



Impacts of Radio-Frequency Electromagnetic Field (RF-EMF) on Lettuce (Lactuca sativa)—Evidence for RF-EMF Interference with Plant Stress Responses


Abstract
:1. Introduction
2. Results
2.1. Prolonged RF-EMF Exposure Caused Changes in Plant Fast Chlorophyll Fluorescence Kinetics
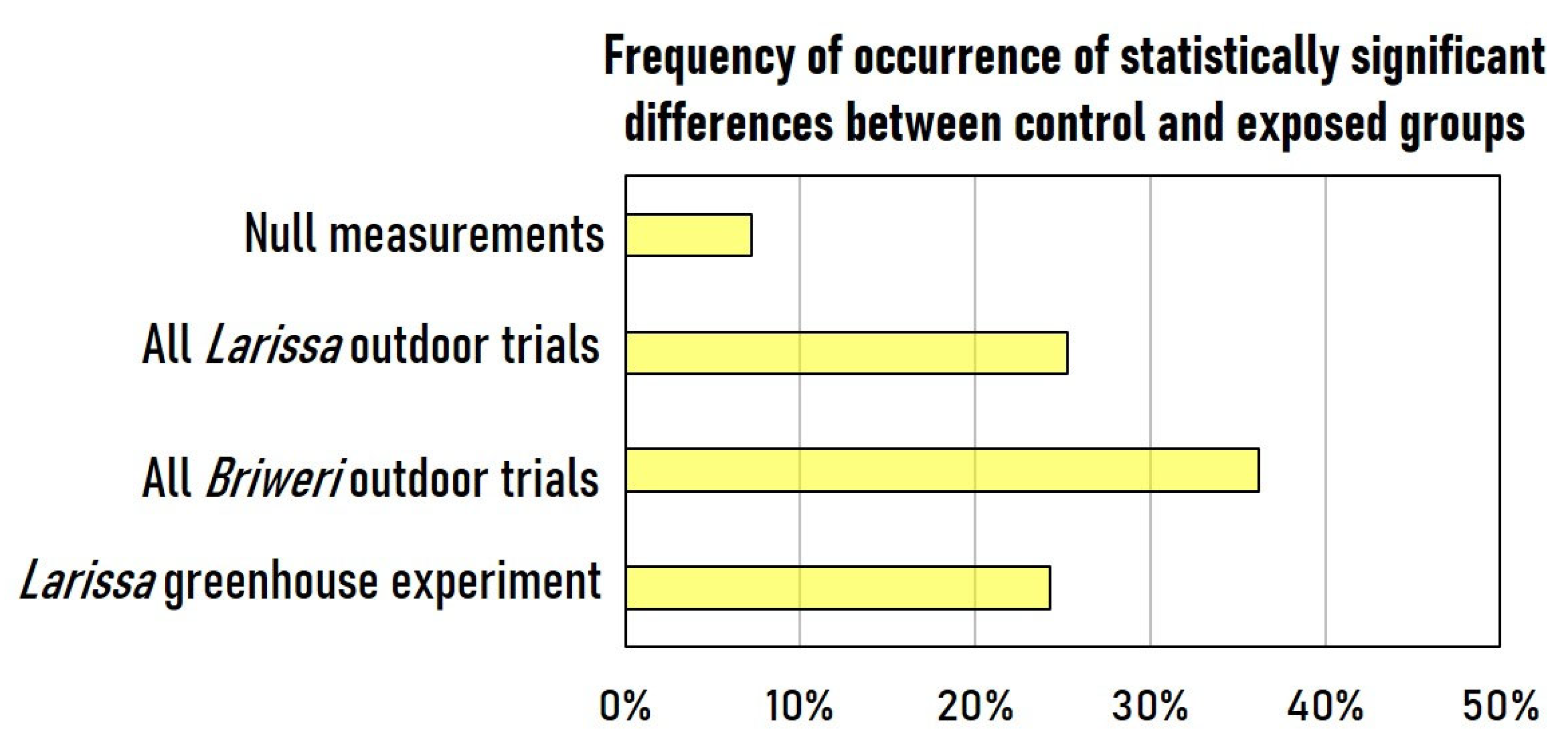
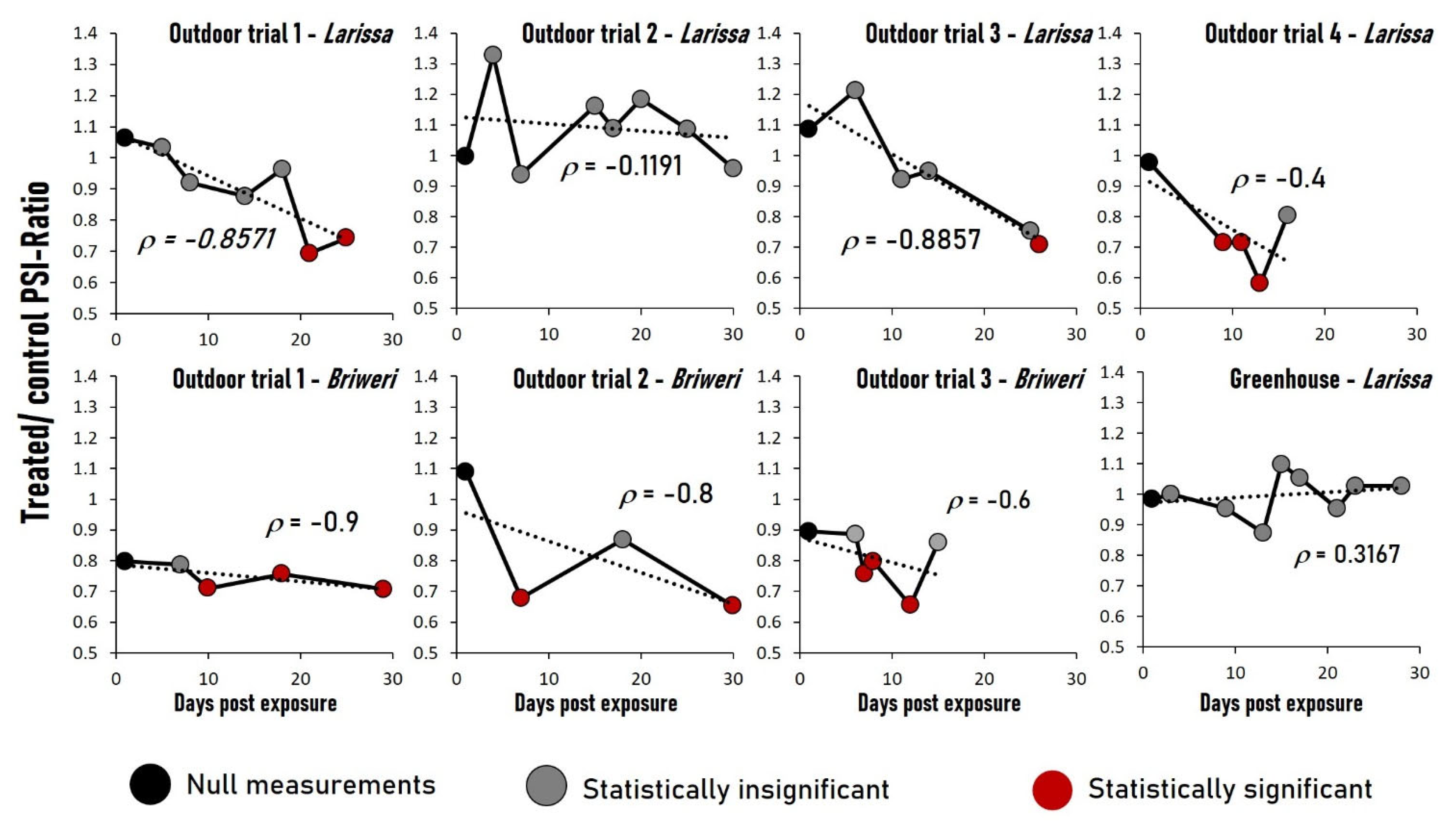
2.2. Continuous RF-EMF Exposure Resulted in Significant Decrease of Photosynthetic Efficiency in Six out of Seven Outdoor Trials but Not in the Greenhouse Experiment
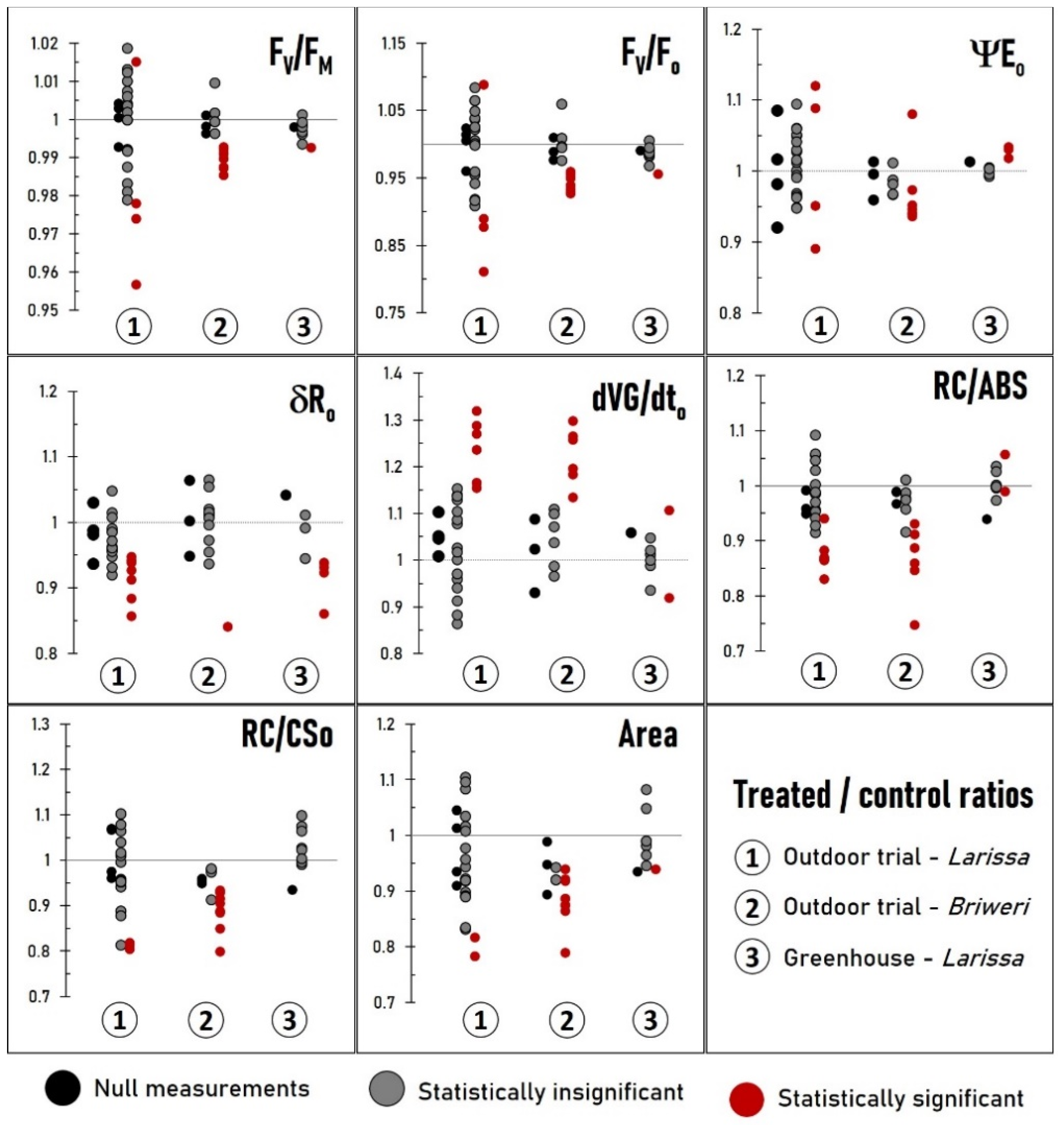
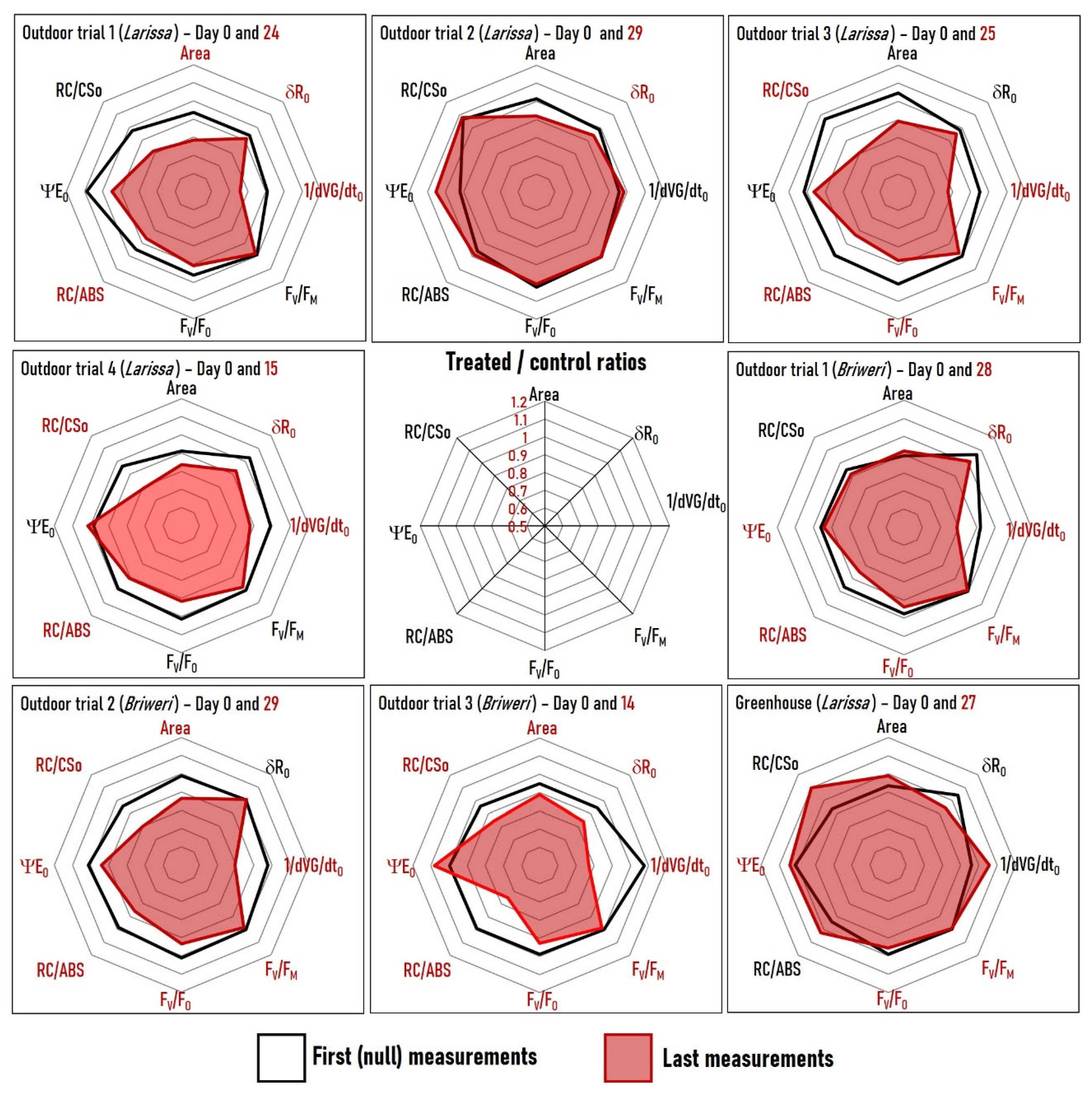
2.3. Analysis of Individual OJIP Parameters Reveals That Many Photosynthetic Processes Are Simultaneously Affected by RF-EMF
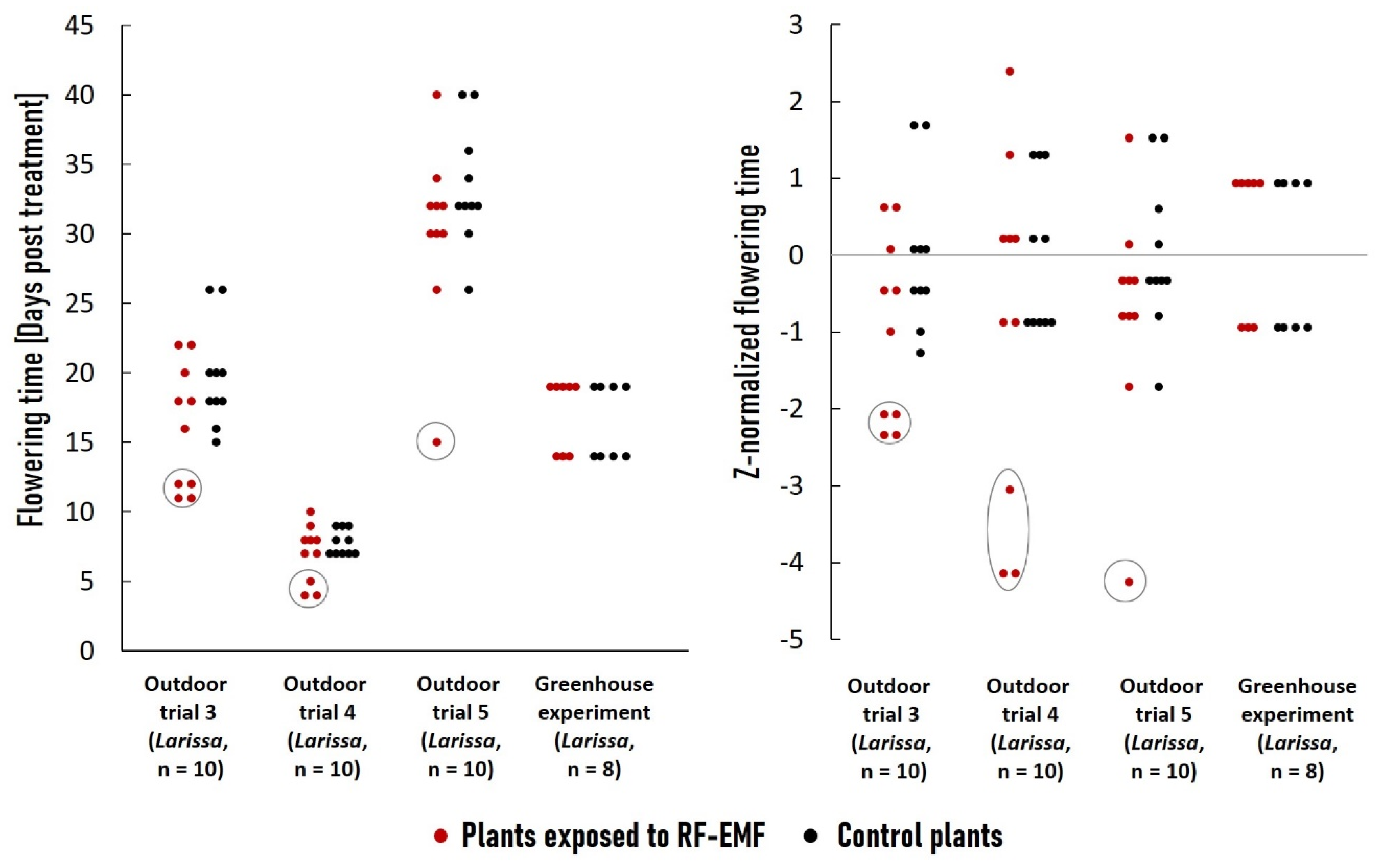
2.4. Long-Term RF-EMF Exposure Led to Accelerated Flowering Time in Outdoor Environments Only
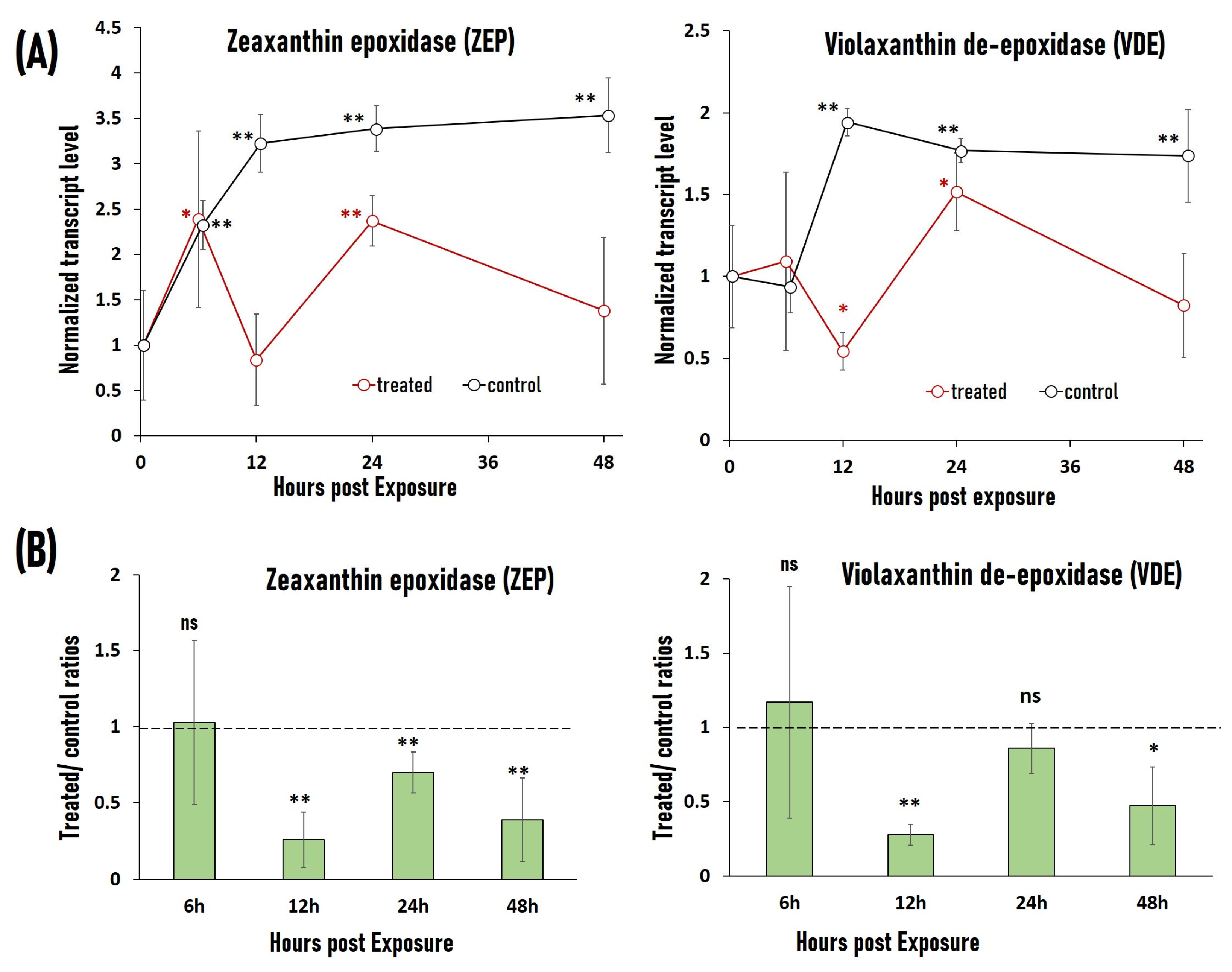
2.5. RF-EMF Exposure Causes Downregulation of Stress-Related Genes VDE and ZEP
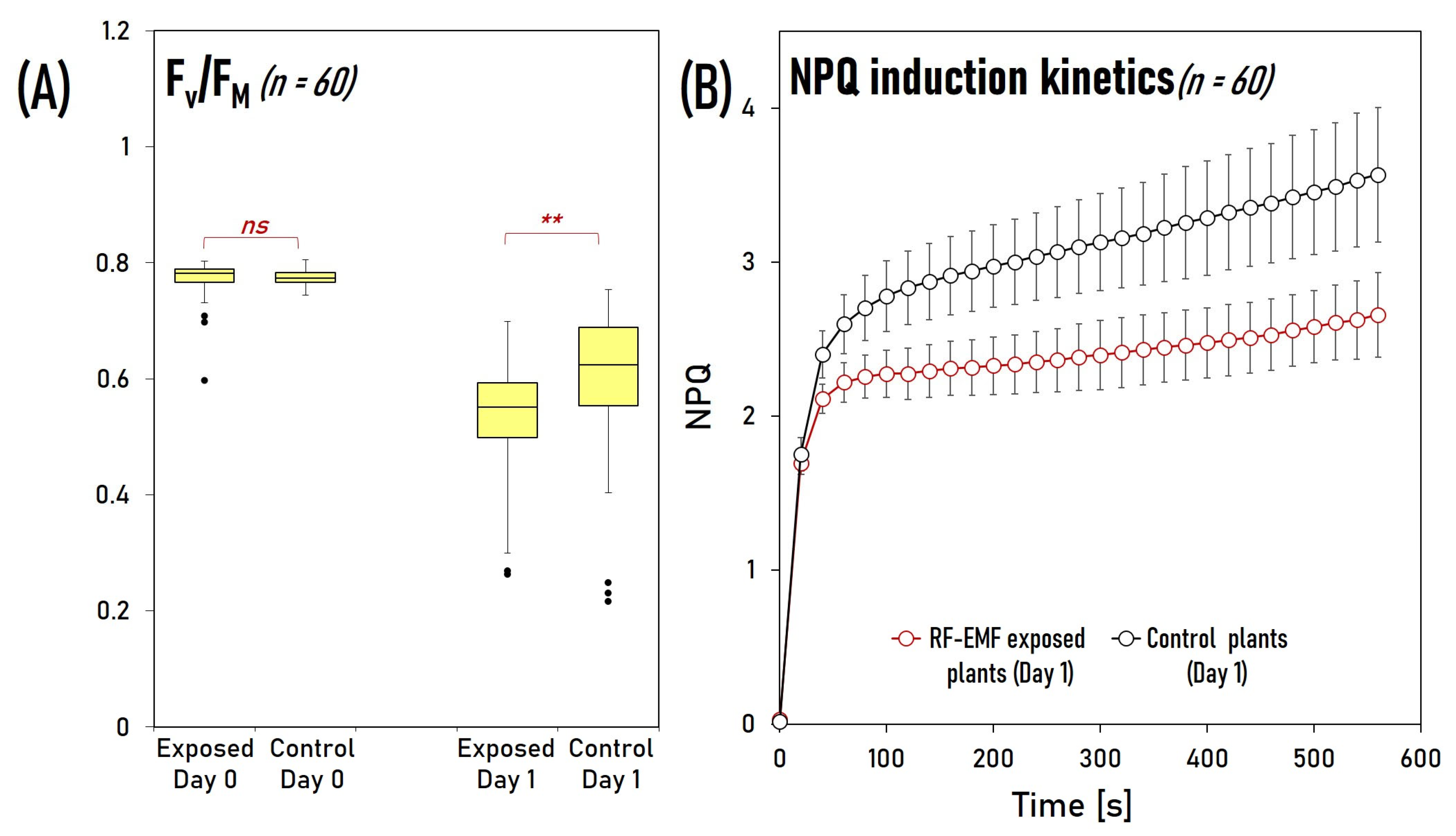
2.6. RF-EMF Exposure Reduces Photosystem II’s Maximal Photochemical Quantum Yield and Non-Photochemical Quenching under Light Stress Conditions
3. Discussion
4. Material and Methods
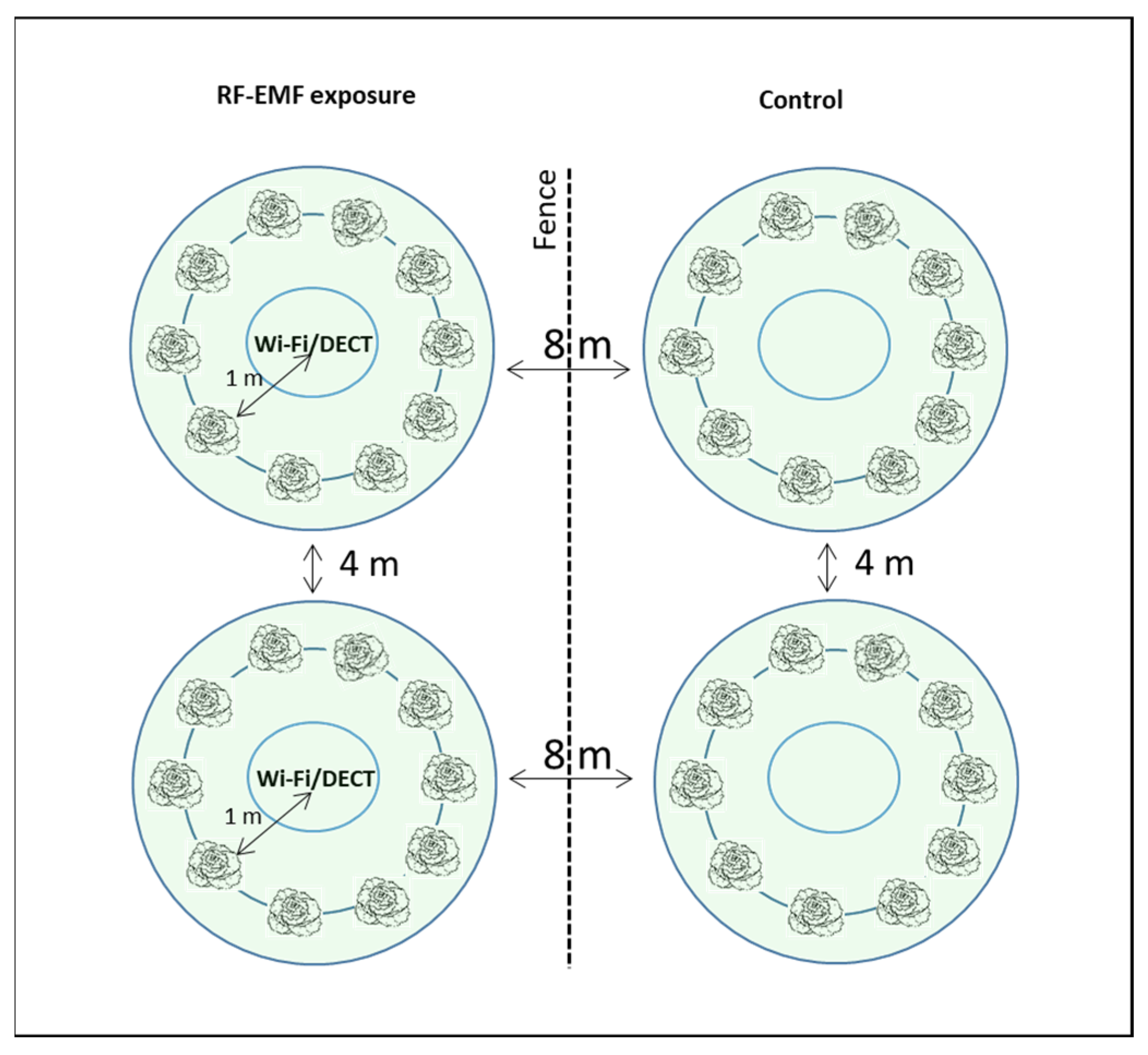
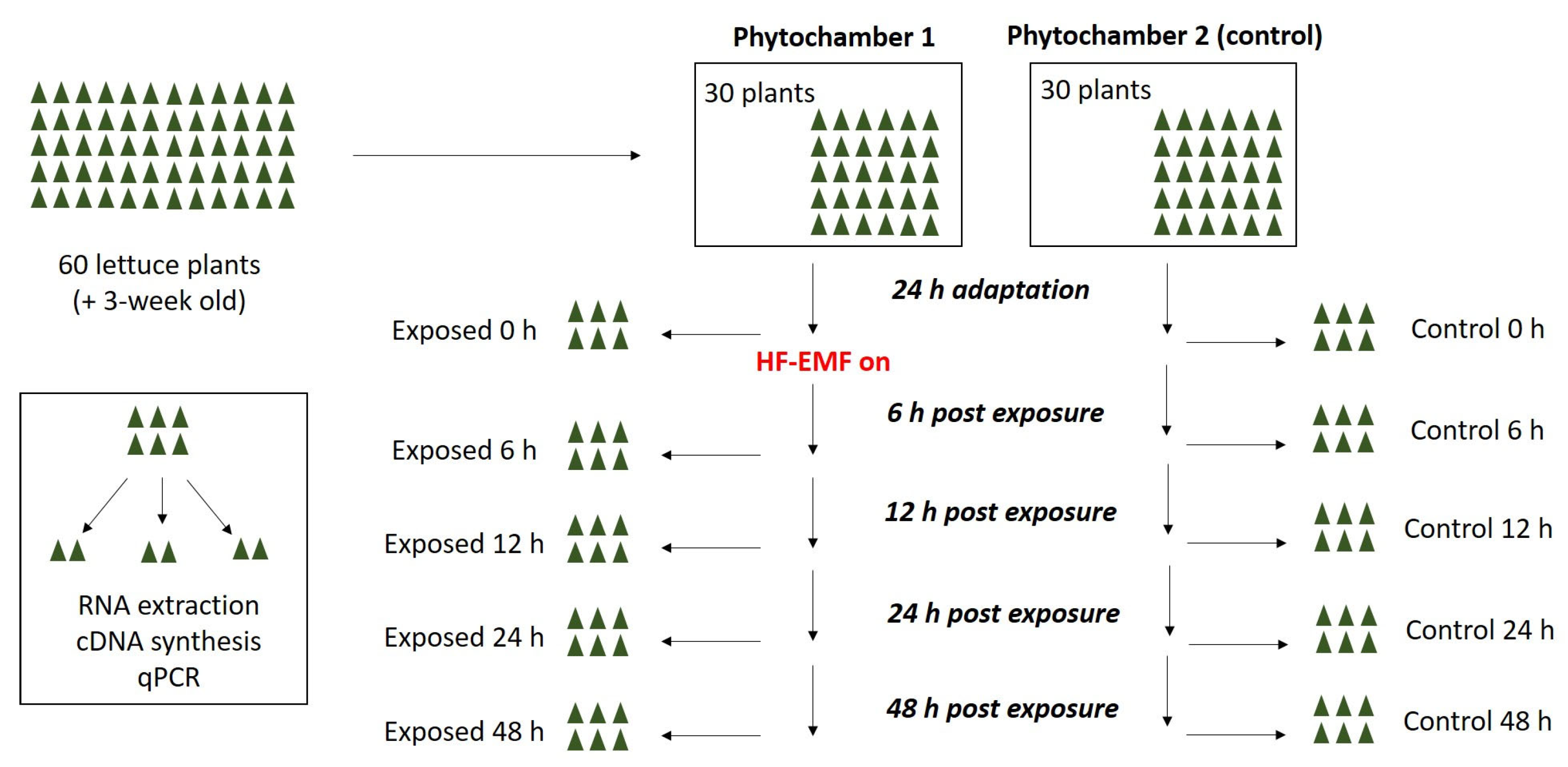
4.1. Overview of the Experimental Plan
4.2. Plant Cultivation
4.3. RF-EMF Exposure
4.4. Gene Expression Analysis
4.5. Flowering Time Analysis
4.6. Measurements of Fast Chlorophyll Fluorescence Kinetics
4.7. Light Stress Experiment and Imaging PAM Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Panagopoulos, D.J.; Johansson, O.; Carlo, G.L. Polarization: A Key Difference between Man-made and Natural Electromagnetic Fields, in regard to Biological Activity. Sci. Rep. 2015, 5, 14914. [Google Scholar] [CrossRef] [Green Version]
- Kaur, S.; Vian, A.; Chandel, S.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Sensitivity of plants to high frequency electromagnetic radiation: Cellular mechanisms and morphological changes. Rev. Environ. Sci. Bio/Technol. 2021, 20, 55–74. [Google Scholar] [CrossRef]
- Pourlis, A.F. Reproductive and developmental effects of EMF in vertebrate animal models. Pathophysiology 2009, 16, 179–189. [Google Scholar] [CrossRef] [PubMed]
- International Agency for Research on Cancer (IARC). IARC Classifies Radiofrequency Electromagnetic Fields as Possibly Carcinogenic to Humans; World Health Organization. 2011, pp. 1–6. Available online: https://www.iarc.who.int/wp-content/uploads/2018/07/pr208_E.pdf (accessed on 4 January 2023).
- Cucurachi, S.; Tamis, W.; Vijver, M.; Peijnenburg, W.; Bolte, J.; de Snoo, G. A review of the ecological effects of radiofrequency electromagnetic fields (RF-EMF). Environ. Int. 2013, 51, 116–140. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration. Scientific Evidence for Cell Phone Safety. Available online: www.fda.gov/radiation-emitting-products/cell-phones/scientific-evidence-cell-phone-safety (accessed on 4 January 2023).
- U.S. Food and Drug Administration. Letter from the FDA to the FCC on Radiofrequency Exposure. 2019. Available online: https://fda.report/media/135022/Letter+from+the+FDA+to+the+FCC+on+Radiofrequency+Exposure_1.pdf (accessed on 4 January 2023).
- U.S. Federal Communications Commission. Policy on Human Exposure to Radiofrequency Electromagnetic Fields. Available online: https://www.fcc.gov/general/fcc-policy-human-exposure (accessed on 4 January 2023).
- Council of the European Union. EU Recommendation 1999/519/EC on the Limitation of Exposure of the General Public to Electromagnetic Fields (0 Hz to 300 GHz). 1999. Available online: https://eur-lex.europa.eu/eli/reco/1999/519/oj (accessed on 4 January 2023).
- Halgamuge, M.N. Review: Weak radiofrequency radiation exposure from mobile phone radiation on plants. Electromagn. Biol. Med. 2016, 36, 213–235. [Google Scholar] [CrossRef]
- Vian, A.; Faure, C.; Girard, S.; Davies, E.; Hallé, F.; Bonnet, P.; Ledoigt, G.; Paladian, F. Plants Respond to GSM-Like Radiations. Plant Signal. Behav. 2007, 2, 522–524. [Google Scholar] [CrossRef] [Green Version]
- Roux, D.; Vian, A.; Girard, S.; Bonnet, P.; Paladian, F.; Davies, E.; Ledoigt, G. High frequency (900 MHz) low amplitude (5 V m−1) electromagnetic field: A genuine environmental stimulus that affects transcription, translation, calcium and energy charge in tomato. Planta 2007, 227, 883–891. [Google Scholar] [CrossRef]
- Sharma, V.P.; Singh, H.P.; Kohli, R.K.; Batish, D.R. Mobile phone radiation inhibits Vigna radiata (mung bean) root growth by inducing oxidative stress. Sci. Total. Environ. 2009, 407, 5543–5547. [Google Scholar] [CrossRef]
- Tkalec, M.; Malarić, K.; Pavlica, M.; Pevalek-Kozlina, B.; Vidaković-Cifrek, Ž. Effects of radiofrequency electromagnetic fields on seed germination and root meristematic cells of Allium cepa L. Mutat. Res. Toxicol. Environ. Mutagen. 2009, 672, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Vian, A.; Roux, D.; Girard, S.; Bonnet, P.; Paladian, F.; Davies, E.; Ledoigt, G. Microwave Irradiation Affects Gene Expression in Plants. Plant Signal. Behav. 2006, 1, 67–69. [Google Scholar] [CrossRef] [Green Version]
- Roux, D.; Vian, A.; Girard, S.; Bonnet, P.; Paladian, F.; Davies, E.; Ledoigt, G. Electromagnetic fields (900 MHz) evoke consistent molecular responses in tomato plants. Physiol. Plant. 2006, 128, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Beaubois, É.; Girard, S.; Lallechere, S.; Davies, E.; Paladian, F.; Bonnet, P.; Ledoigt, G.; Vian, A. Intercellular communication in plants: Evidence for two rapidly transmitted systemic signals generated in response to electromagnetic field stimulation in tomato. Plant, Cell Environ. 2007, 30, 834–844. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.; Sharma, V.; Kumari, N. Microwave pretreatment of tomato seeds and fruit to enhance plant photosynthesis, nutritive quality and shelf life of fruit. Postharvest Biol. Technol. 2019, 159, 111015. [Google Scholar] [CrossRef]
- Mildaziene, V.; Aleknavičiūtė, V.; Žūkienė, R.; Paužaitė, G.; Naučienė, Z.; Filatova, I.; Lyushkevich, V.; Haimi, P.; Tamošiūnė, I.; Baniulis, D. Treatment of Common Sunflower (Helianthus annus L.) Seeds with Radio-frequency Electromagnetic Field and Cold Plasma Induces Changes in Seed Phytohormone Balance, Seedling Development and Leaf Protein Expression. Sci. Rep. 2019, 9, 6437. [Google Scholar] [CrossRef] [Green Version]
- Engelmann, J.C.; Deeken, R.; Müller, T.; Nimtz, G.; Roelfsema, M.R.G.; Redrich, R. Is gene activity in plant cells affected by UMTS-irradiation? A whole genome approach. Adv. Appl. Bioinform. Chem. 2008, 1, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Magone, I. The effect of electromagnetic radiation from the Skrunda Radio Location Station on Spirodela polyrhiza (L.) Schleiden cultures. Sci. Total. Environ. 1996, 180, 75–80. [Google Scholar] [CrossRef]
- Grémiaux, A.; Girard, S.; Guérin, V.; Lothier, J.; Baluška, F.; Davies, E.; Bonnet, P.; Vian, A. Low-amplitude, high-frequency electromagnetic field exposure causes delayed and reduced growth in Rosa hybrida. J. Plant Physiol. 2016, 190, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Scialabba, A.; Tamburello, C. Microwave effects on germination and growth of radish (Raphanus sativus L.) seedlings. Acta Bot. Gallica 2002, 149, 113–123. [Google Scholar] [CrossRef] [Green Version]
- Tkalec, M.; Malarić, K.; Pevalek-Kozlina, B. Exposure to radiofrequency radiation induces oxidative stress in duckweed Lemna minor L. Sci. Total. Environ. 2007, 388, 78–89. [Google Scholar] [CrossRef]
- Kouzmanova, M.; Dimitrova, M.; Dragolova, D.; Atanasova, G.; Atanasov, N. Alterations in Enzyme Activities in Leaves after Exposure of Plectranthus Sp. Plants to 900 MHZ Electromagnetic Field. Biotechnol. Biotechnol. Equip. 2009, 23, 611–615. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Singh, H.P.; Batish, D.R.; Kaur, S.; Kohli, R.K. EMF radiations (1800 MHz)-inhibited early seedling growth of maize (Zea mays) involves alterations in starch and sucrose metabolism. Protoplasma 2015, 253, 1043–1049. [Google Scholar] [CrossRef]
- Chen, Y.-P.; Jia, J.-F.; Wang, Y.-J. Weak Microwave Can Enhance Tolerance of Wheat Seedlings to Salt Stress. J. Plant Growth Regul. 2009, 28, 381–385. [Google Scholar] [CrossRef]
- Soran, M.-L.; Stan, M.; Niinemets; Copolovici, L. Influence of microwave frequency electromagnetic radiation on terpene emission and content in aromatic plants. J. Plant Physiol. 2014, 171, 1436–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lung, I.; Soran, M.-L.; Opriş, O.; Truşcă, M.R.C.; Niinemets, Ü.; Copolovici, L. Induction of stress volatiles and changes in essential oil content and composition upon microwave exposure in the aromatic plant Ocimum basilicum. Sci. Total. Environ. 2016, 569–570, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Beliaeff, B.; Burgeot, T. Integrated biomarker response: A useful tool for ecological risk assessment. Environ. Toxicol. Chem. 2002, 21, 1316–1322. [Google Scholar] [CrossRef]
- Broeg, K.; Lehtonen, K.K. Indices for the assessment of environmental pollution of the Baltic Sea coasts: Integrated assessment of a multi-biomarker approach. Mar. Pollut. Bull. 2006, 53, 508–522. [Google Scholar] [CrossRef] [Green Version]
- Duarte, B.; Pedro, S.; Marques, J.C.; Adão, H.; Caçador, I. Zostera noltii development probing using chlorophyll a transient analysis (JIP-test) under field conditions: Integrating physiological insights into a photochemical stress index. Ecol. Indic. 2017, 76, 219–229. [Google Scholar] [CrossRef]
- Strasserf, R.J.; Srivastava, A.; Govindjee. POLYPHASIC CHLOROPHYLL a FLUORESCENCE TRANSIENT IN PLANTS AND CYANOBACTERIA*. Photochem. Photobiol. 1995, 61, 32–42. [Google Scholar] [CrossRef]
- Tsimilli-Michael, M.; Strasser, R.J. In vivo Assessment of Stress Impact on Plant’s Vitality: Applications in Detecting and Evaluating the Beneficial Role of Mycorrhization on Host Plants. Mycorrhiza 2008, 679–703. [Google Scholar] [CrossRef]
- Ripoll, J.; Bertin, N.; Bidel, L.P.R.; Urban, L. A User’s View of the Parameters Derived from the Induction Curves of Maximal Chlorophyll a Fluorescence: Perspectives for Analyzing Stress. Front. Plant Sci. 2016, 7, 1679. [Google Scholar] [CrossRef] [Green Version]
- Anjum, N.A.; Duarte, B.; Caçador, I.; Sleimi, N.; Duarte, A.C.; Pereira, E. Biophysical and Biochemical Markers of Metal/Metalloid-Impacts in Salt Marsh Halophytes and Their Implications. Front. Environ. Sci. 2016, 4, 24. [Google Scholar] [CrossRef] [Green Version]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a fluorescence as a tool to monitor physiological status of plants under abiotic stress conditions. Acta Physiol. Plant. 2016, 38, 102. [Google Scholar] [CrossRef] [Green Version]
- Wada, K.C.; Takeno, K. Stress-induced flowering. Plant Signal. Behav. 2010, 5, 944–947. [Google Scholar] [CrossRef]
- Havaux, M.; Niyogi, K.K. The violaxanthin cycle protects plants from photooxidative damage by more than one mechanism. Proc. Natl. Acad. Sci. USA 1999, 96, 8762–8767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.; Han, H.; Feng, H.-L.; Zhao, S.-J.; Meng, Q.-W. Overexpression and Suppression of Violaxanthin De-epoxidase Affects the Sensitivity of Photosystem II Photoinhibition to High Light and Chilling Stress in Transgenic Tobacco. J. Integr. Plant Biol. 2010, 52, 332–339. [Google Scholar] [CrossRef]
- Guan, C.; Ji, J.; Zhang, X.; Li, X.; Jin, C.; Guan, W.; Wang, G. Positive feedback regulation of a Lycium chinense-derived VDE gene by drought-induced endogenous ABA, and over-expression of this VDE gene improve drought-induced photo-damage in Arabidopsis. J. Plant Physiol. 2015, 175, 26–36. [Google Scholar] [CrossRef]
- Na Sun, L.; Wang, F.; Wang, J.W.; Gao, W.R.; Song, X.S. Overexpression of the ChVDE gene, encoding a violaxanthin de-epoxidase, improves tolerance to drought and salt stress in transgenic Arabidopsis. 3 Biotech 2019, 9, 197. [Google Scholar] [CrossRef]
- Marin, E.; Nussaume, L.; Quesada, A.; Gonneau, M.; Sotta, B.; Hugueney, P.; Frey, A.; Marion-Poll, A. Molecular identification of zeaxanthin epoxidase of Nicotiana plumbaginifolia, a gene involved in abscisic acid biosynthesis and corresponding to the ABA locus of Arabidopsis thaliana. EMBO J. 1996, 15, 2331–2342. [Google Scholar] [CrossRef]
- Wang, N.; Fang, W.; Han, H.; Sui, N.; Li, B.; Meng, Q.-W. Overexpression of zeaxanthin epoxidase gene enhances the sensitivity of tomato PSII photoinhibition to high light and chilling stress. Physiol. Plant. 2007, 132, 384–396. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, Y.; Chang, L.; Zhang, T.; An, J.; Liu, Y.; Cao, Y.; Zhao, X.; Sha, X.; Hu, T.; et al. MsZEP, a novel zeaxanthin epoxidase gene from alfalfa (Medicago sativa), confers drought and salt tolerance in transgenic tobacco. Plant Cell Rep. 2015, 35, 439–453. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Schansker, G.; Ladle, R.J.; Goltsev, V.; Bosa, K.; Allakhverdiev, S.I.; Brestic, M.; Bussotti, F.; Calatayud, A.; Dąbrowski, P.; et al. Frequently asked questions about in vivo chlorophyll fluorescence: Practical issues. Photosynth. Res. 2014, 122, 121–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vian, A.; Henry-Vian, C.; Davies, E. Rapid and Systemic Accumulation of Chloroplast mRNA-Binding Protein Transcripts after Flame Stimulus in Tomato. Plant Physiol. 1999, 121, 517–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kollist, H.; Zandalinas, S.I.; Sengupta, S.; Nuhkat, M.; Kangasjärvi, J.; Mittler, R. Rapid Responses to Abiotic Stress: Priming the Landscape for the Signal Transduction Network. Trends Plant Sci. 2018, 24, 25–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebrasseur, N.D.; MacIntosh, G.; Perez-Amador, M.A.; Saitoh, M.; Green, P.J. Local and systemic wound-induction of RNase and nuclease activities in Arabidopsis: RNS1 as a marker for a JA-independent systemic signaling pathway. Plant J. 2002, 29, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Koppel, T.; Ahonen, M.; Carlberg, M.; Hardell, L. Very high radiofrequency radiation at Skeppsbron in Stockholm, Sweden from mobile phone base station antennas positioned close to pedestrians’ heads. Environ. Res. 2022, 208, 112627. [Google Scholar] [CrossRef]
- Panda, D.; Rao, D.N.; Sharma, S.G.; Strasser, R.J.; Sarkar, R.K. Submergence effects on rice genotypes during seedling stage: Probing of submergence driven changes of photosystem 2 by chlorophyll a fluorescence induction O-J-I-P transients. Photosynthetica 2006, 44, 69–75. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RF-EFM Sources | Frequency Range |
|---|---|
| Natural | |
| Lightning discharge | <30 MHz |
| Sun | >30 MHz |
| Man-made | |
| AM radio | 550–1600 MHz |
| FM radio | 88–108 MHz |
| Mobile phones | 800–5500 MHz |
| Wi-Fi system | 2.4 GHz, 5.8 GHz |
| Smart meters | 902–928 MHz |
| Microwave ovens | 2.45 GHz |
| Name | Time | Location | Lettuce Cultivar | Number of Plants (Treated/Control) | Exposure Duration | Analysis |
|---|---|---|---|---|---|---|
| Gene expression analysis | August 2022 | Indoor | Larissa | 30/30 | Up to 48 h | G |
| Outdoor trial 1 (Larissa) | August–September 2021 | Outdoor | Larissa | 10/10 | 4 weeks | O |
| Outdoor trial 2 (Larissa) | October–November 2021 | Outdoor | Larissa | 10/10 | 5 weeks | O |
| Outdoor trial 3 (Larissa) | June–July 2022 | Outdoor | Larissa | 10/10 | 4 weeks | F, O |
| Outdoor trial 4 (Larissa) | August 2022 | Outdoor | Larissa | 10/10 | 2 weeks | F, O |
| Outdoor trial 5 (Larissa) | September–October 2022 | Outdoor | Larissa | 10/10 | 6 weeks | F |
| Outdoor trial 1 (Briweri) | June–July 2022 | Outdoor | Briweri | 10/10 | 4 weeks | O |
| Outdoor trial 2 (Briweri) | June–July 2022 | Outdoor | Briweri | 10/10 | 4 weeks | O |
| Outdoor trial 3 (Briweri) | August 2022 | Outdoor | Briweri | 10/10 | 2 weeks | O |
| Greenhouse experiment (Larissa) | September 2022 | Indoor | Larissa | 9/9 | 4 weeks | F, O |
| Light stress experiment | February 2023 | Indoor | Larissa | 6/6 | 24 h | P |
| Primer | Sequence |
|---|---|
| Actin forward primer | TAC ATG TTC ACC ACC ACA GC |
| Actin reverse primer | ATG AGC TGG ACT TGG CGG TTT |
| VDE forward primer | CGT TTC GAC CTC GGT AGT ATA CG |
| VDE reverse primer | CTG CTG TGC AAA CAT TTG TTC AAG |
| ZEP forward primer | TCT GAT GTT GGG GGT GGA AA |
| ZEP reverse primer | TCC GTC GCA AGC AAC AAA T |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tran, N.T.; Jokic, L.; Keller, J.; Geier, J.U.; Kaldenhoff, R. Impacts of Radio-Frequency Electromagnetic Field (RF-EMF) on Lettuce (Lactuca sativa)—Evidence for RF-EMF Interference with Plant Stress Responses. Plants 2023, 12, 1082. https://doi.org/10.3390/plants12051082
Tran NT, Jokic L, Keller J, Geier JU, Kaldenhoff R. Impacts of Radio-Frequency Electromagnetic Field (RF-EMF) on Lettuce (Lactuca sativa)—Evidence for RF-EMF Interference with Plant Stress Responses. Plants. 2023; 12(5):1082. https://doi.org/10.3390/plants12051082
Chicago/Turabian StyleTran, Nam Trung, Luca Jokic, Julian Keller, Jens Uwe Geier, and Ralf Kaldenhoff. 2023. "Impacts of Radio-Frequency Electromagnetic Field (RF-EMF) on Lettuce (Lactuca sativa)—Evidence for RF-EMF Interference with Plant Stress Responses" Plants 12, no. 5: 1082. https://doi.org/10.3390/plants12051082