
The Genotypic Variability among Short-Season Soybean Cultivars for Nitrogen Fixation under Drought Stress
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Germplasm, Plant Materials, and Growth Conditions
2.2. Field Capacity of Growth Media
2.3. Drought Treatment and Yield Data Collection
2.4. Determination of Nitrogen Fixation-Related Parameters
2.5. Statistical Analysis
2.6. Genotyping Data
2.7. Population Structure
2.8. Genome-Wide Association Study
2.9. Candidate Gene Identification
3. Results
3.1. Phenotypic Variation of Yield- and Nitrogen Fixation-Related Traits in Soybean
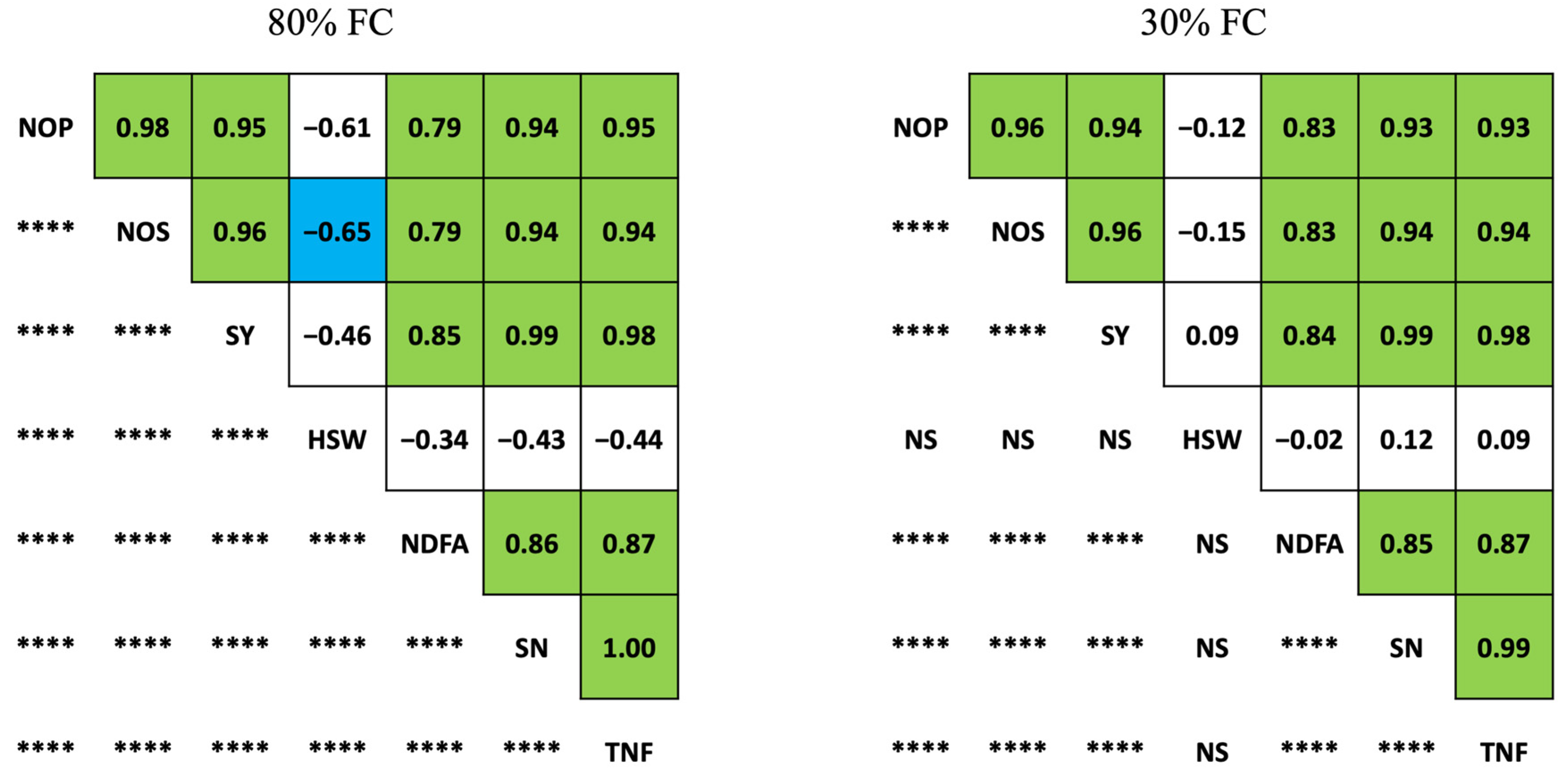
3.2. Correlations among Yield- and Nitrogen Fixation-Related Traits in Soybean
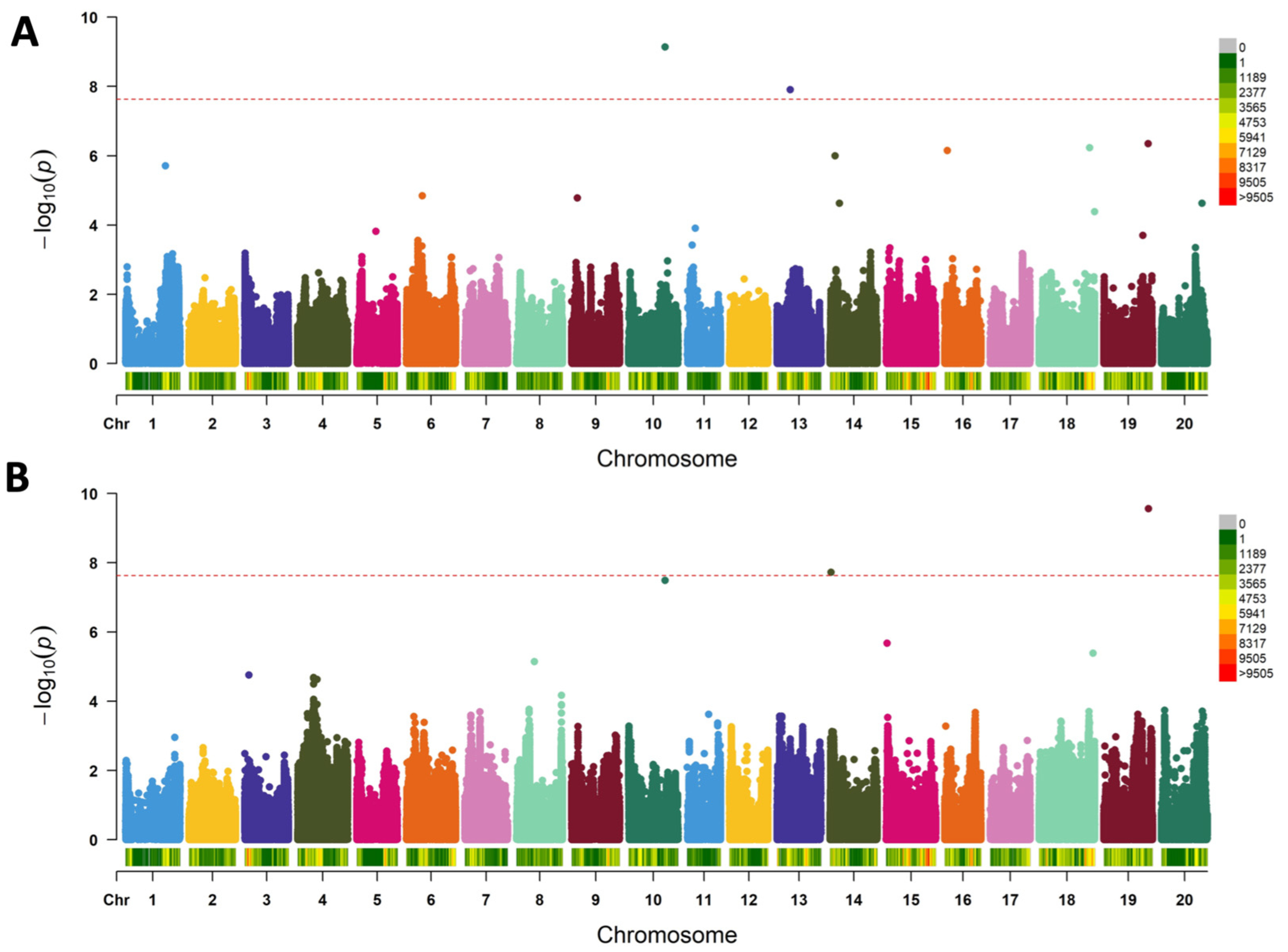
3.3. Genome-Wide Association of Yield- and Nitrogen Fixation-Related Traits
3.4. Yield- and Nitrogen Fixation-Trait-Related Candidate Genes
4. Discussion
4.1. Significant Phenotypic Variation of Yield- and Nitrogen Fixation-Related Traits in Soybean
4.2. Drought Stress on Symbiotic Nitrogen Fixation
4.3. Correlations among Yield- and Nitrogen Fixation-Related Traits
4.4. Genome-Wide Association Using Whole-Genome Data Revealed Five QTLs Controlling %Ndfa
4.5. Candidate Genes for %Ndfa-Associated QTLs
4.6. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Herridge, D.F.; Peoples, M.B.; Boddey, R.M. Global Inputs of Biological Nitrogen Fixation in Agricultural Systems. Plant Soil 2008, 311, 1–18. [Google Scholar] [CrossRef]
- Guo, J.; Wang, Y.; Song, C.; Zhou, J.; Qiu, L.; Huang, H.; Wang, Y. A Single Origin and Moderate Bottleneck during Domestication of Soybean (Glycine Max): Implications from Microsatellites and Nucleotide Sequences. Ann. Bot. 2010, 106, 505–514. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chu, S.; Zhang, H.; Zhu, Y.; Cheng, H.; Yu, D. Development and Application of a Novel Genome-Wide SNP Array Reveals Domestication History in Soybean. Nature 2016, 6, 20728. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Zhang, M.; Feng, F.; Tian, Z. Toward a “Green Revolution” for Soybean. Mol. Plant 2020, 13, 688–697. [Google Scholar] [CrossRef]
- SOY Canada 2022. Available online: https://soycanada.ca/ (accessed on 5 January 2023).
- Wilhite, D.A. Drought as a Natural Hazard: Concepts and Definitions. In Drought Mitigation Center Faculty Publications; Routledge: London, UK, 2000; pp. 1–21. [Google Scholar]
- Bonsal, B.; Regier, M. Historical Comparison of the 2001/2002 Drought in the Canadian Prairies. Clim. Res. 2007, 33, 229–242. [Google Scholar] [CrossRef] [Green Version]
- Quero, G.; Simondi, S.; Ceretta, S.; Otero, Á.; Garaycochea, S.; Fernández, S.; Borsani, O.; Bonnecarrère, V. An Integrative Analysis of Yield Stability for a GWAS in a Small Soybean Breeding Population. Crop Sci. 2021, 61, 1903–1914. [Google Scholar] [CrossRef]
- Wei, Y.; Jin, J.; Jiang, S.; Ning, S.; Liu, L. Quantitative Response of Soybean Development and Yield to Drought Stress during Different Growth Stages in the Huaibei Plain, China. Agronomy 2018, 8, 97. [Google Scholar] [CrossRef] [Green Version]
- Farooq, M.; Gogoi, N.; Barthakur, S.; Baroowa, B.; Bharadwaj, N.; Alghamdi, S.S.; Siddique, K.H.M. Drought Stress in Grain Legumes during Reproduction and Grain Filling. J. Agron. Crop Sci. 2017, 203, 81–102. [Google Scholar] [CrossRef]
- Arya, H.; Singh, M.B.; Bhalla, P.L. Towards Developing Drought-Smart Soybeans. Front. Plant Sci. 2021, 12, 750664. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Jiang, Y.; Dong, Y.; Wang, L.; Wang, W.; Ma, Z.; Yan, C.; Ma, C.; Liu, L. A Study on Soybean Responses to Drought Stress and Rehydration. Saudi J. Biol. Sci. 2019, 26, 2006–2017. [Google Scholar] [CrossRef]
- Desclaux, D.; Huynh, T.T.; Roumet, P. Identification of Soybean Plant Characteristics That Indicate the Timing of Drought Stress. Crop Sci. 2000, 40, 716–722. [Google Scholar] [CrossRef]
- Frederick, J.R.; Camp, C.R.; Bauer, P.J. Crop Ecology, Production & Management: Drought-Stress Effects on Branch and Mainstem Seed Yield and Yield Components of Determinate Soybean. Crop Sci. 2001, 41, 759–763. [Google Scholar] [CrossRef]
- Du, Y.; Zhao, Q.; Chen, L.; Yao, X.; Xie, F. Effect of Drought Stress at Reproductive Stages on Growth and Nitrogen Metabolism in Soybean. Agronomy 2020, 10, 302. [Google Scholar] [CrossRef] [Green Version]
- Thilakarathna, M.S.; Raizada, M.N. A Meta-Analysis of the Effectiveness of Diverse Rhizobia Inoculants on Soybean Traits under Field Conditions. Soil Biol. Biochem. 2017, 105, 177–196. [Google Scholar] [CrossRef]
- Hungria, M.; Vargas, M.A.T. Environmental Factors Affecting N2 Fixation in Grain Legumes in the Tropics, with an Emphasis on Brazil. Field Crops Res. 2000, 65, 151–164. [Google Scholar] [CrossRef]
- Hungria, M.; Bohrer, T.R.J. Variability of Nodulation and Dinitrogen Fixation Capacity among Soybean Cultivars. Biol. Fertil. Soils 2000, 31, 45–52. [Google Scholar] [CrossRef]
- Santachiara, G.; Salvagiotti, F.; Rotundo, J.L. Nutritional and Environmental Effects on Biological Nitrogen Fixation in Soybean: A Meta-Analysis. Field Crops Res. 2019, 240, 106–115. [Google Scholar] [CrossRef]
- Serraj, R.; Sinclair, T.R.; Purcell, L.C. Symbiotic N2 Fixation Response to Drought. J. Exp. Bot. 1999, 50, 143–155. [Google Scholar] [CrossRef] [Green Version]
- Serraj, R. Effects of Drought Stress on Legume Symbiotic Nitrogen Fixation: Physiological Mechanisms. Indian J. Exp. Biol. 2003, 41, 1136–1141. [Google Scholar]
- Sulieman, S.; Tran, L.S.P. Legume Nitrogen Fixation in a Changing Environment: Achievements and Challenges; Springer International Publishing: Cham, Switzerland, 2016; ISBN 9783319062129. [Google Scholar]
- Arreseigorl, C.; Gonzalezl, E.M.; Anthony, J.; Minchin, F.R.; Galvezl, L.; Cabrerizol, P.M.; Aricio-tejol, P.M.A.P. Sucrose Synthase and Nodule Nitrogen Fixation under Drought and Other Environmental Stresses. Symbiosis 1999, 27, 189–212. [Google Scholar]
- Einsle, O.; Rees, D.C. Structural Enzymology of Nitrogenase Enzymes. Chem. Rev. 2020, 120, 4969–5004. [Google Scholar] [CrossRef] [PubMed]
- Arrese-Igor, C.; Esther González, B.M.; Daniel Marino, B.; Rubén Ladrera, B.; Estíbaliz Larrainzar, B.; Erena Gil-Quintana, B. Physiological Responses of Legume Nodules to Drought. Plant Stress 2011, 5, 24–31. [Google Scholar]
- Collier, R.; Tegeder, M. Soybean Ureide Transporters Play a Critical Role in Nodule Development, Function and Nitrogen Export. Plant J. 2012, 72, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Kunert, K.J.; Vorster, B.J.; Fenta, B.A.; Kibido, T.; Giuseppe, D.; Foyer, C.H. Drought Stress Responses in Soybean Roots and Nodules. Front. Plant Sci. 2016, 7, 1015. [Google Scholar] [CrossRef] [Green Version]
- Torkamaneh, D.; Chalifour, F.P.; Beauchamp, C.J.; Agrama, H.; Boahen, S.; Maaroufi, H.; Rajcan, I.; Belzile, F. Genome-Wide Association Analyses Reveal the Genetic Basis of Biomass Accumulation under Symbiotic Nitrogen Fixation in African Soybean. Theor. Appl. Genet. 2020, 133, 665–676. [Google Scholar] [CrossRef]
- Hu, D.; Zhang, H.; Du, Q.; Hu, Z.; Yang, Z.; Li, X.; Wang, J.; Huang, F.; Yu, D.; Wang, H.; et al. Genetic Dissection of Yield-Related Traits via Genome-Wide Association Analysis across Multiple Environments in Wild Soybean (Glycine Soja Sieb. and Zucc.). Planta 2020, 251, 39. [Google Scholar] [CrossRef]
- Contreras-Soto, R.I.; Mora, F.; De Oliveira, M.A.R.; Higashi, W.; Scapim, C.A.; Schuster, I. A Genome-Wide Association Study for Agronomic Traits in Soybean Using SNP Markers and SNP-Based Haplotype Analysis. PLoS ONE 2017, 12, 0171105. [Google Scholar] [CrossRef]
- Adeboye, K.A.; Akhter Bhat, J.; Ganie, S.A.; Varshney, R.K.; Yu, D. Genome-Wide Association Analysis of Yield-Related Traits of Soybean Using Haplotype-Based Framework. Biol. Life Sci. Forum 2021, 11, 49. [Google Scholar]
- Priyanatha, C.; Torkamaneh, D.; Rajcan, I. Genome-Wide Association Study of Soybean Germplasm Derived from Canadian × Chinese Crosses to Mine for Novel Alleles to Improve Seed Yield and Seed Quality Traits. Front. Plant Sci. 2022, 13, 823. [Google Scholar] [CrossRef]
- Li, X.; Zhou, Y.; Bu, Y.; Wang, X.; Zhang, Y.; Guo, N.; Zhao, J.; Xing, H. Genome-Wide Association Analysis for Yield-Related Traits at the R6 Stage in a Chinese Soybean Mini Core Collection. Genes Genom. 2021, 43, 897–912. [Google Scholar] [CrossRef]
- Ravelombola, W.; Qin, J.; Shi, A.; Song, Q.; Yuan, J.; Wang, F.; Chen, P.; Yan, L.; Feng, Y.; Zhao, T.; et al. Genome-Wide Association Study and Genomic Selection for Yield and Related Traits in Soybean. PLoS ONE 2021, 16, e0255761. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Xu, W.; Zhang, H.; Liu, X.; Cui, X.; Li, S.; Song, L.; Zhu, Y.; Chen, X.; Chen, H. Comparative Selective Signature Analysis and High-Resolution GWAS Reveal a New Candidate Gene Controlling Seed Weight in Soybean. Theor. Appl. Genet. 2021, 134, 1329–1341. [Google Scholar] [CrossRef] [PubMed]
- Jo, H.; Lee, J.Y.; Lee, J.D. Genome-Wide Association Mapping for Seed Weight in Soybean with Black Seed Coats and Green Cotyledons. Agronomy 2022, 12, 250. [Google Scholar] [CrossRef]
- Chen, L.; Fang, Y.; Li, X.; Zeng, K.; Chen, H.; Zhang, H.; Yang, H.; Cao, D.; Hao, Q.; Yuan, S.; et al. Identification of Soybean Drought-Tolerant Genotypes and Loci Correlated with Agronomic Traits Contributes New Candidate Genes for Breeding. Plant Mol. Biol. 2020, 102, 109–122. [Google Scholar] [CrossRef]
- Dhanapal, A.P.; Ray, J.D.; Singh, S.K.; Hoyos-Villegas, V.; Smith, J.R.; Purcell, L.C.; King, C.A.; Fritschi, F.B. Genome-Wide Association Analysis of Diverse Soybean Genotypes Reveals Novel Markers for Nitrogen Traits. Plant Genome 2015, 8, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Malle, S.; Eskandari, M.; Morrison, M.; Belzile, F. Genome-Wide Association Identifies Several QTLs Controlling Cysteine and Methionine Content in Soybean Seed Including Some Promising Candidate Genes. Sci. Rep. 2020, 10, 21812. [Google Scholar] [CrossRef]
- Thilakarathna, M.S.; Moroz, N.; Raizada, M.N. A Biosensor-Based Leaf Punch Assay for Glutamine Correlates to Symbiotic Nitrogen Fixation Measurements in Legumes to Permit Rapid Screening of Rhizobia Inoculants under Controlled Conditions. Front. Plant Sci. 2017, 8, 1714. [Google Scholar] [CrossRef] [Green Version]
- Earl, H.J. A Precise Gravimetric Method for Simulating Drought Stress in Pot Experiments. Crop Sci. 2003, 43, 1868–1873. [Google Scholar] [CrossRef]
- Liyanage, D.K.; Chathuranga, I.; Mori, B.A.; Thilakarathna, M.S. A Simple, Semi-Automated, Gravimetric Method to Simulate Drought Stress on Plants. Agronomy 2022, 12, 349. [Google Scholar] [CrossRef]
- Thilakarathna, M.S.; Papadopoulos, Y.A.; Rodd, A.V.; Grimmett, M.; Fillmore, S.A.E.; Crouse, M.; Prithiviraj, B. Nitrogen Fixation and Transfer of Red Clover Genotypes under Legume–Grass Forage Based Production Systems. Nutr. Cycl. Agroecosyst. 2016, 106, 233–247. [Google Scholar] [CrossRef]
- Thilakarathna, M.S.; Torkamaneh, D.; Bruce, R.W.; Rajcan, I.; Chu, G.; Grainger, C.M.; Szczyglowski, K.; Hill, B.; Raizada, M.N. Testing Whether Pre-Pod-Fill Symbiotic Nitrogen Fixation in Soybean Is Subject to Drift or Selection over 100 Years of Soybean Breeding. Front. Agron. 2021, 3, 725813. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Sonah, H.; Bastien, M.; Iquira, E.; Tardivel, A.; Légaré, G.; Boyle, B.; Normandeau, É.; Laroche, J.; Larose, S.; Jean, M.; et al. An Improved Genotyping by Sequencing (GBS) Approach Offering Increased Versatility and Efficiency of SNP Discovery and Genotyping. PLoS ONE 2013, 8, e54603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torkamaneh, D.; Laroche, J.; Tardivel, A.; O’Donoughue, L.; Cober, E.; Rajcan, I.; Belzile, F. Comprehensive Description of Genomewide Nucleotide and Structural Variation in Short-Season Soya Bean. Plant Biotechnol. J. 2018, 16, 749–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torkamaneh, D.; Laroche, J.; Bastien, M.; Abed, A.; Belzile, F. Fast-GBS: A New Pipeline for the Efficient and Highly Accurate Calling of SNPs from Genotyping-by-Sequencing Data. BMC Bioinform. 2017, 18, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome Sequence of the Palaeopolyploid Soybean. Nature 2010, 463, 178–183. [Google Scholar] [CrossRef] [Green Version]
- Browning, B.L.; Browning, S.R. Genotype Imputation with Millions of Reference Samples. Am. J. Hum. Genet. 2016, 98, 116–126. [Google Scholar] [CrossRef] [Green Version]
- Torkamaneh, D.; Belzile, F. Accurate Imputation of Untyped Variants from Deep Sequencing Data. Deep. Seq. Data Anal. 2021, 2243, 271–281. [Google Scholar]
- Danecek, P.; Auton, A.; Abecasis, G.; Albers, C.A.; Banks, E.; DePristo, M.A.; Handsaker, R.E.; Lunter, G.; Marth, G.T.; Sherry, S.T.; et al. The Variant Call Format and VCFtools. Bioinformatics 2011, 27, 2156–2158. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Raj, A.; Stephens, M.; Pritchard, J.K. FastSTRUCTURE: Variational Inference of Population Structure in Large SNP Data Sets. Genetics 2014, 197, 573–589. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative Usage of Fixed and Random Effect Models for Powerful and Efficient Genome-Wide Association Studies. Public Libr. Sci. 2016, 12, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Zhang, H.; Tang, Z.; Xu, J.; Yin, D.; Zhang, Z.; Yuan, X.; Zhu, M.; Zhao, S.; Li, X.; et al. RMVP: A Memory-Efficient, Visualization-Enhanced, and Parallel-Accelerated Tool for Genome-Wide Association Study. Genom. Proteom. Bioinform. 2021, 19, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Grant, D.; Nelson, R.T.; Cannon, S.B.; Shoemaker, R.C. SoyBase, the USDA-ARS Soybean Genetics and Genomics Database. Nucleic Acids Res. 2009, 38, D843–D846. [Google Scholar] [CrossRef]
- Waese, J.; Fan, J.; Pasha, A.; Yu, H.; Fucile, G.; Shi, R.; Cumming, M.; Kelley, L.A.; Sternberg, M.J.; Krishnakumar, V.; et al. ePlant: Visualizing and Exploring Multiple Levels of Data for Hypothesis Generation in Plant Biology. Plant Cell 2017, 29, 1806–1821. [Google Scholar] [CrossRef] [Green Version]
- Seck, W.; Torkamaneh, D.; Belzile, F. Comprehensive Genome-Wide Association Analysis Reveals the Genetic Basis of Root System Architecture in Soybean. Front. Plant Sci. 2020, 11, 590740. [Google Scholar] [CrossRef]
- Yan, L.; Hofmann, N.; Li, S.; Ferreira, M.E.; Song, B.; Jiang, G.; Ren, S.; Quigley, C.; Fickus, E.; Cregan, P.; et al. Identification of QTL with Large Effect on Seed Weight in a Selective Population of Soybean with Genome-Wide Association and Fixation Index Analyses. BMC Genom. 2017, 18, 529. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Song, Q.; Cregan, P.B.; Jiang, G.L. Genome-Wide Association Study, Genomic Prediction and Marker-Assisted Selection for Seed Weight in Soybean (Glycine Max). Theor. Appl. Genet. 2016, 129, 117–130. [Google Scholar] [CrossRef] [Green Version]
- Sonah, H.; O’Donoughue, L.; Cober, E.; Rajcan, I.; Belzile, F. Identification of Loci Governing Eight Agronomic Traits Using a GBS-GWAS Approach and Validation by QTL Mapping in Soya Bean. Plant Biotechnol. J. 2015, 13, 211–221. [Google Scholar] [CrossRef]
- Copley, T.R.; Duceppe, M.O.; O’Donoughue, L.S. Identification of Novel Loci Associated with Maturity and Yield Traits in Early Maturity Soybean Plant Introduction Lines. BMC Genom. 2018, 19, 167. [Google Scholar] [CrossRef] [Green Version]
- Vance, C.P.; Gantt, S.J. Control of Nitrogen and Carbon Metabolism in Root Nodules. Physiol. Plant 1992, 85, 266–274. [Google Scholar] [CrossRef]
- Gonzalez, E.M.; Gordon, A.J.; James, C.L. The Role of Sucrose Synthase in the Response of Soybean Nodules to Drought. J. Exp. Bot. 1995, 46, 1515–1523. [Google Scholar] [CrossRef]
- King, C.A.; Purcell, L.C. Inhibition of N2 Fixation in Soybean Is Associated with Elevated Ureides and Amino Acids. Plant Physiol. 2005, 137, 1389–1396. [Google Scholar] [CrossRef] [Green Version]
- Mustofa, A.; Zubaidah, S.; Kuswantoro, H. Correlation and Path Analysis on Yield and Yield Components in Segregating Populations. Genetika 2021, 53, 157–166. [Google Scholar] [CrossRef]
- Hao, D.; Cheng, H.; Yin, Z.; Cui, S.; Zhang, D.; Wang, H.; Yu, D. Identification of Single Nucleotide Polymorphisms and Haplotypes Associated with Yield and Yield Components in Soybean (Glycine Max) Landraces across Multiple Environments. Theor. Appl. Genet. 2012, 124, 447–458. [Google Scholar] [CrossRef]
- Silva, A.F.; Sediyama, T.; Silva, F.C.S.; Bezerra, A.R.G.; Ferreira, L.V. Correlation and Path Analysis of Yield Components in Soybean Varieties. Turk. J. Field Crops 2015, 10, 177–179. [Google Scholar]
- Ciampitti, I.A.; Salvagiotti, F. New Insights into Soybean Biological Nitrogen Fixation. Agron. J. 2018, 110, 1185–1196. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Gao, H.; Wang, H.; Guo, Y.; He, M.; Peng, Y.; Wang, X. GSK3-Mediated Stress Signaling Inhibits Legume–Rhizobium Symbiosis by Phosphorylating GmNSP1 in Soybean. Mol. Plant 2021, 14, 488–502. [Google Scholar] [CrossRef]
- Singh, J.; Verma, P.K. NSP1 Allies with GSK3 to Inhibit Nodule Symbiosis. Trends Plant Sci. 2021, 26, 999–1001. [Google Scholar] [CrossRef]
- Phang, T.-H.; Li, M.-W.; Cheng, C.-C.; Wong, F.-L.; Chan, C.; Lam, H.-M. Molecular Responses to Osmotic Stresses in Soybean. In Soybean Molecular Aspects of Breeding; InTech: Rijeka, Croatia, 2011; pp. 215–240. [Google Scholar] [CrossRef] [Green Version]
- Taji, T.; Ohsumi, C.; Iuchi, S.; Seki, M.; Kasuga, M.; Kobayashi, M.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Important Roles of Drought- and Cold-Inducible Genes for Galactinol Synthase in Stress Tolerance in Arabidopsis thaliana. Plant J. 2002, 29, 417–426. [Google Scholar] [CrossRef]
- Bellaloui, N.; Hu, Y.; Mengistu, A.; Kassem, M.A.; Abel, C.A. Effects of Foliar Boron Application on Seed Composition, Cell Wall Boron, and Seed δ15N and δ13C Isotopes in Water-Stressed Soybean Plants. Front. Plant Sci. 2013, 4, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Oh, M.W.; Komatsu, S. Characterization of S-Adenosylmethionine Synthetases in Soybean under Flooding and Drought Stresses. Biol. Plant 2016, 60, 269–278. [Google Scholar] [CrossRef]
- Yuan, S.; Li, X.; Li, R.; Wang, L.; Zhang, C.; Chen, L.; Hao, Q.; Zhang, X.; Chen, H.; Shan, Z.; et al. Genome-Wide Identification and Classification of Soybean C2H2 Zinc Finger Proteins and Their Expression Analysis in Legume-Rhizobium Symbiosis. Front. Microbiol. 2018, 9, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.Y.; Lu, Z.W.; Sun, Y.; Fang, Z.W.; Chen, J.; Zhou, Y.B.; Chen, M.; Ma, Y.Z.; Xu, Z.S.; Min, D.H. The Ankyrin-Repeat Gene GmANK114 Confers Drought and Salt Tolerance in Arabidopsis and Soybean. Front. Plant Sci. 2020, 11, 584167. [Google Scholar] [CrossRef]
- Son, S.; Oh, C.J.; Bae, J.H.; Lee, H.; An, C.S. GmREM1.1 and GmREM2.1, Which Encode the Remorin Proteins in Soybean, Have Distinct Roles during Root Nodule Development. J. Plant Biol. 2015, 58, 17–25. [Google Scholar] [CrossRef]
- Gordon, A.J.; Minchin, F.R.; Skøt, L.; James, C.L. Stress-Induced Declines in Soybean N2 Fixation Are Related to Nodule Sucrose Synthase Activity. Plant Physiol. 1997, 114, 937–946. [Google Scholar] [CrossRef] [Green Version]
- Marcolino-Gomes, J.; Rodrigues, F.A.; Fuganti-Pagliarini, R.; Bendix, C.; Nakayama, T.J.; Celaya, B.; Molinari, H.B.C.; De Oliveira, M.C.N.; Harmon, F.G.; Nepomuceno, A. Diurnal Oscillations of Soybean Circadian Clock and Drought Responsive Genes. PLoS ONE 2014, 9, e86402. [Google Scholar] [CrossRef] [Green Version]
- Le, D.T.; Nishiyama, R.; Watanabe, Y.; Tanaka, M.; Seki, M.; Ham, L.H.; Yamaguchi-Shinozaki, K.; Shinozaki, K.; Tran, L.S.P. Differential Gene Expression in Soybean Leaf Tissues at Late Developmental Stages under Drought Stress Revealed by Genome-Wide Transcriptome Analysis. PLoS ONE 2012, 7, e495. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Moisture Effect | Chr Number | MSS Position | QTL ID | Minor Allele Frequency | p Value | Effect |
|---|---|---|---|---|---|---|
| 30% FC | 10 | 37,995,110 | qNDFA-30-1 | 0.19 | <0.0001 | −6.87 |
| 13 | 13,866,995 | qNDFA-30-2 | 0.29 | <0.0001 | −4.79 | |
| Relative | 6 | 18,244,365 | qNDFA-RP-1 | 0.06 | <0.0001 | −0.12 |
| Performance | 14 | 994,141 | qNDFA-RP-2 | 0.12 | <0.0001 | −0.05 |
| 19 | 46,575,733 | qNDFA-RP-3 | 0.11 | <0.0001 | −0.09 |
| Treatment | Chr Number | MSS Position | REF/ALT | QTL ID | Candidate Genes | Orthologous Genes in Arabidopsis | Annotations |
|---|---|---|---|---|---|---|---|
| 30% FC | 10 | 37,995,110 | G/A | qNDFA-30-1 | Glyma.10g144600 | Gene Model: AT5G26751.1 | Glycogen synthase kinase—FJ460228 |
| Glyma.10g144600 | Gene Model: AT5G26751.1 | Glycogen synthase kinase-3—BT093874 | |||||
| Glyma.10g145300 | Gene Model: AT2G47180.1 | Galactinol synthase 1- AK245720, AY126715 | |||||
| Glyma.10g144300 | Gene Model: AT4G01850.1 | S-adenosylmethionine synthetase 2 | |||||
| Relative Performance | 6 | 18,244,365 | G/A | qNDFA-RP-1 | Glyma.06g197700 | Gene Model: AT5G61190.1 | Putative endonuclease or glycosyl hydrolase with C2H2-type zinc finger domain |
| Glyma.06g198600 | Gene Model: AT2G03430.1 | Ankyrin repeat family protein | |||||
| Glyma.06g199700 | Gene Model: AT5G61280.1 | Remorin family protein | |||||
| 19 | 46,575,733 | C/T | qNDFA-RP-3 | Glyma.19g212800 | Gene Model: AT4G02280.1 | Sucrose synthase 3 | |
| Glyma.19g213900 | Gene Model: AT4G02200.1 | Drought-responsive family protein |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 His Majesty the King in Right of Canada. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liyanage, D.K.; Torkamaneh, D.; Belzile, F.; Balasubramanian, P.; Hill, B.; Thilakarathna, M.S. The Genotypic Variability among Short-Season Soybean Cultivars for Nitrogen Fixation under Drought Stress. Plants 2023, 12, 1004. https://doi.org/10.3390/plants12051004
Liyanage DK, Torkamaneh D, Belzile F, Balasubramanian P, Hill B, Thilakarathna MS. The Genotypic Variability among Short-Season Soybean Cultivars for Nitrogen Fixation under Drought Stress. Plants. 2023; 12(5):1004. https://doi.org/10.3390/plants12051004
Chicago/Turabian StyleLiyanage, Dilrukshi Kombala, Davoud Torkamaneh, François Belzile, Parthiba Balasubramanian, Brett Hill, and Malinda S. Thilakarathna. 2023. "The Genotypic Variability among Short-Season Soybean Cultivars for Nitrogen Fixation under Drought Stress" Plants 12, no. 5: 1004. https://doi.org/10.3390/plants12051004