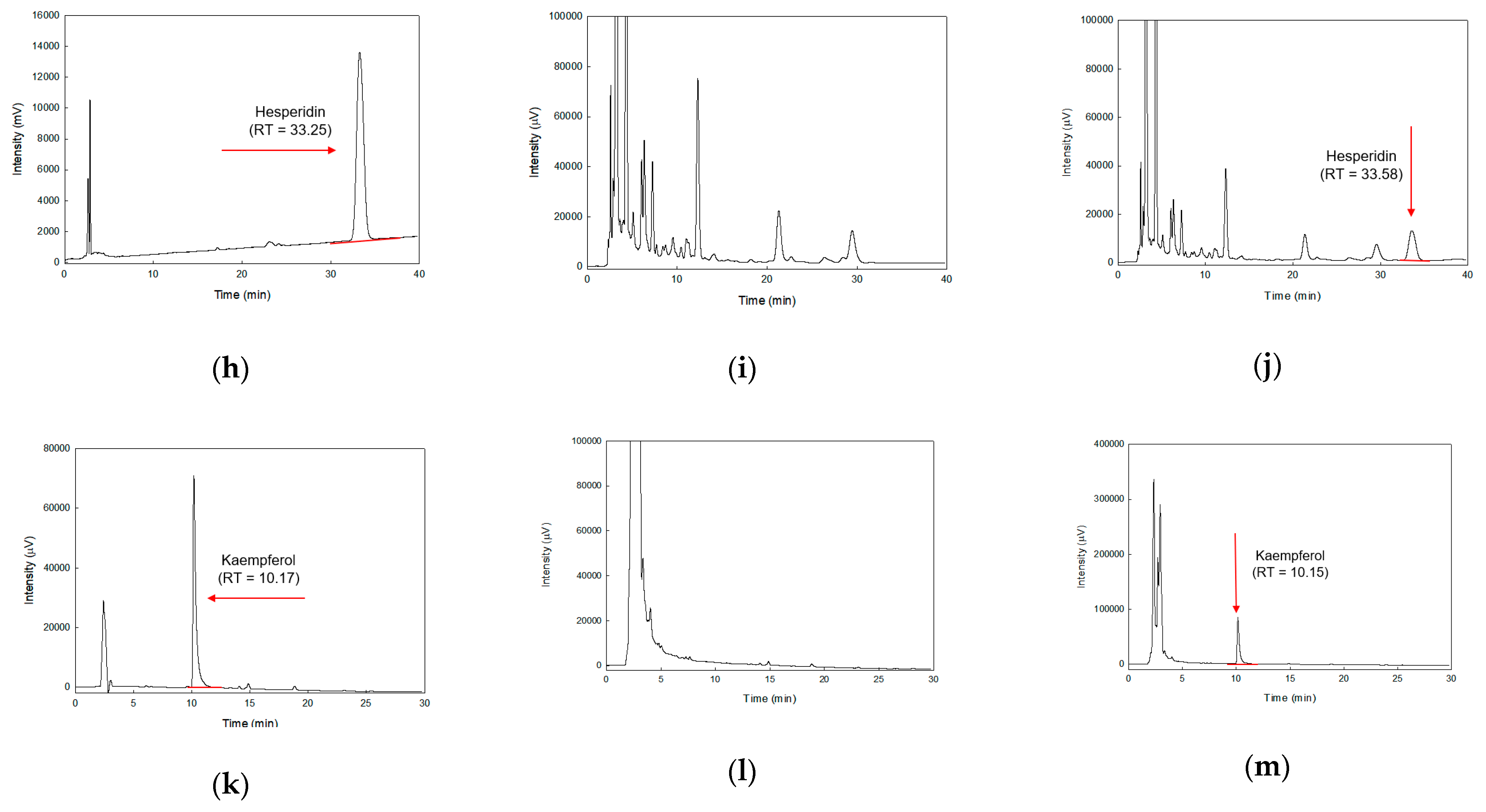
4.2. Prepartion of As-EE and HPLC Analysis
4.3. Determination of Total Phenolic Content
4.4. DPPH Assay
4.5. ABTS Assay
4.6. Cupric Ion Reducing Antioxidant Capacity (CUPRAC) Assay
4.7. Ferric-Reducing Antioxidant Power (FRAP) Assay
4.8. Cell Culture
4.9. Cell Viability Test
4.10. mRNA Preparation and Reverse Transcription Polymerase Chain Reaction (PCR)
4.11. Reporter Gene Assays
4.12. Total Cell Lysate Preparation
4.13. Immunoblotting Analysis
4.14. UVB Irradiation
4.15. Morphological Changes
4.16. PI and Annexin V Staining (FACS)
4.17. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| As-EE | Ardisia silvestris ethanol extract |
| UVB | Ultraviolet B |
| ROS | reactive oxygen species |
| GC-MS | Gas chromatography–mass spectrometry |
| MTT | 3-4-5-dimethylthiazol-2-yl)-2-5-diphenyltetrazolium bromide |
| CUPRAC | Cupric Ion Reducing Antioxidant Capacity |
| FRAP | Ferric-Reducing Antioxidant Power |
| TPTZ | 2,4,6-tri(2-pyridyl)-s-triazine |
| HAS | Hyaluronic acid synthase |
| TGM-1 | Transglutaminase-1 |
| FITC | fluorescein isothiocyanate |
| PI | propidium iodide |
References
- You, L.; Cho, J.Y. The Regulatory Role of Korean Ginseng in Skin Cells. J. Ginseng Res. 2021, 45, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Chambers, E.S.; Vukmanovic-Stejic, M. Skin Barrier Immunity and Ageing. Immunology 2020, 160, 116–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Than, U.T.T.; Leavesley, D.I.; Parker, T.J. Characteristics and Roles of Extracellular Vesicles Released by Epidermal Keratinocytes. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 2264–2272. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, A.V.; Harding, C.R. Moisturization and Skin Barrier Function. Dermatol. Ther. 2004, 17, 43–48. [Google Scholar] [CrossRef]
- Baroni, A.; Buommino, E.; De Gregorio, V.; Ruocco, E.; Ruocco, V.; Wolf, R. Structure and Function of the Epidermis Related to Barrier Properties. Clin. Dermatol. 2012, 30, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Tomita, Y.; Akiyama, M.; Shimizu, H. Stratum Corneum Hydration and Flexibility Are Useful Parameters to Indicate Clinical Severity of Congenital Ichthyosis. Exp. Dermatol. 2005, 14, 619–624. [Google Scholar] [CrossRef]
- Batt, M.D.; Fairhurst, E. Hydration of the Stratum Corneum. Int. J. Cosmet. Sci. 1986, 8, 253–264. [Google Scholar] [CrossRef]
- Ratz-Lyko, A.; Arct, J.; Pytkowska, K. Moisturizing and Antiinflammatory Properties of Cosmetic Formulations Containing Centella asiatica Extract. Indian J. Pharm. Sci. 2016, 78, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Rabe, J.H.; Mamelak, A.J.; McElgunn, P.J.S.; Morison, W.L.; Sauder, D.N. Photoaging: Mechanisms and Repair. J. Am. Acad. Dermatol. 2006, 55, 1–19. [Google Scholar] [CrossRef]
- Gu, Y.; Han, J.; Jiang, C.; Zhang, Y. Biomarkers, Oxidative Stress and Autophagy in Skin Aging. Ageing Res. Rev. 2020, 59, 101036. [Google Scholar] [CrossRef]
- D’Orazio, J.; Jarrett, S.; Amaro-Ortiz, A.; Scott, T. UV Radiation and the Skin. Int. J. Mol. Sci. 2013, 14, 12222–12248. [Google Scholar] [CrossRef] [Green Version]
- Coppo, R.; Orso, F.; Virga, F.; Dalmasso, A.; Baruffaldi, D.; Nie, L.; Clapero, F.; Dettori, D.; Quirico, L.; Grassi, E.; et al. ESDN Inhibits Melanoma Progression by Blocking E-Selectin Expression in Endothelial Cells via STAT3. Cancer Lett. 2021, 510, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Singhal, S.S.; Srivastava, S.; Mirzapoiazova, T.; Horne, D.; Awasthi, S.; Salgia, R. Targeting the Mercapturic Acid Pathway for the Treatment of Melanoma. Cancer Lett. 2021, 518, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Hu, Z.; Wang, X.; Li, L.; Zhu, B.; Lin, X.; Zhang, J.; Hua, Z. ANXA10 Promotes Melanoma Metastasis by Suppressing E3 Ligase TRIM41-Directed PKD1 Degradation. Cancer Lett. 2021, 519, 237–249. [Google Scholar] [CrossRef]
- Schuch, A.P.; Moreno, N.C.; Schuch, N.J.; Martins, C.F.; Menck, C.C.M.G.; Schuch, J.; Menck, C.F.M.; Machado, C.C. Sunlight damage to cellular DNA: Focus on oxidatively generated lesions. Free Radic. Biol. Med. 2017, 107, 110–124. [Google Scholar] [CrossRef]
- Jenkins, G.I. Signal Transduction in Responses to UV-B Radiation. Annu. Rev. Plant Biol. 2009, 60, 407–431. [Google Scholar] [CrossRef]
- Ma, Q.; Xu, Q.; Zhao, J.; Zhang, W.; Wang, Q.; Fang, J.; Lu, Z.; Liu, J.; Ma, L. Coupling HDAC4 with Transcriptional Factor MEF2D Abrogates SPRY4-Mediated Suppression of ERK Activation and Elicits Hepatocellular Carcinoma Drug Resistance. Cancer Lett. 2021, 520, 243–254. [Google Scholar] [CrossRef]
- Nyati, K.K.; Hashimoto, S.; Singh, S.K.; Tekguc, M.; Metwally, H.; Liu, Y.C.; Okuzaki, D.; Gemechu, Y.; Kang, S.; Kishimoto, T. The Novel Long Noncoding RNA AU021063, Induced by IL-6/Arid5a Signaling, Exacerbates Breast Cancer Invasion and Metastasis by Stabilizing Trib3 and Activating the Mek/Erk Pathway. Cancer Lett. 2021, 520, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Clark, O.; Daga, S.; Stoker, A.W. Tyrosine Phosphatase Inhibitors Combined with Retinoic Acid Can Enhance Differentiation of Neuroblastoma Cells and Trigger ERK- and AKT-Dependent, P53-Independent Senescence. Cancer Lett. 2013, 328, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Pearson, G.; Robinson, F.; Gibson, T.B.; Xu, B.E.; Karandikar, M.; Berman, K.; Cobb, M.H. Mitogen-Activated Protein (MAP) Kinase Pathways: Regulation and Physiological Functions. Endocr. Rev. 2001, 22, 153–183. [Google Scholar] [CrossRef]
- Gabasa, M.; Radisky, E.S.; Ikemori, R.; Bertolini, G.; Arshakyan, M.; Hockla, A.; Duch, P.; Rondinone, O.; Llorente, A.; Maqueda, M.; et al. MMP1 Drives Tumor Progression in Large Cell Carcinoma of the Lung through Fibroblast Senescence. Cancer Lett. 2021, 507, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Prelowska, M.K.; Mehlich, D.; Ugurlu, M.T.; Kedzierska, H.; Cwiek, A.; Kosnik, A.; Kaminska, K.; Marusiak, A.A.; Nowis, D. Inhibition of the ʟ-Glutamine Transporter ASCT2 Sensitizes Plasma Cell Myeloma Cells to Proteasome Inhibitors. Cancer Lett. 2021, 507, 13–25. [Google Scholar] [CrossRef]
- Zhao, D.; Qian, L.; Zhuang, D.; Wang, L.; Cao, Y.; Zhou, F.; Zhang, S.; Liu, Y.; Liang, Y.; Zhang, W.; et al. Inhibition of Ribosomal RNA Processing 15 Homolog (RRP15), Which Is Overexpressed in Hepatocellular Carcinoma, Suppresses Tumour Growth via Induction of Senescence and Apoptosis. Cancer Lett. 2021, 519, 315–327. [Google Scholar] [CrossRef]
- Sang, J.; Li, W.; Diao, H.J.; Fan, R.Z.; Huang, J.L.; Gan, L.; Zou, M.F.; Tang, G.H.; Yin, S. Jolkinolide B Targets Thioredoxin and Glutathione Systems to Induce ROS-Mediated Paraptosis and Apoptosis in Bladder Cancer Cells. Cancer Lett. 2021, 509, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Zou, Z.; Li, J.; Shen, Q.; Liu, L.; An, X.; Yang, S.; Xing, D. Photoactivation of Mitochondrial Reactive Oxygen Species-Mediated Src and Protein Kinase C Pathway Enhances MHC Class II-Restricted T Cell Immunity to Tumours. Cancer Lett. 2021, 523, 57–71. [Google Scholar] [CrossRef]
- Wang, S.F.; Chang, Y.L.; Tzeng, Y.D.; Wu, C.L.; Wang, Y.Z.; Tseng, L.M.; Chen, S.; Lee, H.C. Mitochondrial Stress Adaptation Promotes Resistance to Aromatase Inhibitor in Human Breast Cancer Cells via ROS/Calcium up-Regulated Amphiregulin–Estrogen Receptor Loop Signaling. Cancer Lett. 2021, 523, 82–99. [Google Scholar] [CrossRef] [PubMed]
- Warner, H.R. Apoptosis: A Two-Edged Sword in Aging. Anticancer Res. 1999, 19, 2837–2842. [Google Scholar] [CrossRef]
- Zeisel, S.H. Antioxidants Suppress Apoptosis. J. Nutr. 2004, 134, 3179S–3180S. [Google Scholar] [CrossRef] [Green Version]
- Lu, B.; Chen, H.-D.; Hong-Guang, H.-G. The Relationship between Apoptosis and Aging. Adv. Biosci. Biotechnol. 2012, 3, 705–711. [Google Scholar] [CrossRef] [Green Version]
- Higami, Y.; Shimokawa, I. Apoptosis in the Aging Process. Cell Tissue Res. 2000, 301, 125–132. [Google Scholar] [CrossRef]
- Shields, H.J.; Traa, A.; Van Raamsdonk, J.M. Beneficial and Detrimental Effects of Reactive Oxygen Species on Lifespan: A Comprehensive Review of Comparative and Experimental Studies. Front. Cell Dev. Biol. 2021, 9, 628157. [Google Scholar] [CrossRef]
- Oh, Y.; Jung, H.R.; Min, S.; Kang, J.; Jang, D.; Shin, S.; Kim, J.; Lee, S.E.; Sung, C.O.; Lee, W.S.; et al. Targeting Antioxidant Enzymes Enhances the Therapeutic Efficacy of the BCL-XL Inhibitor ABT-263 in KRAS-Mutant Colorectal Cancers. Cancer Lett. 2021, 497, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Gleneadie, H.J.; Baker, A.H.; Batis, N.; Bryant, J.; Jiang, Y.; Clokie, S.J.H.; Mehanna, H.; Garcia, P.; Gendoo, D.M.A.; Roberts, S.; et al. The Anti-Tumour Activity of DNA Methylation Inhibitor 5-Aza-2′-Deoxycytidine Is Enhanced by the Common Analgesic Paracetamol through Induction of Oxidative Stress. Cancer Lett. 2021, 501, 172–186. [Google Scholar] [CrossRef]
- Codenotti, S.; Marampon, F.; Triggiani, L.; Bonù, M.L.; Magrini, S.M.; Ceccaroli, P.; Guescini, M.; Gastaldello, S.; Tombolini, V.; Poliani, P.L.; et al. Caveolin-1 Promotes Radioresistance in Rhabdomyosarcoma through Increased Oxidative Stress Protection and DNA Repair. Cancer Lett. 2021, 505, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.B.; Wang, W.J.; Xu, C.; Xie, Y.J.; Wang, X.R.; Zhang, Y.Z.; Huang, J.M.; Huang, M.; Xie, C.; Liu, P.; et al. Luteolin and Its Derivative Apigenin Suppress the Inducible PD-L1 Expression to Improve Anti-Tumor Immunity in KRAS-Mutant Lung Cancer. Cancer Lett. 2021, 515, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Ge, C.; Wang, Y.; Feng, Y.; Wang, S.; Zhang, K.; Xu, X.; Zhang, Z.; Zhao, Y.; Wang, Y.; Gao, L.; et al. Suppression of Oxidative Phosphorylation and IDH2 Sensitizes Colorectal Cancer to a Naphthalimide Derivative and Mitoxantrone. Cancer Lett. 2021, 519, 30–45. [Google Scholar] [CrossRef] [PubMed]
- Nichols, J.A.; Katiyar, S.K. Skin Photoprotection by Natural Polyphenols: Anti-Inflammatory, Antioxidant and DNA Repair Mechanisms. Arch. Dermatol. Res. 2010, 302, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Kasote, D.M.; Katyare, S.S.; Hegde, M.V.; Bae, H. Significance of Antioxidant Potential of Plants and Its Relevance to Therapeutic Applications. Int. J. Biol. Sci. 2015, 11, 982–991. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, B.T.A.; Peloi, K.E.; Ximenes, V.F.; Nakamura, C.V.; de Oliveira Silva Lautenschlager, S. 2-Acetylphenothiazine Protects L929 Fibroblasts against UVB-Induced Oxidative Damage. J. Photochem. Photobiol. B Biol. 2021, 216, 112130. [Google Scholar] [CrossRef]
- Bayerl, C. Topical Hormonal Treatment in Anti-Aging of the Skin. Hautarzt 2020, 71, 786–790. [Google Scholar] [CrossRef]
- Jadoon, S.; Karim, S.; Bin Asad, M.H.H.; Akram, M.R.; Kalsoom Khan, A.; Malik, A.; Chen, C.; Murtaza, G. Anti-Aging Potential of Phytoextract Loaded-Pharmaceutical Creams for Human Skin Cell Longetivity. Oxid. Med. Cell. Longev. 2015, 2015, 709628. [Google Scholar] [CrossRef] [Green Version]
- Cao, C.; Xiao, Z.; Wu, Y.; Ge, C. Diet and Skin Aging—From the Perspective of Food Nutrition. Nutrients 2020, 12, 870. [Google Scholar] [CrossRef] [Green Version]
- Avadhani, K.S.; Manikkath, J.; Tiwari, M.; Chandrasekhar, M.; Godavarthi, A.; Vidya, S.M.; Hariharapura, R.C.; Kalthur, G.; Udupa, N.; Mutalik, S. Skin Delivery of Epigallocatechin-3-Gallate (EGCG) and Hyaluronic Acid Loaded Nano-Transfersomes for Antioxidant and Anti-Aging Effects in UV Radiation Induced Skin Damage. Drug Deliv. 2017, 24, 61–74. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Li, N.; Yan, Y.; Liu, Y.; Xiong, K.; Liu, Y.; Xia, Q.; Zhang, H.; Liu, Z. Recent Advances in the Anti-Aging Effects of Phytoestrogens on Collagen, Water Content, and Oxidative Stress. Phyther. Res. 2020, 34, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Xiao, F.; Xu, T.; Lu, B.; Liu, R. Guidelines for Antioxidant Assays for Food Components. Food Front. 2020, 1, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Munteanu, I.G.; Apetrei, C. Analytical Methods Used in Determining Antioxidant Activity: A Review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef] [PubMed]
- Shen, P.; Lin, W.; Deng, X.; Ba, X.; Han, L.; Chen, Z.; Qin, K.; Huang, Y.; Tu, S. Potential Implications of Quercetin in Autoimmune Diseases. Front. Immunol. 2021, 12, 1991. [Google Scholar] [CrossRef]
- Liu, S.; Adewole, D.; Yu, L.; Sid, V.; Wang, B.; Karmin, O.; Yang, C. Rutin Attenuates Inflammatory Responses Induced by Lipopolysaccharide in an in Vitro Mouse Muscle Cell (C2C12) Model. Poult. Sci. 2019, 98, 2756–2764. [Google Scholar] [CrossRef]
- Song, H.L.; Zhang, X.; Wang, W.Z.; Liu, R.H.; Zhao, K.; Liu, M.Y.; Gong, W.M.; Ning, B. Neuroprotective Mechanisms of Rutin for Spinal Cord Injury through Anti-Oxidation and Anti-Inflammation and Inhibition of P38 Mitogen Activated Protein Kinase Pathway. Neural Regen. Res. 2018, 13, 128–134. [Google Scholar] [CrossRef]
- Chen, C.Y.; Kao, C.L.; Liu, C.M. The Cancer Prevention, Anti-Inflammatory and Anti-Oxidation of Bioactive Phytochemicals Targeting the TLR4 Signaling Pathway. Int. J. Mol. Sci. 2018, 19, 2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera-Pool, E.; Ramos-Díaz, A.L.; Lizardi-Jiménez, M.A.; Pech-Cohuo, S.; Ayora-Talavera, T.; Cuevas-Bernardino, J.C.; García-Cruz, U.; Pacheco, N. Effect of Solvent Polarity on the Ultrasound Assisted Extraction and Antioxidant Activity of Phenolic Compounds from Habanero Pepper Leaves (Capsicum chinense) and Its Identification by UPLC-PDA-ESI-MS/MS. Ultrason. Sonochem. 2021, 76, 105658. [Google Scholar] [CrossRef] [PubMed]
- Jokela, T.; Kärnä, R.; Rauhala, L.; Bart, G.; Pasonen-Seppänen, S.; Oikari, S.; Tammi, M.I.; Tammi, R.H. Human Keratinocytes Respond to Extracellular UTP by Induction of Hyaluronan Synthase 2 Expression and Increased Hyaluronan Synthesis. J. Biol. Chem. 2017, 292, 4861–4872. [Google Scholar] [CrossRef] [Green Version]
- Warshaw, E.M.; Ruggiero, J.L.; DeKoven, J.G.; Silverberg, J.I.; Maibach, H.I.; Taylor, J.S.; Atwater, A.R.; Zug, K.A.; Sasseville, D.; Fowler, J.F.; et al. Patch Testing With Tocopherol and Tocopherol Acetate: The North American Contact Dermatitis Group Experience, 2001 to 2016. Dermat. Contact Atopic Occup. Drug 2021, 32, 308–318. [Google Scholar] [CrossRef]
- Cavinato, M.; Waltenberger, B.; Baraldo, G.; Grade, C.V.C.; Stuppner, H.; Jansen-Dürr, P. Plant Extracts and Natural Compounds Used against UVB-Induced Photoaging. Biogerontology 2017, 18, 499–516. [Google Scholar] [CrossRef] [Green Version]
- Mohania, D.; Chandel, S.; Kumar, P.; Verma, V.; Digvijay, K.; Tripathi, D.; Choudhury, K.; Mitten, S.K.; Shah, D. Ultraviolet Radiations: Skin Defense-Damage Mechanism. Adv. Exp. Med. Biol. 2017, 996, 71–87. [Google Scholar] [PubMed]
- Zheng, Z.; Xiao, Z.; He, Y.L.; Tang, Y.; Li, L.; Zhou, C.; Hong, P.; Luo, H.; Qian, Z.J. Heptapeptide Isolated from Isochrysis Zhanjiangensis Exhibited Anti-Photoaging Potential via Mapk/Ap-1/Mmp Pathway and Anti-Apoptosis in Uvb-Irradiated Hacat Cells. Mar. Drugs 2021, 19, 626. [Google Scholar] [CrossRef]
- Stern, R.; Maibach, H.I. Hyaluronan in Skin: Aspects of Aging and Its Pharmacologic Modulation. Clin. Dermatol. 2008, 26, 106–122. [Google Scholar] [CrossRef] [PubMed]
- Beck, L.A.; Cork, M.J.; Amagai, M.; De Benedetto, A.; Kabashima, K.; Hamilton, J.D.; Rossi, A.B. Type 2 Inflammation Contributes to Skin Barrier Dysfunction in Atopic Dermatitis. JID Innov. 2022, 2, 100131. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Elias, B.C.; Seth, A.; Shen, L.; Turner, J.R.; Giorgianni, F.; Desiderio, D.; Guntaka, R.; Rao, R. PKCη Regulates Occludin Phosphorylation and Epithelial Tight Junction Integrity. Proc. Natl. Acad. Sci. USA 2009, 106, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Cutler, R.G. Antioxidants and Aging. Am. J. Clin. Nutr. 1991, 53, 373S–379S. [Google Scholar] [CrossRef]
- Jiang, Q. Natural Forms of Vitamin E: Metabolism, Antioxidant, and Anti-Inflammatory Activities and Their Role in Disease Prevention and Therapy. Free Radic. Biol. Med. 2014, 72, 76–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kammeyer, A.; Luiten, R.M. Oxidation Events and Skin Aging. Ageing Res. Rev. 2015, 21, 16–29. [Google Scholar] [CrossRef]
- Van Hung, P. Phenolic Compounds of Cereals and Their Antioxidant Capacity. Crit. Rev. Food Sci. Nutr. 2016, 56, 25–35. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An Overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pietta, P.G. Flavonoids as Antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Dudonné, S.; Vitrac, X.; Coutiére, P.; Woillez, M.; Mérillon, J.M. Comparative Study of Antioxidant Properties and Total Phenolic Content of 30 Plant Extracts of Industrial Interest Using DPPH, ABTS, FRAP, SOD, and ORAC Assays. J. Agric. Food Chem. 2009, 57, 1768–1774. [Google Scholar] [CrossRef]
- Shin, K.K.; Park, S.H.; Lim, H.Y.; Lorza, L.R.; Qomaladewia, N.P.; You, L.; Aziz, N.; Kim, S.A.; Lee, J.S.; Choung, E.S.; et al. In Vitro Anti-Photoaging and Skin Protective Effects of Licania Macrocarpa Cuatrec Methanol Extract. Plants 2022, 11, 1383. [Google Scholar] [CrossRef]
- Kim, S.R.; To, D.C.; Nguyen, P.H.; Nguyen, Y.N.; Cho, B.J.; Tran, M.H. Antioxidant and Cell Proliferation Properties of the Vietnamese Traditional Medicinal Plant Peltophorum Pterocarpum. Molecules 2020, 25, 4800. [Google Scholar] [CrossRef]
- Singh, P.; Vishwakarma, S.P.; Singh, R.L. Antioxidant, Oxidative DNA Damage Protective and Antimicrobial Activities of the Plant Trigonella foenum-graecum. J. Sci. Food Agric. 2014, 94, 2497–2504. [Google Scholar] [CrossRef] [PubMed]
- Hymery, N.; Dauvergne, X.; Boussaden, H.; Cérantola, S.; Faugère, D.; Magné, C. Evaluation of the Antioxidant, Anti-Inflammatory and Cytoprotective Activities of Halophyte Extracts against Mycotoxin Intoxication. Toxins 2021, 13, 312. [Google Scholar] [CrossRef]
- Rubnawaz, S.; Kayani, W.K.; Akhtar, N.; Mahmood, R.; Khan, A.; Okla, M.K.; Alamri, S.A.; Alaraidh, I.A.; Alwasel, Y.A.; Mirza, B. Correction: Rubnawaz et al. Polyphenol Rich Ajuga bracteosa Transgenic Regenerants Display Better Pharmacological Potential. Molecules 2021, 26, 4874. [Google Scholar] [CrossRef]
- Kim, K.H.; Park, Y.J.; Chung, K.H.; Yip, M.L.R.; Clardy, J.; Senger, D.; Cao, S. Iridoid Glycosides from Barleria lupulina. J. Nat. Prod. 2015, 78, 320–324. [Google Scholar] [CrossRef] [PubMed]
- Benrahou, K.; El Guourrami, O.; Mrabti, H.N.; Cherrah, Y.; Faouzi, M.E.A. Investigation of Antioxidant, Hypoglycemic and Anti-Obesity Effects of Euphorbia resinifera L. J. Pharmacopunct. 2022, 25, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Elbouzidi, A.; Ouassou, H.; Aherkou, M.; Kharchoufa, L.; Meskali, N.; Baraich, A.; Mechchate, H.; Bouhrim, M.; Idir, A.; Hano, C.; et al. LC–MS/MS Phytochemical Profiling, Antioxidant Activity, and Cytotoxicity of the Ethanolic Extract of Atriplex halimus L. against Breast Cancer Cell Lines: Computational Studies and Experimental Validation. Pharmaceuticals 2022, 15, 1156. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.H.; Kim, J.H.; Choi, E.; Park, S.H.; Cho, J.Y. Antioxidative and Skin Protective Effects of Canarium subulatum Methanol Extract on Keratinocytes. Evid. Based Complement. Altern. Med. 2021, 2021, 6692838. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Lee, C.Y.; Lee, H.P.; Hossain, M.A.; Zhang, Z.; Kim, S.Y.; Song, M.; Kim, J.H.; Cho, J.Y. Protective Function of Malus baccata (L.) Borkh Methanol Extract against UVB/Hydrogen Peroxide-Induced Skin Aging via Inhibition of MAPK and NF-ΚB Signaling. Plants 2022, 11, 2368. [Google Scholar] [CrossRef]
- Bode, A.M.; Dong, Z. Mitogen-Activated Protein Kinase Activation in UV-Induced Signal Transduction. Sci. STKE 2003, 2003, re2. [Google Scholar] [CrossRef] [PubMed]
- Basuroy, S.; Seth, A.; Elias, B.; Naren, A.P.; Rao, R. MAPK Interacts with Occludin and Mediates EGF-Induced Prevention of Tight Junction Disruption by Hydrogen Peroxide. Biochem. J. 2006, 393, 69–77. [Google Scholar] [CrossRef]
- Akimov, S.S.; Belkin, A.M. Opposing Roles of Ras/Raf Oncogenes and the MEK1/ERK Signaling Module in Regulation of Expression and Adhesive Function of Surface Transglutaminase. J. Biol. Chem. 2003, 278, 35609–35619. [Google Scholar] [CrossRef] [Green Version]
- Yuki, T.; Hachiya, A.; Kusaka, A.; Sriwiriyanont, P.; Visscher, M.O.; Morita, K.; Muto, M.; Miyachi, Y.; Sugiyama, Y.; Inoue, S. Characterization of Tight Junctions and Their Disruption by UVB in Human Epidermis and Cultured Keratinocytes. J. Investig. Dermatol. 2011, 131, 744–752. [Google Scholar] [CrossRef] [Green Version]
- Kagan, V.E.; Fabisiak, J.P.; Shvedova, A.A.; Tyurina, Y.Y.; Tyurin, V.A.; Schor, N.F.; Kawai, K. Oxidative Signaling Pathway for Externalization of Plasma Membrane Phosphatidylserine during Apoptosis. FEBS Lett. 2000, 477, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Gęgotek, A.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Cytoprotective Effect of Ascorbic Acid and Rutin against Oxidative Changes in the Proteome of Skin Fibroblasts Cultured in a Three-Dimensional System. Nutrients 2020, 12, 1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gęgotek, A.; Ambrożewicz, E.; Jastrząb, A.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Rutin and Ascorbic Acid Cooperation in Antioxidant and Antiapoptotic Effect on Human Skin Keratinocytes and Fibroblasts Exposed to UVA and UVB Radiation. Arch. Dermatol. Res. 2019, 311, 203–219. [Google Scholar] [CrossRef] [Green Version]
- Martins, R.M.; de Siqueira Martins, S.; Barbosa, G.L.F.; Fonseca, M.J.V.; Rochette, P.J.; Moulin, V.J.; de Freitas, L.A.P. Photoprotective Effect of Solid Lipid Nanoparticles of Rutin against UVB Radiation Damage on Skin Biopsies and Tissue-Engineered Skin. J. Microencapsul. 2022, 39, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Her, Y.; Lee, T.K.; Kim, J.D.; Kim, B.; Sim, H.; Lee, J.C.; Ahn, J.H.; Park, J.H.; Lee, J.W.; Hong, J.; et al. Topical Application of Aronia Melanocarpa Extract Rich in Chlorogenic Acid and Rutin Reduces UVB-Induced Skin Damage via Attenuating Collagen Disruption in Mice. Molecules 2020, 25, 4577. [Google Scholar] [CrossRef] [PubMed]
- Martins, R.M.; Alves, G.D.A.D.; Martins, S.D.S.; De Freitas, L.A.P.; Rochette, P.J.; Moulin, V.J.; Fonseca, M.J.V. Apple Extract (Malus Sp.) and Rutin as Photochemopreventive Agents: Evaluation of Ultraviolet B-Induced Alterations on Skin Biopsies and Tissue-Engineered Skin. Rejuvenation Res. 2020, 23, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Bimbiraitė-Survilienė, K.; Stankevičius, M.; Šuštauskaitė, S.; Gęgotek, A.; Maruška, A.; Skrzydlewska, E.; Barsteigienė, Z.; Akuņeca, I.; Ragažinskienė, O.; Lukošius, A. Evaluation of Chemical Composition, Radical Scavenging and Antitumor Activities of Satureja hortensis L. Herb Extracts. Antioxidants 2021, 10, 53. [Google Scholar] [CrossRef]
- Seo, C.; Lym, S.H.; Jeong, W.; Lee, J.E.; Lee, J.A.; Ahn, E.K.; Kang, J.S.; Kim, W.H.; Choi, C.W.; Oh, J.S.; et al. Flavonoids, Stilbenoids, and Phenolic Derivatives from the Stems of Gnetum macrostachyum (Gnetaceae). Biochem. Syst. Ecol. 2020, 90, 104033. [Google Scholar] [CrossRef]
- Kim, H.G.; Choi, S.; Lee, J.; Hong, Y.H.; Jeong, D.; Yoon, K.; Yoon, D.H.; Sung, G.H.; Lee, S.; Hong, S.; et al. Src Is a Prime Target Inhibited by Celtis Choseniana Methanol Extract in Its Anti-Inflammatory Action. Evid. Based Complement. Altern. Med. 2018, 2018, 3909038. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.H.; Song, C.; Shin, K.K.; Choi, E.; Hwang, S.H.; Jang, Y.J.; Taamalli, A.; Yum, J.; Kim, J.H.; Kim, E.; et al. Tunisian Olea europaea L. Leaf Extract Suppresses Freund’s Complete Adjuvant-Induced Rheumatoid Arthritis and Lipopolysaccharide-Induced Inflammatory Responses. J. Ethnopharmacol. 2021, 268, 113602. [Google Scholar] [CrossRef]
- Kim, J.K.; Choi, E.; Hong, Y.H.; Kim, H.; Jang, Y.J.; Lee, J.S.; Choung, E.S.; Woo, B.Y.; Hong, Y.D.; Lee, S.; et al. Syk/NF-ΚB-Targeted Anti-Inflammatory Activity of Melicope Accedens (Blume) T.G. Hartley Methanol Extract. J. Ethnopharmacol. 2021, 271, 113887. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, J.G.; Hong, Y.H.; Shin, K.K.; Kim, J.K.; Kim, Y.D.; Yoon, K.D.; Kim, K.H.; Yoo, B.C.; Sung, G.H.; et al. Sauropus Brevipes Ethanol Extract Negatively Regulates Inflammatory Responses in Vivo and in Vitro by Targeting Src, Syk and IRAK1. Pharm. Biol. 2021, 59, 74–86. [Google Scholar] [CrossRef]
- Dajanta, K.; Janpum, P.; Leksing, W. Antioxidant Capacities, Total Phenolics and Flavonoids in Black and Yellow Soybeans Fermented by Bacillus subtilis: A Comparative Study of Thai Fermented Soybeans (Thua Nao). Int. Food Res. J. 2013, 20, 3125–3132. [Google Scholar]
- Kondakçi, E.; Özyürek, M.; Güçlü, K.; Apak, R. Novel Pro-Oxidant Activity Assay for Polyphenols, Vitamins C and e Using a Modified CUPRAC Method. Talanta 2013, 115, 583–589. [Google Scholar] [CrossRef]
- Song, C.; Jeong, D.; Hong, Y.H.; Li, W.Y.; Lee, S.W.; Hossain, M.A.; Taamalli, A.; Kim, J.H.; Kim, J.H.; Cho, J.Y. Anti-Inflammatory and Photoaging-Protective Effects of Olea Europaea through Inhibition of AP-1 and NF- κ B Pathways. Am. J. Chin. Med. 2020, 48, 1895–1913. [Google Scholar] [CrossRef] [PubMed]
- Naylor, L.H. Reporter Gene Technology: The Future Looks Bright. Biochem. Pharmacol. 1999, 58, 749–757. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Shin, K.K.; Kim, H.; Hong, Y.H.; Choi, W.; Kwak, Y.S.; Han, C.K.; Hyun, S.H.; Cho, J.Y. Korean Red Ginseng Exerts Anti-Inflammatory and Autophagy-Promoting Activities in Aged Mice. J. Ginseng Res. 2021, 45, 717–725. [Google Scholar] [CrossRef]
- Ha, A.T.; Rahmawati, L.; You, L.; Hossain, M.A.; Kim, J.H.; Cho, J.Y. Anti-Inflammatory, Antioxidant, Moisturizing, and Antimelanogenesis Effects of Quercetin 3-o-β-d-Glucuronide in Human Keratinocytes and Melanoma Cells via Activation of Nf-Κb and Ap-1 Pathways. Int. J. Mol. Sci. 2022, 23, 433. [Google Scholar] [CrossRef]
- Hong, Y.H.; Kim, J.H.; Cho, J.Y. Photoaging Protective Effects of Ranunculus Bulumei Methanol Extract. Evid. Based Complement. Altern. Med. 2020, 2020, 1761785. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.O.; Yang, Y.; Tao, Y.; Yi, Y.S.; Cho, J.Y. Korean Red Ginseng Saponin Fraction Exerts Anti-Inflammatory Effects by Targeting the NF-ΚB and AP-1 Pathways. J. Ginseng Res. 2022, 46, 489–495. [Google Scholar] [CrossRef]
- Lee, J.O.; Kim, J.H.; Kim, S.; Kim, M.Y.; Hong, Y.H.; Kim, H.G.; Cho, J.Y. Gastroprotective Effects of the Nonsaponin Fraction of Korean Red Ginseng through Cyclooxygenase-1 Upregulation. J. Ginseng Res. 2020, 44, 655–663. [Google Scholar] [CrossRef]
- Choi, E.; Yi, Y.S.; Lee, J.; Park, S.H.; Kim, S.; Hossain, M.A.; Jang, S.G.; Choi, Y.I.; Park, K.J.; Kim, D.S.; et al. Anti-Apoptotic and Anti-Inflammatory Activities of Edible Fresh Water Algae Prasiola japonica in UVB-Irradiated Skin Keratinocytes. Am. J. Chin. Med. 2019, 47, 1853–1868. [Google Scholar] [CrossRef]
- Song, C.; Lorz, L.R.; Lee, J.; Cho, J.Y. In Vitro Photoprotective, Anti-Inflammatory, Moisturizing, and Antimelanogenic Effects of a Methanolic Extract of Chrysophyllum lucentifolium Cronquist. Plants 2022, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Hwang, K.; Lee, J.; Han, S.Y.; Kim, E.M.; Park, J.; Cho, J.Y. Skin Protective Effect of Epigallocatechin Gallate. Int. J. Mol. Sci. 2018, 19, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Cho, J.Y.; Lee, S.Y.; Lee, K.W.; Lee, J.; Song, J.Y. Vanillin Protects Human Keratinocyte Stem Cells against Ultraviolet B Irradiation. Food Chem. Toxicol. 2014, 63, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Oh, S.W.; Shin, S.W.; Lee, K.W.; Cho, J.Y.; Lee, J. Zingerone Protects Keratinocyte Stem Cells from UVB-Induced Damage. Chem. Biol. Interact. 2018, 279, 27–33. [Google Scholar] [CrossRef]
- Han, S.Y.; Kim, E.; Hwang, K.; Ratan, Z.A.; Hwang, H.; Kim, E.M.; Kim, D.; Park, J.; Cho, J.Y. Cytoprotective Effect of Epigallocatechin Gallate (EGCG)-5′-O-α-Glucopyranoside, a Novel EGCG Derivative. Int. J. Mol. Sci. 2018, 19, 1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.O.; Kim, E.; Kim, J.H.; Hong, Y.H.; Kim, H.G.; Jeong, D.; Kim, J.; Kim, S.H.; Park, C.; Seo, D.B.; et al. Antimelanogenesis and Skin-Protective Activities of Panax Ginseng Calyx Ethanol Extract. J. Ginseng Res. 2018, 42, 389–399. [Google Scholar] [CrossRef]
- Jeong, D.; Lee, J.; Jeong, S.G.; Hong, Y.H.; Yoo, S.; Han, S.Y.; Kim, J.H.; Kim, S.; Kim, J.S.; Chung, Y.S.; et al. Artemisia Asiatica Ethanol Extract Exhibits Anti-Photoaging Activity. J. Ethnopharmacol. 2018, 220, 57–66. [Google Scholar] [CrossRef]
- Choi, W.; Kim, H.S.; Park, S.H.; Kim, D.; Hong, Y.D.; Kim, J.H.; Cho, J.Y. Syringaresinol Derived from Panax Ginseng Berry Attenuates Oxidative Stress-Induced Skin Aging via Autophagy. J. Ginseng Res. 2022, 46, 536–542. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PCR Type | Gene Name | Sequence (5′-3′) | |
|---|---|---|---|
| RT-PCR | Occludin | Forward | GAAGATGAGGATGGCTGTCA |
| Reverse | AAATTCGTACCTGGCATTGA | ||
| HAS-3 | Forward | TATACCGCGCGCTCCAA | |
| Reverse | GCCACTCCCGGAAGTAAGACT | ||
| TGM-1 | Forward | GAAATGCGGCAGATGACGAC | |
| Reverse | AACTCCCCAGCGTCTGATTG | ||
| HAS-1 | Forward | CCACCCAGTACAGCGTCAAC | |
| Reverse | CATGGTGCTTCTGTCGCTCT | ||
| Claudin | Forward | AGGAACACATTTATGATGAGCAG | |
| Reverse | GAAGTCATCCACAGGCGAA | ||
| HAS-2 | Forward | TTCTTTATGTGACTCATCTGTCTCACCGG | |
| Reverse | ATTGTTGGCTACCAGTTTATCCAAACG | ||
| GAPDH | Forward | GACAGTCAGCCGCATCTTCT | |
| Reverse | GCGCCAATACGACCAAATC | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, L.; You, L.; Aziz, N.; Yu, S.H.; Lee, J.S.; Choung, E.S.; Luong, V.D.; Jeon, M.-J.; Hur, M.; Lee, S.; et al. Antiphotoaging and Skin-Protective Activities of Ardisia silvestris Ethanol Extract in Human Keratinocytes. Plants 2023, 12, 1167. https://doi.org/10.3390/plants12051167
Huang L, You L, Aziz N, Yu SH, Lee JS, Choung ES, Luong VD, Jeon M-J, Hur M, Lee S, et al. Antiphotoaging and Skin-Protective Activities of Ardisia silvestris Ethanol Extract in Human Keratinocytes. Plants. 2023; 12(5):1167. https://doi.org/10.3390/plants12051167
Chicago/Turabian StyleHuang, Lei, Long You, Nur Aziz, Seung Hui Yu, Jong Sub Lee, Eui Su Choung, Van Dung Luong, Mi-Jeong Jeon, Moonsuk Hur, Sarah Lee, and et al. 2023. "Antiphotoaging and Skin-Protective Activities of Ardisia silvestris Ethanol Extract in Human Keratinocytes" Plants 12, no. 5: 1167. https://doi.org/10.3390/plants12051167