
The Study on Sea Buckthorn (Genus Hippophae L.) Fruit Reveals Cell Division and Cell Expansion to Promote Morphogenesis
Abstract
:1. Introduction
2. Results
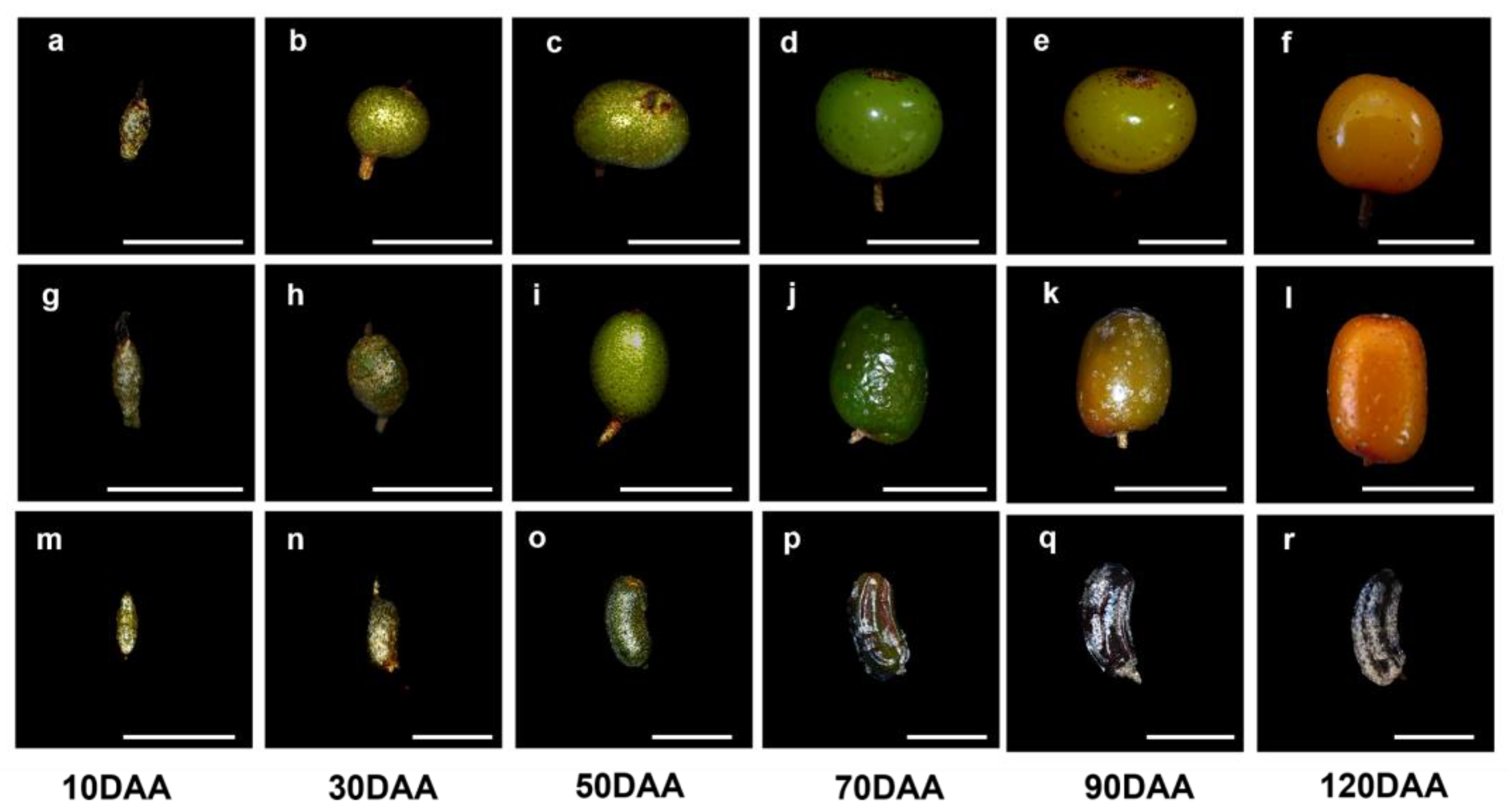
2.1. General Observation
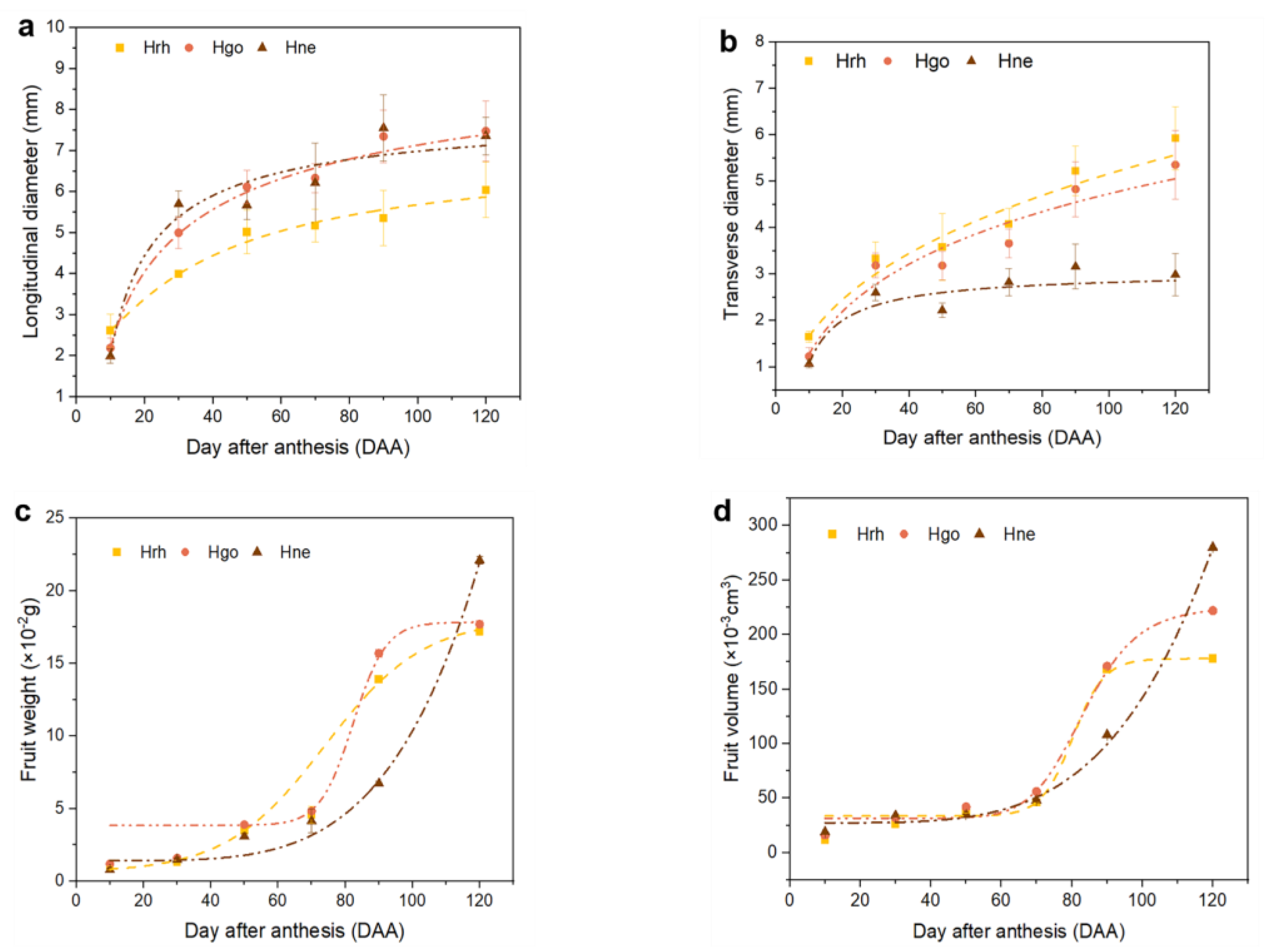
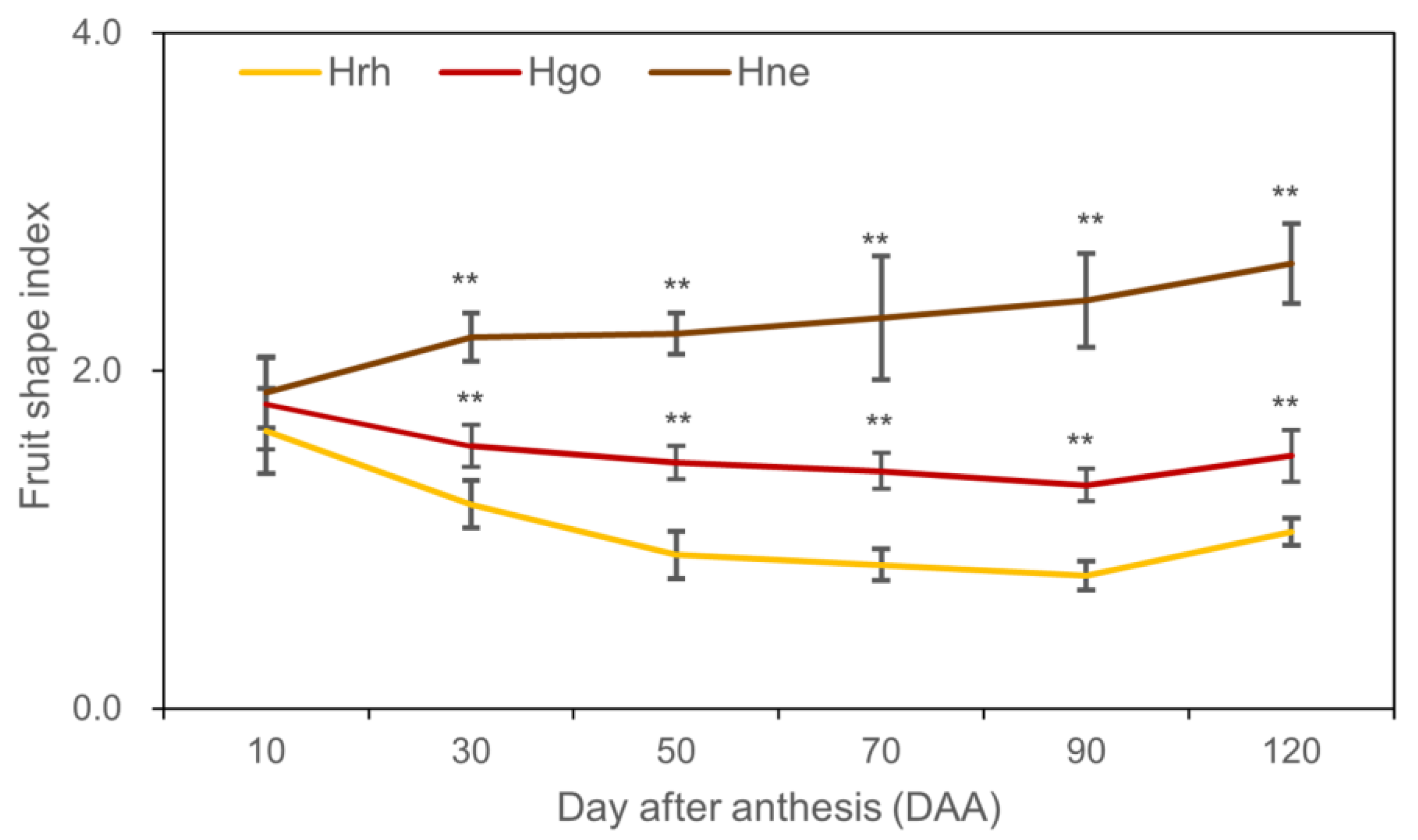
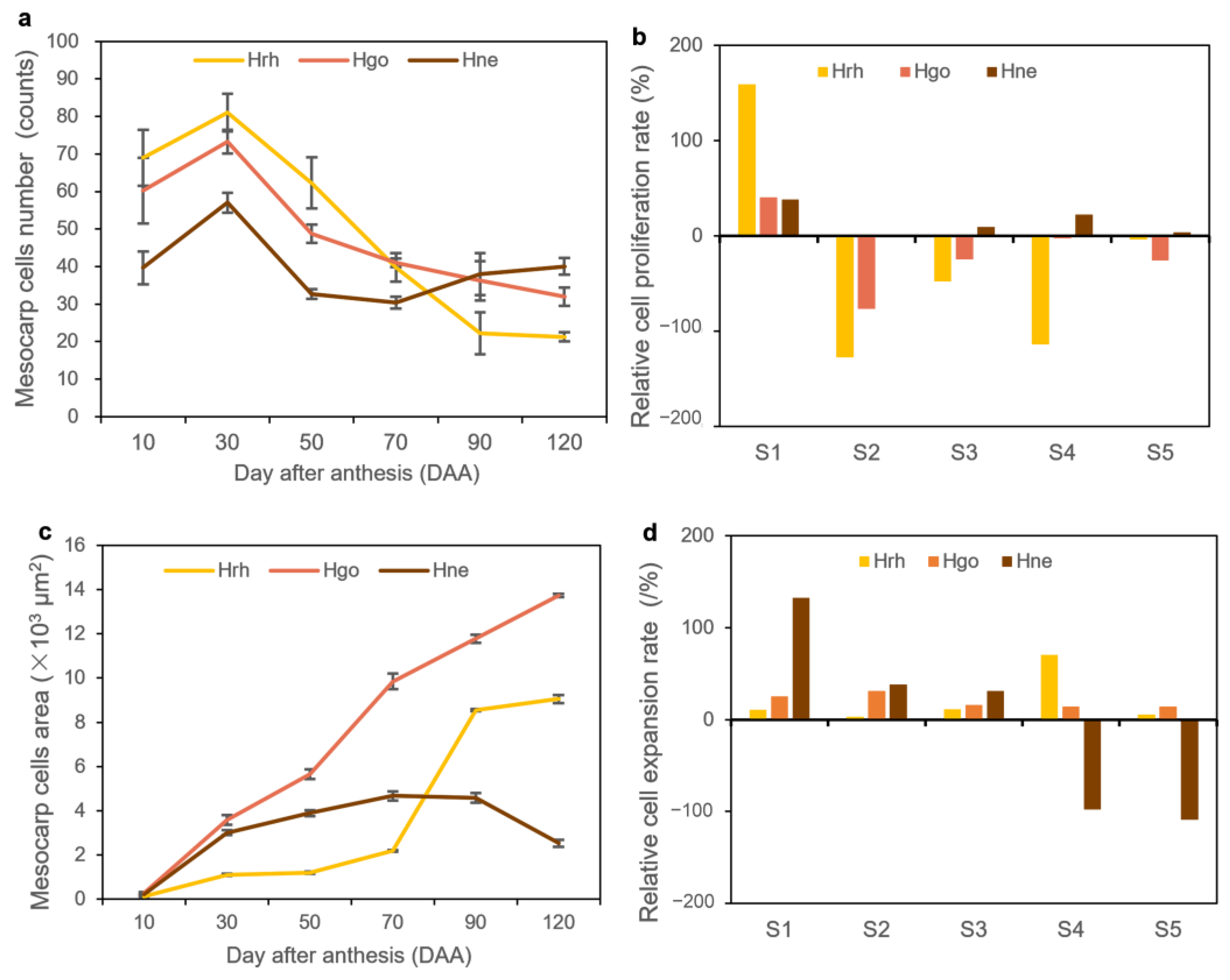
2.2. Fruit Growth Pattern
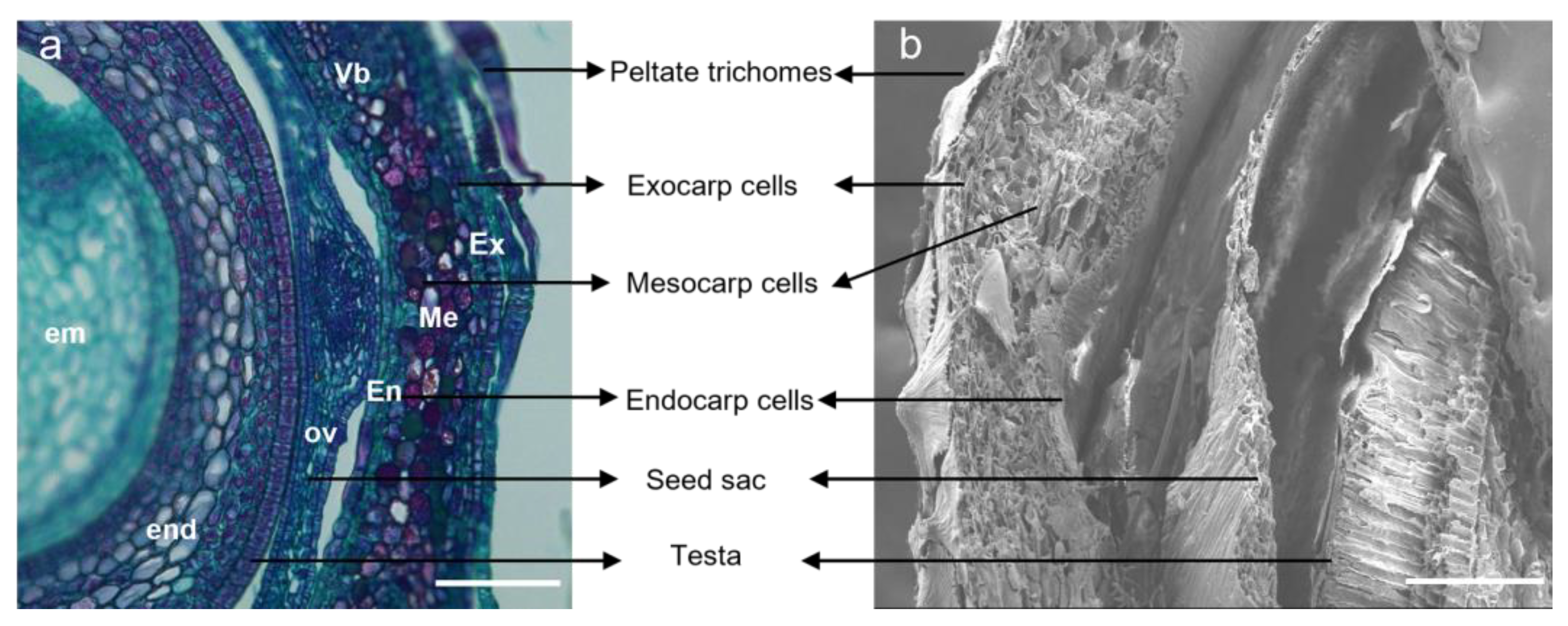
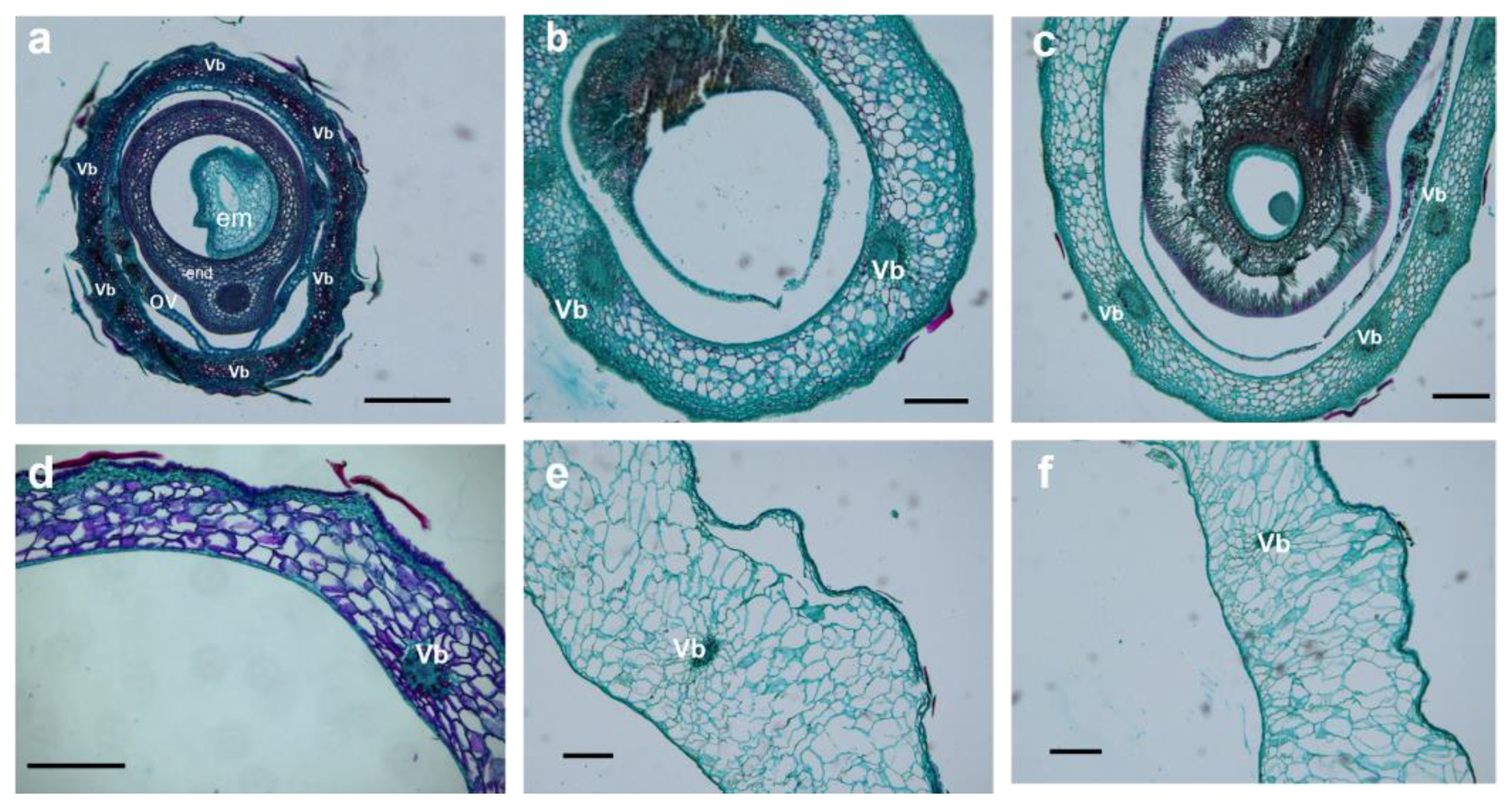
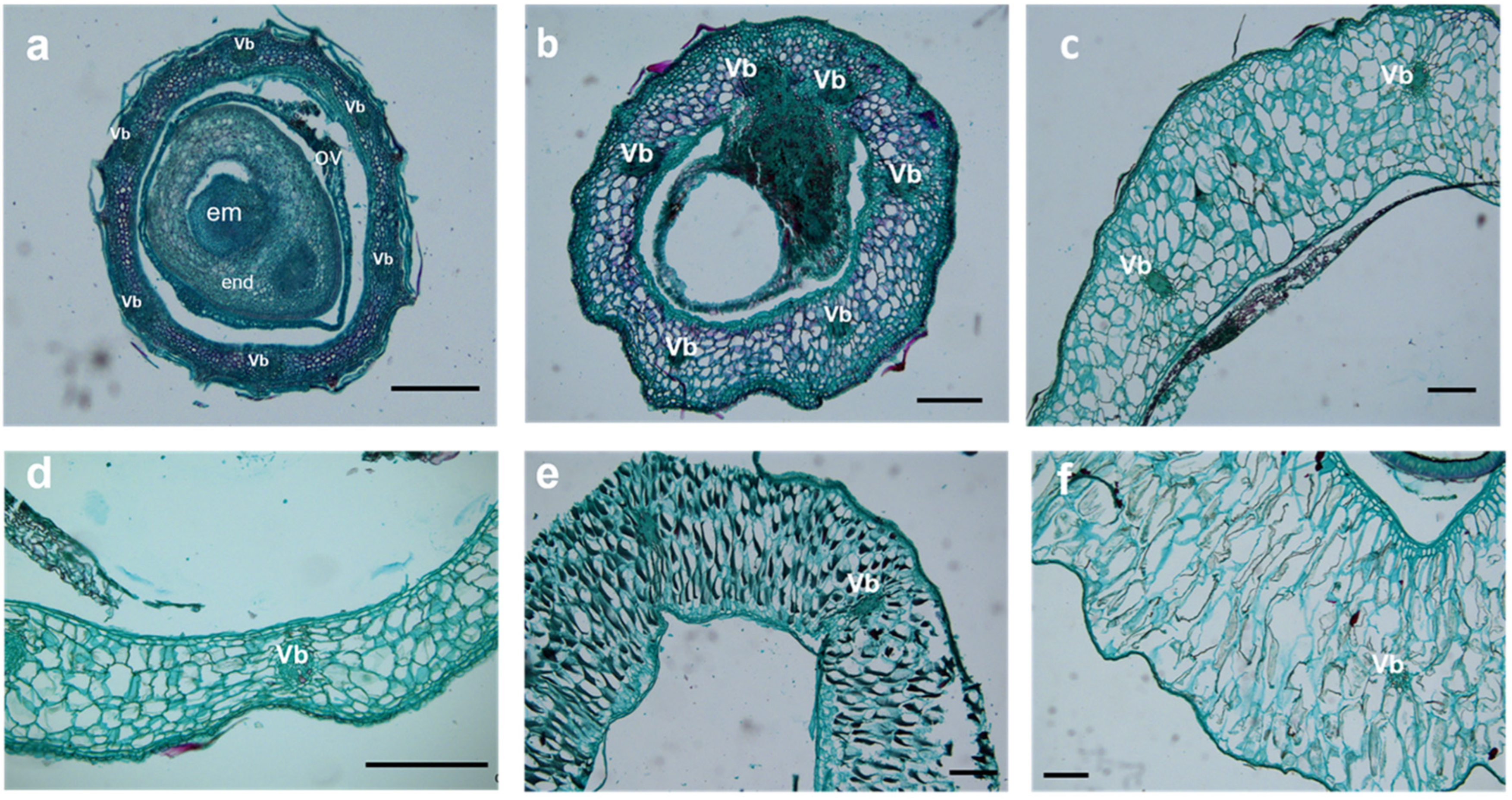
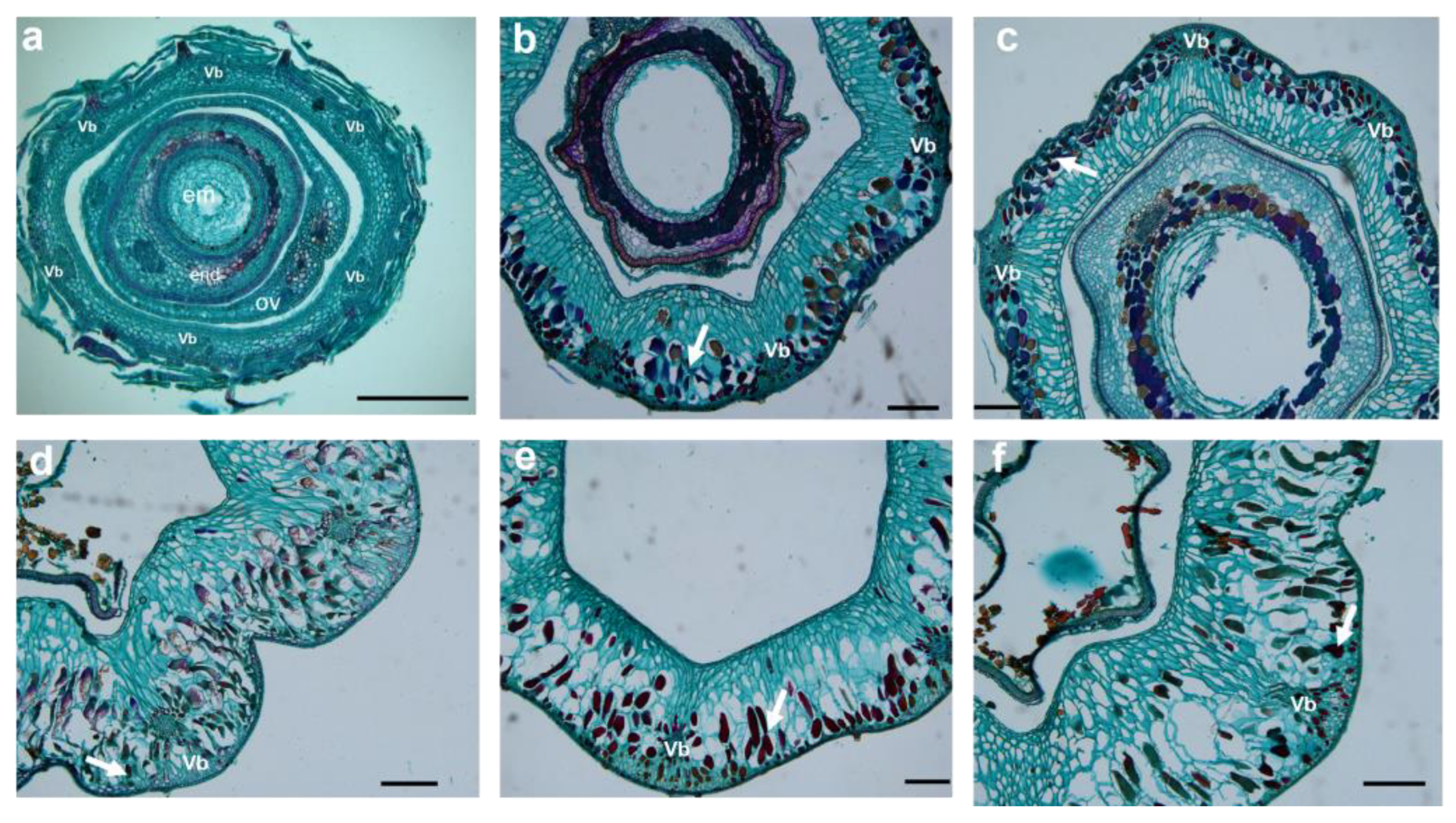
2.3. Cellular Structure Changes
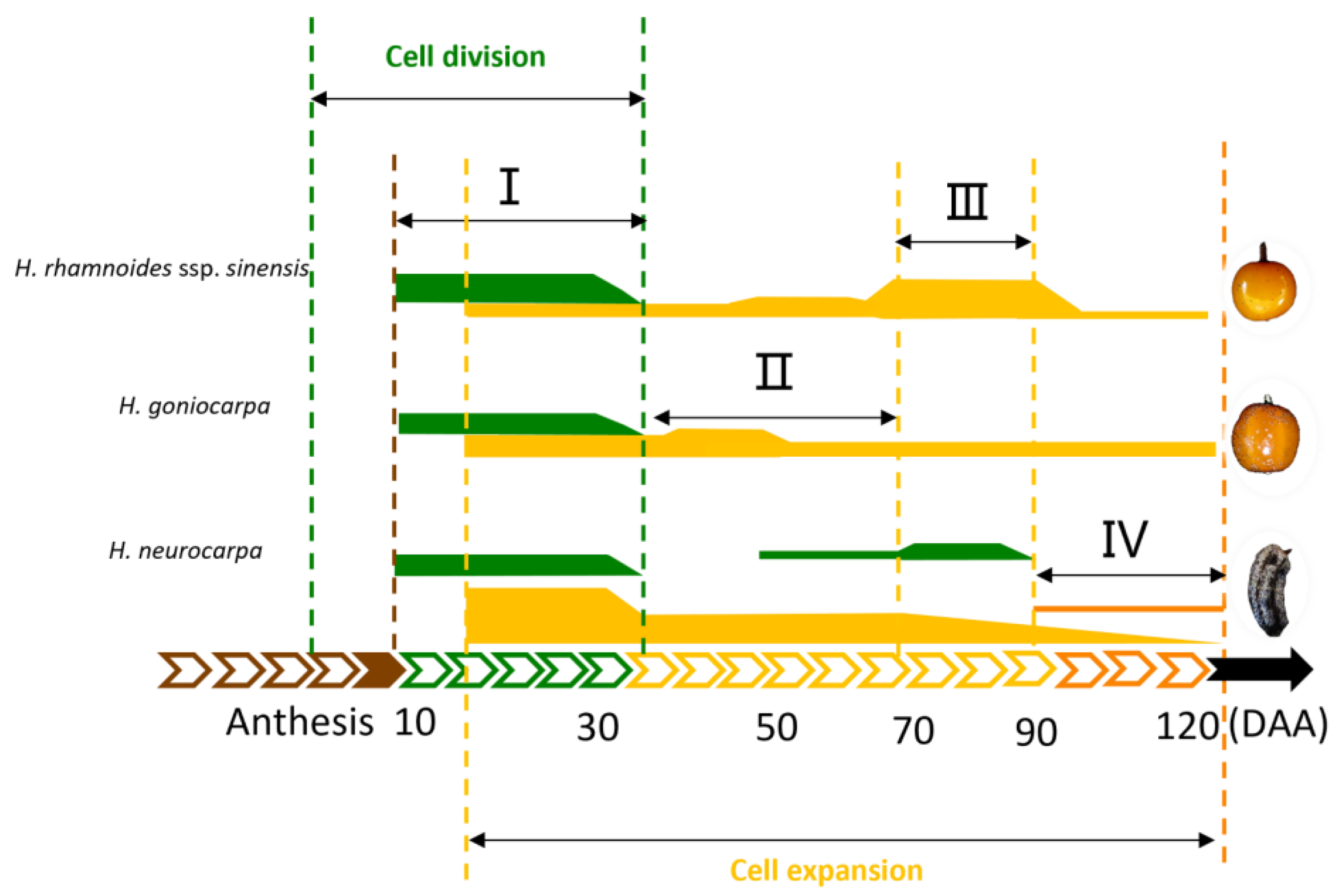
3. Discussion
3.1. Fruit Growth and Development in Hippophae L.
3.2. Effects of Cell Division and Cell Extension on the Fruit Size of Hippophae L.
3.3. A Model of Cell Regulation in Fruit Development of Hippophae L.
4. Materials and Methods
4.1. Plant Materials
4.2. Methods
4.2.1. Fruit Characteristics of Sea Buckthorn during Development
4.2.2. The Microstructure of Fruits at Different Development Stages
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Andersson, S.C.; Olsson, M.E.; Johansson, E.; Rumpunen, K. Carotenoids in sea buckthorn (Hippophae L.) berries during ripening and use of pheophytin a as a maturity marker. J. Agric. Food Chem. 2009, 57, 250–258. [Google Scholar] [CrossRef]
- Bartish, I.V.; Jeppsson, N.; Nybom, H.; Swenson, U. Phylogeny of Hippophae (Elaeagnaceae) inferred from parsimony analysis of chloroplast DNA and morphology. Syst. Bot. 2002, 27, 41–54. [Google Scholar] [CrossRef]
- Manickam, M.; Bhardwaj, A.; Rahamathulla, S.; Duraisamy, A.J. Hippophae sp.: A Boon for High-Altitude Maladies. In Management of High-Altitude Pathophysiology; Academic Press: Cambridge, MA, USA, 2018; pp. 29–68. [Google Scholar] [CrossRef]
- Rousi, A. The genus Hippophae L. A taxonomic study. Ann. Bot. Fenn. 1971, 8, 177–227. [Google Scholar]
- Husain, M.; Rathore, J.P.; Rasool, A.; Parrey, A.A.; Vishwakarma, D.K.; Mahendar, K. Seabuckthorn: A multipurpose shrubs species in Ladakh cold desert. J. Entomol. 2018, 6, 1330–1337. [Google Scholar]
- Mangla, Y.; Chaudhary, M.; Gupta, H.; Thakur, R.; Goel, S.; Raina, S.N.; Tandon, R. Facultative apomixis and development of fruit in a deciduous shrub with medicinal and nutritional uses. AoB Plants 2015, 7, 98. [Google Scholar] [CrossRef]
- Lian, Y.S.; Chen, X.L.; Ma, R.J.; Li, C.B. The primary study of biological and ecological property for the genus Hippophae L. Hippophae 1998, 11, 1–7. [Google Scholar]
- Lian, Y.S.; Chen, X.L.; Sun, K.; Ma, R.J. Clarification of the systematic position of Hippophae goniocarpa (Elaeagnaceae). Bot. J. Linn. Soc. 2003, 142, 425–430. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Kumar, P.; Sharma, A.; Sharma, D.P.; Thakur, M. Scientific insights to existing know-how, breeding, genetics, and biotechnological interventions pave the way for the adoption of high-value underutilized super fruit Sea buckthorn (Hippophae L.). S. Afr. J. Bot. 2021, 145, 348–359. [Google Scholar] [CrossRef]
- Ciesarová, Z.; Murkovic, M.; Cejpek, K.; Kreps, F.; Tobolková, B.; Koplík, R.; Belajová, E.; Kukurová, K.; Daško, L.; Panovská, Z.; et al. Why is sea buckthorn (Hippophae L.) so exceptional? A review. Food Res. Int. 2020, 133, 109170. [Google Scholar] [CrossRef]
- Olas, B. The beneficial health aspects of sea buckthorn (Elaeagnus rhamnoides L. A. Nelson) oil. J. Ethnopharmacol. 2017, 213, 183–190. [Google Scholar] [CrossRef]
- Yu, L.; Diao, S.; Zhang, G.; Yu, J.; Zhang, T.; Luo, H.; Duan, A.; Wang, J.; He, C.; Zhang, J. Genome sequence and population genomics provide insights into chromosomal evolution and phytochemical innovation of Hippophae rhamnoides. Plant Biotechnol. Rep. 2022, 20, 1257–1273. [Google Scholar] [CrossRef]
- Lee, Y.H.; Jang, H.J.; Park, K.H.; Kim, S.H.; Kim, J.K.; Kim, J.C.; Jang, T.S.; Kim, K.H. Phytochemical analysis of the fruits of sea buckthorn (Hippophaë rhamnoides): Identification of organic acid derivatives. Plants 2021, 10, 860. [Google Scholar] [CrossRef]
- Dupak, R.; Hrnkova, J.; Simonova, N.; Kovac, J.; Ivanisova, E.; Kalafova, A.; Schneidgenova, M.; Prnova, M.S.; Brindza, J.; Tokarova, K.; et al. The consumption of sea buckthorn (Hippophae L.) effectively alleviates type 2 diabetes symptoms in spontaneous diabetic rats. Res. Vet. Sci. 2022, 152, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Guan, Y.; Sang, Z.; Dong, J.; Wei, R. Isolation and characterization of auronlignan derivatives with hepatoprotective and hypolipidemic activities from the fruits of Hippophae L. Food Funct. 2022, 13, 7750–7761. [Google Scholar] [CrossRef] [PubMed]
- Crisosto, C.H.; Costa, G. Preharvest factors affecting peach quality. In The Peach: Botany Production and Uses; Layne, D.R., Bassi, D., Eds.; CABI: Cambridge, UK, 2008; pp. 536–549. [Google Scholar]
- NeSmith, D.S. Blueberry cultivar development at the University of Georgia. Acta Hortic. 2009, 810, 79–86. [Google Scholar] [CrossRef]
- Bain, J.M.; Robertson, R.N. The physiology of growth in apple fruits. I. Cell size, cell number, and fruit development. Aust. J. Sci. Res. 1951, 4, 75–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corelli-Grappadelli, L.; Lasko, A.N. Fruit development in deciduous tree crops as affected by physiological factors and environmental conditions. Acta Hortic. 2004, 636, 425–441. [Google Scholar] [CrossRef]
- Blaker, K.M.; Olmstead, J.W. Stone cell frequency and cell area variation of crisp and standard texture southern highbush blueberry fruit. J. Am. Soc. Hortic. Sci. 2014, 139, 553–557. [Google Scholar] [CrossRef] [Green Version]
- Forney, C.F.; Kalt, W.; Jordan, M.A.; Vinqvist-Tymchuk, M.R.; Fillmore, S.A. Blueberry and cranberry fruit composition during development. J. Berry Res. 2012, 2, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Takeo, H.; Wakako, K.; Masumi, Y.; Yuhya, W.; Takashi, S. Involvent of cell proliferation and cell enlargem in increasing the fruit size of Malus species. Sci. Hortic. 2005, 8, 447–456. [Google Scholar] [CrossRef]
- Bartolini, S.; Vitagliano, C.; Andreucci, A.C. Anatomical and biochemical studies in persisting and shedding seeds during the development of peach fruits. Plant Biosyst. 2001, 135, 175–181. [Google Scholar] [CrossRef]
- Joubes, J.; Phan, T.H.; Just, D.; Rothan, C.; Bergounioux, C.; Raymond, P.; Chevalier, C. Molecular and biochemical characterization of the involvement of cyclin-dependent kinase A during the early development of tomato fruit. Plant Physiol. 1999, 121, 857–869. [Google Scholar] [CrossRef] [Green Version]
- Malladi, A.; Hirst, P.M. Increase in fruit size of a spontaneous mutant of ‘Gala’ apple (Malus × domestica Borkh.) is facilitated by altered cell production and enhanced cell size. J. Exp. Bot. 2010, 61, 3003–3013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, W.; Zhu, Y.; Zhang, L.; Yang, X.; Gao, Y.; Lin, S. The cellular physiology of loquat (Eriobotrya japonica Lindl.) fruit with a focus on how cell division and cell expansion processes contribute to pome morphogenesis. Sci. Hortic. 2017, 224, 142–149. [Google Scholar] [CrossRef]
- Schroeder, C.A. Growth and development of the Fuerte avocado fruit. In Proceedings of the American Society for Horticultural Science. J. Am. Soc. Hortic. Sci. 1953, 61, 103–109. [Google Scholar]
- Hu, D.L.; Richards, P.; Alexeev, A. The growth of giant pumpkins: How extreme weight influences shape. Int. Non-Linear Mech. 2011, 46, 637–647. [Google Scholar] [CrossRef]
- Nakata, Y.; Taniguchi, G.; Takazaki, S.; Ode-Uda, N.; Miyahara, K.; Ohshima, Y. Comparative analysis of cells and proteins of pumpkin plants for the control of fruit size. J. Biosci. Bioeng. 2012, 114, 334–341. [Google Scholar] [CrossRef]
- Johnson, L.K.; Malladi, A.; Scott NeSmith, D. Differences in cell number facilitate fruit size variation in rabbiteye blueberry genotypes. J. Am. Soc. Hortic. Sci. 2011, 136, 10–15. [Google Scholar] [CrossRef] [Green Version]
- Bartish, I.V.; Jeppsson, N.; Bartish, G.I. Inter-and intraspecific genetic variation in Hippophae (Elaeagnaceae) investigated by RAPD markers. Plant Syst. Evol. 2002, 225, 85–101. [Google Scholar] [CrossRef]
- Cowan, R.S. Review: [Untitled]: Vascular Plant Systematics. Taxon 1975, 24, 383–384. [Google Scholar] [CrossRef]
- Bewley, J.D.; Black, M.; Halmer, P. The encyclopedia of seeds: Science, technology and uses. Cabi 2006, 23, 137–143. [Google Scholar] [CrossRef]
- Harrison, J.E.; Beveridge, T. Fruit structure of Hippophae rhamnoides cv. Indian Summer (sea buckthorn). Can. J. Bot. 2002, 80, 399–409. [Google Scholar] [CrossRef]
- Hrabovský, M.; Randáková, Z.; Rendeková, A.; Mičieta, K. Classification of fruits of vascular plants–news in morphology and Slovak nomenclature. Acta Bot. Univ. Comen. 2017, 52, 71–83. Available online: https://www.researchgate.net/publication/327107576 (accessed on 20 February 2023).
- Ortiz, J.P.; Quarin, C.L.; Pessino, S.C.; Acuña, C.; Martínez, E.J.; Espinoza, F.; Hojsgaard, D.H.; Sartor, M.E.; Pupilli, F. Harnessing apomictic reproduction in grasses: What we have learned from Paspalum. Ann. Bot. 2013, 112, 767–787. [Google Scholar] [CrossRef] [Green Version]
- Higgins, J.; Tomaszewska, P.; Pellny, T.K.; Castiblanco, V.; Arango, J.; Tohme, J.; Schwarzacher, T.; Mitchell, R.A.; Heslop-Harrison, J.S.; De Vega, J. Diverged subpopulations in tropical Urochloa (Brachiaria) forage species indicate a role for facultative apomixis and varying ploidy in their population structure and evolution. Ann. Bot. 2022, 130, 657–669. [Google Scholar] [CrossRef]
- Cuevas, J.; Salvador-Sola, F.J.; Gavilan, J.; Lorente, N.; Hueso, J.J.; Gonzalez-Padierna, C.M. Loquat fruit sink strength and growth pattern. Sci. Hortic. 2003, 98, 131–137. [Google Scholar] [CrossRef]
- Hernández, M.S.; Martínez, O.; Fernández-Trujillo, J.P. Behavior of arazá (Eugenia stipitata Mc Vaugh) fruit quality traits during growth, development and ripening. Sci. Hortic. 2007, 111, 220–227. [Google Scholar] [CrossRef]
- Tee, Y.K.; Ding, P.; Rahman, N.A.A. Physical and cellular structure changes of Rastali banana (Musa AAB) during growth and development. Sci. Hortic. 2011, 129, 382–389. [Google Scholar] [CrossRef] [Green Version]
- Gi Mohammad, M.; Ding, P. Physico-textural and cellular structure changes of Carissa congesta fruit during growth and development. Sci. Hortic. 2019, 246, 380–389. [Google Scholar] [CrossRef]
- Rančić, D.; Quarrie, S.P.; Pećinar, I. Anatomy of tomato fruit and fruit pedicel during fruit development. Microsc. Sci. Technol. Appl. Educ. 2010, 2, 851–861. [Google Scholar]
- Gllaspy, G.; Ben-David, H.; Gruissem, W. Fruits: A developmental perspective. Plant Cell 1993, 5, 1439–1451. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Tanabe, K.; Wang, S.; Tamura, F.; Yoshida, A.; Matsumoto, K. The impact of cell division and cell enlargement on the evolution of fruit size in Pyrus pyrifolia. Ann. Bot. 2006, 98, 537–543. [Google Scholar] [CrossRef]
- Ding, C.; Zhang, H. The effect of plant hormones on fruit development of loquat. Acta Hortic. Sin. 1988, 15, 148–154. [Google Scholar]
- Harada, T.; Kurahashi, W.; Yanai, M.; Wakasa, Y.; Satoh, T. Involvement of cell proliferation and cell enlargement in increasing the fruit size of Malus species. Sci. Hortic. 2005, 105, 447–456. [Google Scholar] [CrossRef]
- Olmstead, J.W.; Iezzoni, A.F.; Whiting, M.D. Genotypic differences in sweet cherry fruit size are primarily a function of cell number. J. Am. Soc. Hortic. Sci. 2017, 132, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Jullien, A.; Munier-Jolain, N.G.; Malezieux, E.; Chillet, M.; Ney, B. Effect of pulp cell number and assimilate availabity on dry matter accumulation rate in a banana fruit [Musa sp. AAA group ‘Grande Naine’ (Cavendish subgroup)]. Ann. Bot. 2011, 88, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Guan, L.; Fan, X.C.; Zheng, T.; Dong, T.Y.; Liu, C.H.; Fang, J.G. Anatomical characteristics associated with different degrees of berry cracking in grapes. Sci. Hortic. 2020, 261, 108992. [Google Scholar] [CrossRef]
- Zhang, C.; Fan, X.; Liu, C.; Fang, J. Anatomical berry characteristics during the development of grape berries with different shapes. Hortic. Plant J. 2021, 7, 295–306. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Dependent Variable | Logistic Models | R2 |
|---|---|---|---|
| H. rhamnoides ssp. sinensis | longitudinal diameter (mm) | 0.95192 | |
| transverse diameter (mm) | 0.97472 | ||
| fruit weight (×10−2 g) | 0.99241 | ||
| fruit volume (×10−3 cm3) | 0.99669 | ||
| H. goniocarpa | longitudinal diameter (mm) | 0.90595 | |
| transverse diameter (mm) | 0.94341 | ||
| fruit weight (×10−2 g) | 0.90155 | ||
| fruit volume (×10−3 cm3) | 0.99714 | ||
| H. neuvocarpa | longitudinal diameter (mm) | 0.94016 | |
| transverse diameter (mm) | 0.90831 | ||
| fruit weight (×10−2 g) | 0.98673 | ||
| fruit volume (×10−3 cm3) | 0.99948 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Zhang, Z.; Zhou, H.; Bai, Z.; Sun, K. The Study on Sea Buckthorn (Genus Hippophae L.) Fruit Reveals Cell Division and Cell Expansion to Promote Morphogenesis. Plants 2023, 12, 1005. https://doi.org/10.3390/plants12051005
Zhao J, Zhang Z, Zhou H, Bai Z, Sun K. The Study on Sea Buckthorn (Genus Hippophae L.) Fruit Reveals Cell Division and Cell Expansion to Promote Morphogenesis. Plants. 2023; 12(5):1005. https://doi.org/10.3390/plants12051005
Chicago/Turabian StyleZhao, Jing, Zhihua Zhang, Hongdan Zhou, Zengfu Bai, and Kun Sun. 2023. "The Study on Sea Buckthorn (Genus Hippophae L.) Fruit Reveals Cell Division and Cell Expansion to Promote Morphogenesis" Plants 12, no. 5: 1005. https://doi.org/10.3390/plants12051005