
Spring Wheat’s Ability to Utilize Nitrogen More Effectively Is Influenced by Root Phene Variation

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Experiment I
2.2.1. Hydroponics Plant Culture
2.2.2. Measurements
2.3. Experiment II
3. Statistical Analysis
4. Results
4.1. Experiment I
4.1.1. Analysis of Variance and Variability Parameters
4.1.2. Mean Performance, Broad Sense Heritability, and Genetic Advance
4.1.3. Diversity Analysis of Genotypes under High N
4.1.4. Diversity Analysis of Genotypes under Low N
4.1.5. Principal Component Analysis
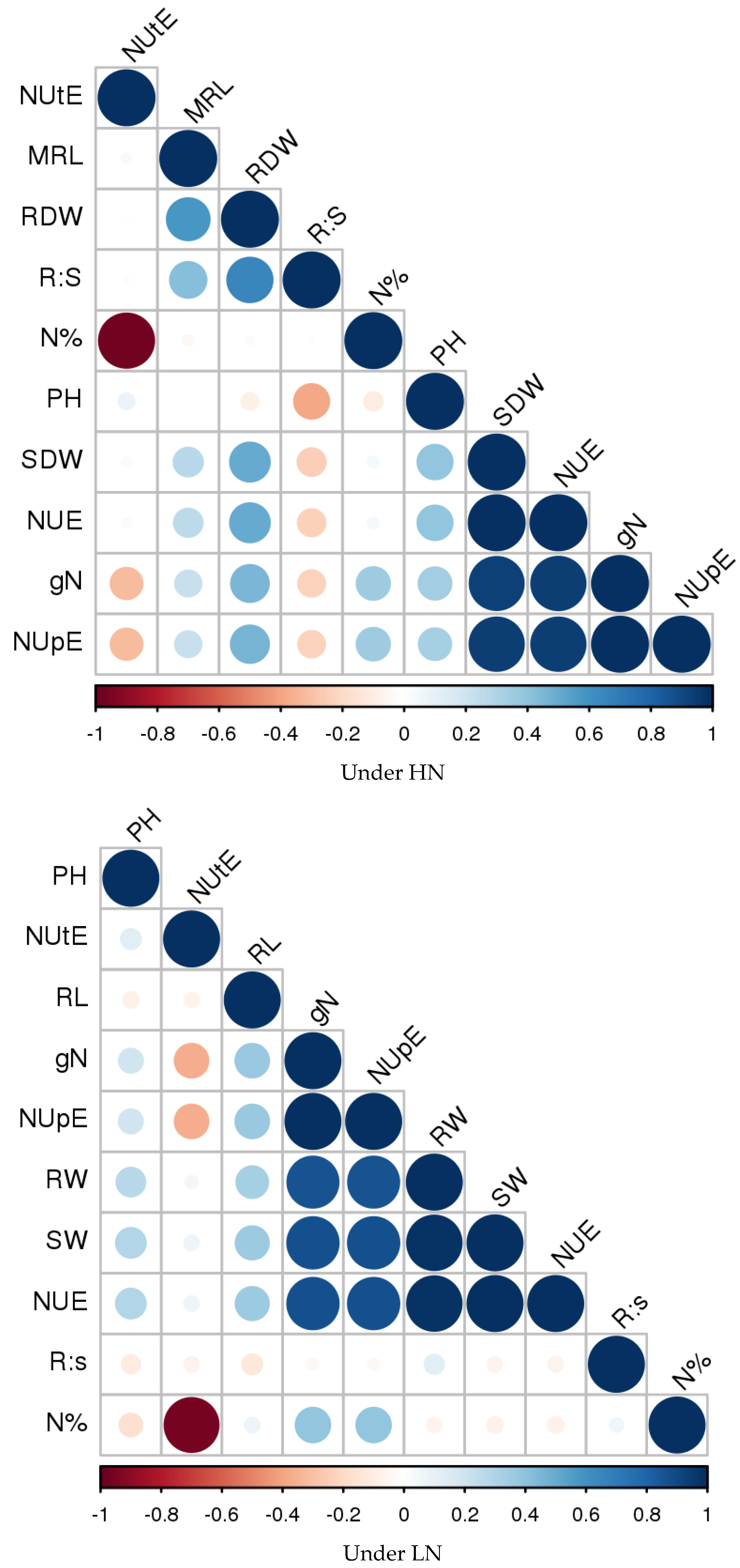
4.1.6. Association Analysis
4.2. Experiment II
4.2.1. Root Traits of the Subset Genotypes under HN and LN Environments
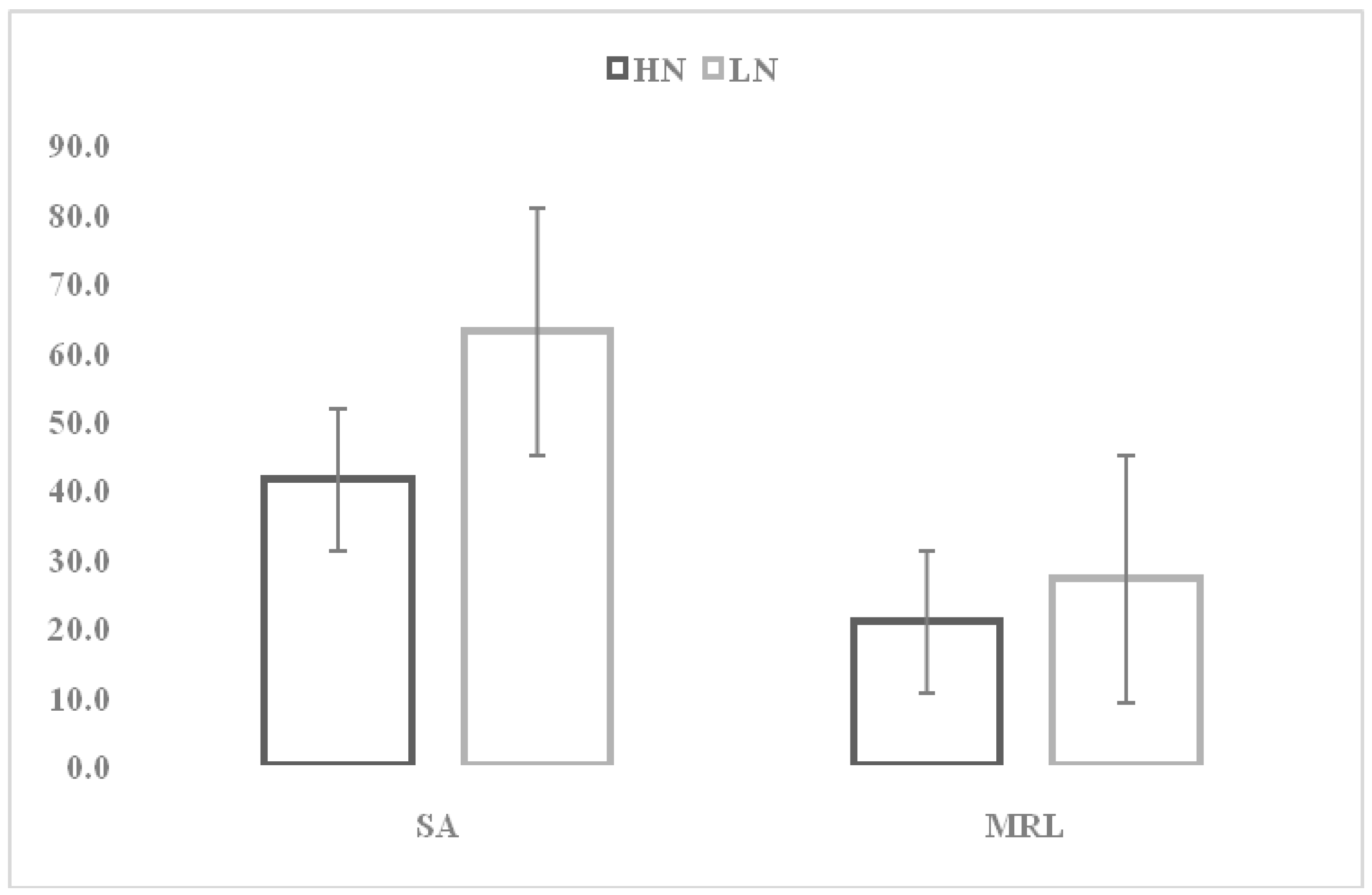
Mean Performance
Association Analysis
Path Analysis
4.2.2. Validation of Traits Contrivance for NUE in a Subset of Genotypes Was Grown for Eight Weeks
5. Discussion
6. Conclusions
- The germplasm under study shows the existence of sufficient genetic variability for the shoot and root traits relevant to NUE.
- A low nitrogen environment is more effective in discriminating the genotypes for root traits compared to a HN environment.
- RDW was found to be highly associated with better NUE under both nitrogen-rich and nitrogen-poor environments.
- Under N poor environment, the maximum root length can be directly selected to improve the NUE whereas N uptake can be improved by directly selecting the root biomass under an N-rich environment.
- Selection for root biomass is not in conflict with above-ground biomass, which is highly relevant for higher yield realization in the future.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.; Singh, S.S.; Jain, N.; Pratap Singh, G.; Prabhu, K.V. Wheat production in India: Technologies to face future challenges. J. Agric. Sci. 2010, 2, 164. [Google Scholar] [CrossRef] [Green Version]
- Marschner, P. Rhizosp here biology. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: Cambridge, MA, USA, 2012; pp. 369–388. [Google Scholar]
- Robertson, G.P.; Vitousek, P.M. Nitrogen in agriculture: Balancing the cost of an essential resource. Annu. Rev. Environ. Resour. 2009, 34, 97–125. [Google Scholar] [CrossRef] [Green Version]
- Campbell, C.A.; Zentner, R.P.; Selles, F.; McConkey, B.G.; Dyck, F.B. Nitrogen management for spring wheat grown annually onzero-tillage: Yield sand nitrogen use efficiency. Agron. J. 1993, 85, 107–114. [Google Scholar] [CrossRef]
- Good, A.G.; Beatty, P.H. Biotechnological approaches to improving nitrogen use efficiency in plants: A lanineamino transferaseasa case study. In The Molecular and Physiological Basis of Nutrient Use Efficiency in Crops; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011; pp. 165–191. [Google Scholar]
- Ranjan, R.; Yadav, R. Targeting nitrogen use efficiency for sustained production of cereal crops. J. Plant Nutr. 2019, 42, 1086–1113. [Google Scholar] [CrossRef]
- Malyan, S.K.; Bhatia, A.; Kumar, A.; Gupta, D.K.; Singh, R.; Kumar, S.S.; Tomer, R.; Kumar, O.; Jain, N. Methane production, oxidation and mitigation: A mechanistic understanding and comprehensive evaluation of influencing factors. Sci. Total Environ. 2016, 572, 874–896. [Google Scholar] [CrossRef] [PubMed]
- Raun, W.R.; Johnson, G.V. Improving nitrogen use efficiency for cereal production. Agron. J. 1999, 91, 357–363. [Google Scholar] [CrossRef] [Green Version]
- Andrews, M.; Lea, P.J.; Raven, J.A.; Lindsey, K. Can genetic manipulation of plant nitrogen assimilation enzymes result in increased crop yield and greater N-use efficiency? An assessment. Ann. Appl. Biol. 2004, 145, 25–40. [Google Scholar] [CrossRef]
- Foulkes, M.J.; Hawkesford, M.J.; Barraclough, P.B.; Holdsworth, M.J.; Kerr, S.; Kightley, S.; Shewry, P.R. Identifying traits to improve the nitrogen economy of wheat: Recent advances and future prospects. Field Crops Res. 2009, 114, 329–342. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Jamieson, P.D. Grain number, wheat yield, and bottling beer: An analysis. Field Crops Res. 2006, 98, 60–67. [Google Scholar] [CrossRef]
- Yadav, R.; Gupta, S.; Gaikwad, K.B.; Bainsla, N.K.; Kumar, M.; Babu, P.; Ansari, R.; Dhar, N.; Dharmateja, P.; Prasad, R. Genetic Gain in Yield and Associated Changes in Agronomic Traits in Wheat Cultivars Developed between 1900 and 2016 for Irrigated Ecosystems of North western Plain Zone of India. Front. Plant Sci. 2021, 12, 719394. [Google Scholar] [CrossRef] [PubMed]
- Ranjan, R.; Yadav, R.; Kumar, A.; Mandal, S.N. Contributing traits for nitrogen use efficiency in selected wheat genotypes and corollary between screening methodologies. Acta Agric. Scand. Sect. B-Soil Plant Sci. 2019, 69, 588–595. [Google Scholar] [CrossRef]
- Ranjan, R.; Yadav, R. Genetics analysis of nitrogen use efficiency component traits under nitrogen-limiting environment. Cereal Res. Commun. 2020, 48, 431–489. [Google Scholar] [CrossRef]
- Moll, R.H.; Kamprath, E.J.; Jackson, W.A. Analysis and interpretation off actors which contribute to efficiency of nitrogen utilization1. Agron. J. 1982, 74, 562–564. [Google Scholar] [CrossRef]
- Singh, R.K.; Choudhary, B.D. Biometrical Methods in Quantitative Genetics Analysis, 1st ed.; Kalyani Publishers: New Delhi, India, 1977. [Google Scholar]
- Ortiz-Monasterio, R.; Sayre, K.D.; Rajaram, S.; McMahon, M. Genetic progress in wheat yield and nitrogen use efficiency under four nitrogen rates. Crop Sci. 1997, 37, 898–904. [Google Scholar] [CrossRef]
- Foulkes, M.J.; Sylvester-Bradley, R.; Scott, R.K. Evidence for differences between winter wheat cultivars in acquisition of soil mineral nitrogen and uptake and utilization of applied fertilizer nitrogen. J. Agric. Sci. 1998, 130, 29–44. [Google Scholar] [CrossRef] [Green Version]
- Rakotoson, T.; Dusserre, J.; Letourmy, P.; Ramonta, I.R.; Cao, T.V.; Ramanantsoanirina, A.; Roumet, P.; Ahmadi, N.; Raboin, L.M. Genetic variability of nitrogen use efficiency in rain fed up land rice. Field Crops Res. 2017, 213, 194–203. [Google Scholar] [CrossRef]
- Nehe, A.S.; Misra, S.; Murchie, E.H.; Chinnathambi, K.; Foulkes, M.J. Genetic variation in N-use efficiency and associated traits in Indian wheat cultivars. Field Crops Res. 2018, 225, 152–162. [Google Scholar] [CrossRef]
- Almeida, V.C.; Viana, J.M.S.; De Oliveira, H.M.; Risso, L.A.; Ribeiro, A.F.S.; De Lima, R.O. Genetic diversity and path analysis for nitrogen use efficiency of tropical popcorn (Zea mays ssp. everta) in bred lines in adults tage. Plant Breed. 2018, 137, 839–847. [Google Scholar] [CrossRef]
- Ranjan, R.; Yadav, R.; Pandey, R.; Jain, N.; Bainsla, N.K.; Gaikwad, K.B.; Singh, A.M. Variation in wheat (Triticum aestvum) advance lines and released cultivars for traits associated with nitrogen use efficiency under N limiting environment. Indian J. Agric. Sci. 2019, 89, 99–104. [Google Scholar]
- Mălinaş, A.; Vidican, R.; Rotar, I.; Mălinaş, C.; Moldovan, C.M.; Proorocu, M. Current Status and Future Prospective for Nitrogen Use Efficiency in Wheat (Triticum aestivum L.). Plants 2022, 11, 217. [Google Scholar] [CrossRef] [PubMed]
- Decouard, B.; Bailly, M.; Rigault, M.; Marmagne, A.; Arkoun, M.; Soulay, F.; Caïus, J.; Le Roux, C.P.; Louahlia, S.; Jacquard, C.; et al. Genotypic Variation of Nitrogen Use Efficiency and Amino Acid Metabolism in Barley. Front. Plant Sci. 2022, 12, 807798. [Google Scholar] [CrossRef] [PubMed]
- Gaju, O.; Allard, V.; Martre, P.; Snape, J.W.; Heumez, E.; Le Gouis, J.; Moreau, D.; Bogard, M.; Griffiths, S.; Orford, S.; et al. Identification of traits to improve the nitrogen-use efficiency of wheat genotypes. Field Crops Res. 2011, 123, 139–152. [Google Scholar] [CrossRef]
- Petrarulo, M.; Marone, D.; Ferragonio, P.; Cattivelli, L.; Rubiales, D.; De Vita, P.; Mastrangelo, A.M. Genetic analysis of root morphological traits in wheat. Mol. Genet. Genom. 2015, 290, 785–806. [Google Scholar] [CrossRef]
- Ahmadi, J.; Pour-Aboughadareh, A.; Fabriki-Ourang, S.; Mehrabi, A.A.; Siddique, K.H. Screening wheat germplasm for seedling root architectural traits under contrasting water regimes: Potential sources of variability for drought adaptation. Arch. Agron. Soil Sci. 2018, 64, 1351–1365. [Google Scholar] [CrossRef]
- Yin, L.; Dai, X.; He, M. Delayed sowing improves nitrogen utilization efficiency in winter wheat without impacting yield. Field Crops Res. 2018, 221, 90–97. [Google Scholar] [CrossRef]
- Presterl, T.; Seitz, G.; Landbeck, M.; Thiemt, E.M.; Schmidt, W.; Geiger, H.H. Improving nitrogen use efficiency in European maize. Crop Sci. 2003, 43, 1259–1265. [Google Scholar] [CrossRef]
- Laperche, A.; Brancourt Hulmel, M.; Heumez, E.; Gardet, O.; Le Gouis, J. Estimation of genetic parameters of a DH wheat population grown at different N stress levels characterized by probe genotypes. Theor. Appl. Genet. 2006, 112, 797–807. [Google Scholar] [CrossRef]
- Sathisha, T.N.; Desai, S.A. Genetic Variability for Nitrogen Use Efficiency (NUE) and Yield Attributing Traitsin Wheat. Int. J. Agric. Sci. 2016, 8, 2010–2014. [Google Scholar]
- Naveen Kumar, V.M.; Uma, M.S. Genetic variability for nitrogen use efficiency and yield related traits in rice under low nitrogen and Available soil nitrogen conditions. Int. J. Appl. Pure Sci. Agric. 2016, 2, 83–90. [Google Scholar]
- Whalley, W.R.; Binley, A.; Watts, C.W.; Shanahan, P.; Dodd, I.C.; Ober, E.S.; Ashton, R.W.; Webster, C.P.; White, R.P.; Hawkesford, M.J. Methods to estimate changes in soil water for phenotyping root activity in the field. Plant Soil 2017, 415, 407–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, W.; Casao, M.C.; Wang, P.; Sato, K.; Hayes, P.M.; Finnegan, E.J.; Trevaskis, B. Direct links between the vernalization response and other key traits of cereal crops. Nat. Commun. 2015, 6, 5882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palta, J.A.; Chen, X.; Milroy, S.P.; Rebetzke, G.J.; Dreccer, M.F.; Watt, M. Large root systems: A re-they useful in adapting wheat to dry environments? Funct. Plant Biol. 2011, 38, 347–354. [Google Scholar] [CrossRef] [Green Version]
- Hawkesford, M.J. Reducing there lian ceon nitrogen fertilizer for wheat production. J. Cereal Sci. 2014, 59, 276–283. [Google Scholar] [CrossRef] [Green Version]
- Brancourt Hulmel, M.; Doussinault, G.; Lecomte, C.; Berard, P.; Le Buanec, B.; Trottet, M. Genetic improvement of agronomic traits of winter wheat cultivars released in France from 1946 to 1992. Crop Sci. 2003, 43, 37–45. [Google Scholar] [CrossRef]
- Muurinen, S.; Kleemola, J.; Peltonen-Sainio, P. Accumulation and translocation of nitrogen in spring cereal cultivars differing in nitrogen use efficiency. Agron. J. 2007, 99, 441–449. [Google Scholar] [CrossRef]
- Sadras, V.O.; Lawson, C. Nitrogen and water-use efficiency of Australian wheat varieties released between 1958 and 2007. Eur. J. Agron. 2013, 46, 34–41. [Google Scholar] [CrossRef]
- Bertin, P.; Gallais, A. Genetic variation for nitrogen use efficiency in a set of recombinant maize in bredlines. I. Agrophysiological results. Maydica 2000, 45, 53–66. [Google Scholar]
- Gallais, A.; Hirel, B. An approach to the genetics of nitrogen use efficiency in maize. J. Exp. Bot. 2004, 55, 295–306. [Google Scholar] [CrossRef] [Green Version]
- Sinebo, W.; Gretzmacher, R.; Edelbauer, A. Genotypic variation for nitrogen use efficiency in Ethiopian barley. Field Crops Res. 2004, 85, 43–60. [Google Scholar] [CrossRef]
- Srividhya, A.; Vemireddy, L.R.; Ramanarao, P.V.; Sridhar, S.; Jayaprada, M.; Anuradha, G.; Srilakshmi, B.; Reddy, H.K.; Hariprasad, A.S.; Siddiq, E.A. Molecular mapping of QTLs for drought related traits at seedling stage under PEG induced stress condition sin rice. Am. J. Plant Sci. 2011, 2, 190–201. [Google Scholar] [CrossRef] [Green Version]
- Hermans, C.; Hammond, J.P.; White, P.J.; Verbruggen, N. How do plants respond to nutrient shortage by biomass allocation? Trends Plant Sci. 2006, 11, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Hitz, K. Breeding for Nitrogen Use Efficiency in Soft Red Winter Wheat. Master’s Thesis, University of Kentucky, Lexington, KY, USA, 2015. [Google Scholar]
- Lynch, J.P. Roots of the second green revolution. Aust. J. Bot. 2007, 55, 493–512. [Google Scholar] [CrossRef]
- Caassen, N.; Barber, S.A. Simulation Modelfor Nutrient Up take from SoilbyaGrowingPlantRootSystem1. Agron. J. 1976, 68, 961–964. [Google Scholar] [CrossRef]
- Imada, S.; Yamanaka, N.; Tamai, S. Water table depth affects Populus alba fine root growth and whole plant biomass. Funct. Ecol. 2008, 22, 1018–1026. [Google Scholar] [CrossRef]
- McPhee, K. Variation for seedling root architecture in the core collection of pea germplasm. Crop Sci. 2005, 45, 1758–1763. [Google Scholar] [CrossRef]
- Kumar, B.; Abdel-Ghani, A.H.; Reyes-Matamoros, J.; Hochholdinger, F.; Lübberstedt, T. Genotypic variation for root architecture traits in seedlings of maize (Zea mays L.) inbred lines. Plant Breed. 2012, 131, 465–478. [Google Scholar] [CrossRef]
- Hurd, E.A. Root study of three wheat varieties and their resistance to drought and damage by soil cracking. Can. J. Plant Sci. 1964, 44, 240–248. [Google Scholar] [CrossRef]
- Price, A.H.; Townend, J.; Jones, M.P.; Audebert, A.; Courtois, B. Mapping QTLs associated with drought avoidance in upland rice grown in the Philippine sand West Africa. Plant Mol. Biol. 2002, 48, 683–695. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition | SDW | RDW | TDW | R:S | MRL | N% | gN | NUpE | NUtE | NUE |
|---|---|---|---|---|---|---|---|---|---|---|
| HN | 0.200 ± 0.04 | 0.040 ± 0.003 | 0.232 ± 0.04 | 0.012 ± 0.04 | 400.2 ± 0.742 | 0.358 ± 0.14 | 0.000 ± 0.001 | 0.313 ± 0.06 | 95.2 ± 2.24 | 384.2 ± 1.839 |
| LN | 0.049 ± 0.01 | 0.002 ± 0.0055 | 0.071 ± 0.01 | 0.001 ± 0.01 | 1984.04 ± 1.82 | 0.243 ± 0.11 | 0.000 ± 0.14 | 0.385 ± 0.09 | 711.5 ± 6.62 | 1519.8 ± 2.22 |
| GXE | 0.100 ± 0.03 | 0.003 ± 0.004 | 0.123 ± 0.03 | 0.006 ± 0.03 | 1152 ± 1.392 | 0.328 ± 0.13 | 0.000 ± 0.00 | 0.013 ± 0.08 | 428 ± 4.945 | 761.6 ± 2.040 |
| p-Value | ||||||||||
| HN | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| LN | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| GXE | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| Traits | Mean | Range | Hbs | GA@1% | ||||
|---|---|---|---|---|---|---|---|---|
| HN | LN | HN | LN | HN | LN | HN | LN | |
| SDW | 0.617 | 0.333 | 0.195–1.889 | 0.08–0.78 | 0.96 | 0.985 | 0.66 | 0.335 |
| RDW | 0.064 | 0.066 | 0.016–0.246 | 0.02–0.16 | 0.98 | 0.947 | 0.095 | 0.067 |
| TDW | 0.681 | 0.400 | 0.221–2.135 | 0.1–0.93 | 0.96 | 0.988 | 0.717 | 0.404 |
| R:S | 0.109 | 0.199 | 0.026–0.327 | 0.15–0.27 | 0.45 | 0.218 | 0.095 | 0.015 |
| MRL | 32.05 | 90.40 | 17.2–72.0 | 42.5–162 | 0.99 | 0.992 | 30.3 | 67.5 |
| N% | 2.59 | 1.45 | 1.552–4.078 | 0.86–2.41 | 0.77 | 0.776 | 0.766 | 0.632 |
| gN | 0.016 | 0.005 | 0.062–0.004 | 0.007–0.015 | 0.93 | 0.89 | 0.019 | 0.005 |
| NUpE | 0.698 | 0.848 | 0.178–2.704 | 0.123–2.71 | 0.94 | 0.906 | 0.819 | 0.885 |
| NUtE | 39.3 | 72.0 | 24.58–64.56 | 41.6–117.5 | 0.79 | 0.766 | 12.7 | 33.8 |
| NUE | 26.8 | 58.4 | 8.44–82.13 | 14.04–136 | 0.96 | 0.985 | 28.8 | 58.8 |
| Trait | Mean (SE) | Range | ||
|---|---|---|---|---|
| HN | LN | HN | LN | |
| RSA (cm2) | 41.45 (1.98) | 62.91 (3.56) | 20–64 | 30–152.8 |
| RV (cm3) | 0.329 (0.185) | 0.637 (0.685) | 0.145–0.55 | 0.246–2.814 |
| AD (mm) | 0.326 (0.073) | 0.289 (0.068) | 0.277–0.474 | 0.24–0.381 |
| N Tips | 1606.9 (17.5) | 1592.5 (10.8) | 520–2898 | 972.6–2480.6 |
| MRL (cm) | 20.89 (0.53) | 27 (1.07) | 16.5–25.5 | 19.2–43.6 |
| TRL (cm) | 424.9 (6.33) | 514.5 (4.38) | 166–630 | 324.4–690.4 |
| SDW (g) | 0.071 (0.093) | 0.047 (0.048) | 0.034–0.137 | 0.032–0.07 |
| RDW (g) | 0.017 (0.039) | 0.015 (0.038) | 0.01–0.033 | 0.005–0.023 |
| RSA | RV | N Tips | TRL | SDW | RDW | |
|---|---|---|---|---|---|---|
| RSA | 0.642 | −1.096 | −0.276 | 1.586 | −0.146 | 0.709 |
| RV | 0.604 | −1.166 | −0.224 | 1.479 | −0.159 | 0.534 |
| N tips | 0.305 | −0.447 | −0.583 | 1.107 | −0.044 | 0.337 |
| TRL | 0.592 | −1.001 | −0.375 | 1.722 | −0.172 | 0.766 |
| SDW | 0.366 | −0.723 | −0.100 | 1.152 | −0.257 | 0.438 |
| RSA | RV | AD | RL | TRL | SDW | RDW | |
|---|---|---|---|---|---|---|---|
| RSA | −0.119 | 0.072 | 0.032 | 0.001 | 0.383 | 0.223 | 0.592 |
| RV | −0.104 | 0.083 | 0.027 | 0.002 | 0.299 | 0.225 | 0.530 |
| AD | −0.098 | 0.057 | 0.039 | 0.002 | 0.267 | 0.210 | 0.477 |
| MRL | −0.034 | 0.018 | 0.003 | 0.008 | 0.382 | 0.154 | 0.531 |
| TRL | −0.089 | 0.048 | 0.020 | 0.004 | 0.515 | 0.220 | 0.718 |
| SDW | −0.056 | 0.039 | 0.017 | 0.000 | 0.240 | 0.472 | 0.711 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ranjan, R.; Yadav, R.; Gaikwad, K.B.; Bainsla, N.K.; Kumar, M.; Babu, P.; Dharmateja, P. Spring Wheat’s Ability to Utilize Nitrogen More Effectively Is Influenced by Root Phene Variation. Plants 2023, 12, 1010. https://doi.org/10.3390/plants12051010
Ranjan R, Yadav R, Gaikwad KB, Bainsla NK, Kumar M, Babu P, Dharmateja P. Spring Wheat’s Ability to Utilize Nitrogen More Effectively Is Influenced by Root Phene Variation. Plants. 2023; 12(5):1010. https://doi.org/10.3390/plants12051010
Chicago/Turabian StyleRanjan, Rumesh, Rajbir Yadav, Kiran B. Gaikwad, Naresh Kumar Bainsla, Manjeet Kumar, Prashanth Babu, and Palaparthi Dharmateja. 2023. "Spring Wheat’s Ability to Utilize Nitrogen More Effectively Is Influenced by Root Phene Variation" Plants 12, no. 5: 1010. https://doi.org/10.3390/plants12051010