1. Introduction
Biological control agents (BCAs) are important instruments to reduce pests and diseases with the aim of protecting crops. They are defined as living agents that constantly target pests and can be classified into four different categories: whether they are added for permanent or temporary establishment, and whether they are used with or without targeted human intervention [
1]. Moreover, antimicrobial secondary metabolites produced by agriculturally important fungi can also be used as a green solution to protect crops. They are low-molecular-mass metabolites produced during the late growth phase and are divided into different biosynthetic categories such as metabolites derived from shikimic acid, those derived from amino acids, those derived from acetyl-CoA and those derived from sugars [
2]. These strategies would prevent the use of synthetic pesticides or reduce their use, in line with ongoing European Union proposals [
3].
The
Trichoderma genus contains BCAs that are mainly used in biocontrol for agricultural purposes due to their versatility and capability of adaptation [
4]. More than 375 species have been described [
5] and this fungus is one of the most bio-based products worldwide [
6]. As BCAs,
Trichoderma strains can display a great variety of biocontrol mechanisms, such as mycoparasitism, lytic enzymes, antibiosis, secondary metabolites, competition with pathogens and the soil microbial community, plant root colonization and the induction of plant defense responses [
7].
Vitis vinifera L. (grapevine plant) cultivars are the most widely planted in the world and require high production costs, of which one of the most important is associated with pest and disease management. Nowadays the most significant destructive diseases in viticulture are grapevine trunk diseases (GTDs) [
8]. A sustainable and eco-efficient approach is needed to solve this problem [
9]. Furthermore,
Trichoderma strains are among the most widely tested biological control agents against fungi involved in GTDs [
10]. However,
Trichoderma results after field trials are uncertain and still show great variability and disparity [
11,
12]. In the current study, first, we try to elucidate the factors that could favor the persistence of
Trichoderma in the soil in order to protect the roots of vine plants. Farming management practices in soils are an important source of variation in ecosystems because they modify microbial composition, including
Trichoderma, especially in vineyards [
13,
14,
15,
16,
17,
18]. Therefore, a survey of the main farming management practices was performed in this research to give us information about their influence on soil
Trichoderma populations. Herbicides are important tools for weed management and their use is increasing worldwide. In addition, they can alter the soil microbiome and nutrient composition of grapevine plants [
18]. Thus, surveys on the use (or not) of herbicides in rows of vineyards could show a correlation between soil health and
Trichoderma populations. In addition, tillage intensity can modify the soil population of microorganisms such as
Trichoderma spp., Actinomycetes and
Gliocladium spp. [
19]. In addition, fertilizers such as manure or chemical fertilizers could influence the microbial population [
20,
21]. Finally, the main physicochemical nutrients present in the soil have been shown to be an important source of variation in fungal and bacterial communities in soil vineyards [
19,
22,
23]. Thus, all of these variables will be evaluated in our research.
Another point to evaluate is the use of indigenous
Trichoderma from the soil to protect grapevine plants against one of the most prevalent fungi involved in GTDs,
Phaeoacremonium minimum. This pathogen is the pioneer fungus of Petri, Esca and grapevine leaf stripe diseases [
24]. Not only can it colonize woody tissues [
24] but is also a soilborne pathogen [
25]. Previous studies have shown that there are native
Trichoderma strains isolated from soils or grapevine plants that can protect grapevine plants from GTDs. In South Africa, for example, two strains (
T. atroviride T-77 and
T. atroviride USPP T1) can colonize the roots of grapevine plants and activate the host defense system of the plant [
26]. Another study showed that after testing 16
Trichoderma isolates from Southern Italy, some of them protect grapevine plants against a great number of fungal pathogens that cause GTDs [
27]. Moreover, native
Trichoderma strains have been isolated from vineyards in British Columbia, Canada, as effective biocontrol agents against Botryosphaeria dieback [
28]. In Spain, strains of
T. harzianum had positive results after their application in young vine plants [
29].
In this study we analyzed the following hypotheses: (i) soil physicochemical properties and farming management practices affect native Trichoderma populations, and (ii) Trichoderma native strains isolated from vineyard soils are able to control P. minimum. The specific objectives are to (1) isolate Trichoderma strains from the soil, (2) identify the relationship between the physicochemical soil characteristics and the abundance of Trichoderma and (3) identify Trichoderma strains for biocontrol of P. minimum in the soil.
3. Discussion
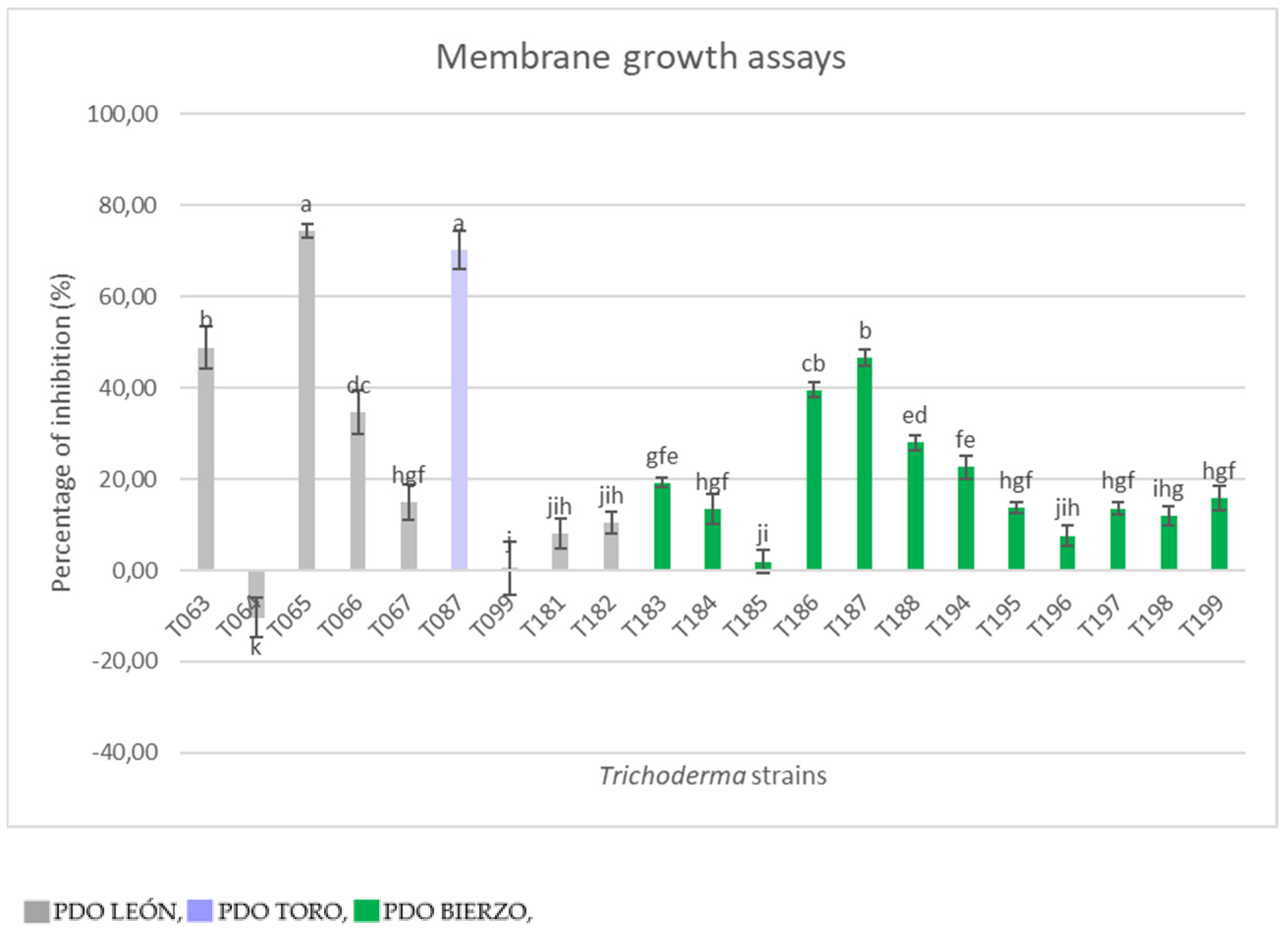
This study investigated the role of Trichoderma in the soil and the use of native strains and/or their metabolites as a solution for protecting grapevine plants from GTDs. We isolated Trichoderma from the soil to protect the roots of grapevine plants while investigating factors that favor its presence in the vineyard. We found that the presence of Trichoderma is positively correlated with a high content of iron in the soil and negatively correlated with the pH value. Moreover, management practices affect Trichoderma soil populations and they are negatively correlated with fertilization and tillage but positively correlated with spraying herbicides in soils. We isolated 21 Trichoderma strains, and first evaluated them for their antibiosis activity. Great variability among strains was found even in the same soils. Two strains were selected (T065 and T087) as effective producers of secondary metabolites with significant activity against P. minimum. Later, a second mechanism of biocontrol was evaluated: mycoparasitism. In this case, the pathogen’s growth was reduced using the two previously selected Trichoderma strains (T065 and T087). Both are promising candidate biological control agents, especially T087, whose high production of spores indicates that it is an appropriate candidate for mass production and commercialization.
First, we collected soil samples from each vineyard with the aim of analyzing its physicochemical characteristics and isolating
Trichoderma strains at the same time. In our study, the media used for
Trichoderma isolation contains dextrose, rose bengal and chloramphenicol, which are suitable for
Trichoderma species isolation [
32]. Only 21 different isolates were found from ten different vineyards. In addition, we did not isolate
Trichoderma spp. in two plots (ARE and TER). All of these plots are located in the inner plateau of Spain and the same climate is experienced by all of them, as described in a previous study of these locations [
33]. However, the soil microbial community could also play an important role [
34] and could drive different compositions of fungal and bacterial communities, as was demonstrated previously [
35,
36]. Other parameters such as the soil temperature, redox status of the soil and moisture could also play an important role [
37]. In this study we identified a negative correlation with pH; the lower the pH, the higher the presence of
Trichoderma. It has been demonstrated that the soil pH crucially influences the population of bacterial and fungal communities [
35,
37].This parameter is considered one of the major factors that affects the activity of
Trichoderma and this fungus has been found to be better adapted to acidic soil [
38]. Another study found that there is also a negative correlation between the pH and the activity of a
Trichoderma koningii biological control strain in agricultural soils of wheat [
39]. A recent study also found that some other biological control
Trichoderma strains (T029 and T059) had a negative correlation with the pH in soils in comparison to its abundance in the soil in bean fields [
40]. In our case, native
Trichoderma populations in vineyards of Castilla y León are increased in acidic soils (pH between 5.5 and 6.5). We also observed a positive correlation between the abundance of
Trichoderma and the iron content in soil.
Trichoderma is able to produce organic acids that permit the solubilization of phosphates or micronutrients and mineral cations [
38,
41]. Several studies have concluded that
Trichoderma strains (
Trichoderma asperellum T34,
Trichoderma asperellum T9) applied to the soil could increase the iron uptake of plants [
42,
43,
44,
45]. An example is harzianic acid, a novel siderophore produced by
Trichoderma harzianum M10 that alters nutrient availability in soil due to the mechanism of iron solubilization [
46]. In this study, this positive correlation could be due to a low pH that increased iron solubilization and the presence in a higher proportion of
Trichoderma strains in natural soils that could increase the availability of iron in soil among other microorganisms [
47]. This hypothesis could be contrasted and evaluated using a blue agar CAS assay for siderophore detection as possible future research [
48]. Using factor analysis in this research with small size samples produces a reliable degree of confidence [
49], but laboratory studies are needed to confirm both trends and the type of correlation. In this case, it is more important to unravel the type of correlation (causality or consequence) in these interactions to increase the
Trichoderma population in soils. Moreover, a deeper analysis is required in order to obtain more insights about the relationships between
Trichoderma and abiotic or biotic factors as described by Sorribas et al. 2008 [
50]. Regarding farming management practices, spraying herbicides in vineyard rows had a positive correlation with the population of
Trichoderma in our research.
Trichoderma indigenous isolates from soil in India demonstrated a high compatibility with all the isolates in comparison to some pesticides. Some of them, such as 2,4-D and glyphosate, showed compatibility with a few strains [
51]. Moreover, some
Trichoderma strains such as
Trichoderma viride strain FRP could degrade glyphosate [
52];
Trichoderma asperellum TJ01 is tolerant to an organophosphorus pesticide [
53].
Trichoderma atroviride UEL257 showed a higher tolerance to some of the herbicides tested and it was recommended to be sprayed in soils where there is a strong presence of herbicides [
54]. Recent studies emphasize the possibility of using glyphosate-eating fungi such as
Trichoderma,
Fusarium,
Aspergillum or
Penicillium to tolerate this pesticide [
55] and another study remarks the efficiency of glyphosate degradation by microorganisms such as
Penicillium,
Aspergillus and
Trichoderma [
56]. However, the use of herbicides sprayed in rows disrupts and reduces fungal communities and could lead to a reduction in crop production [
13] so that native
Trichoderma strains could develop a higher resistance to the pesticides sprayed in comparison to other fungi or bacteria, and thus could occupy this ecological niche due to its rapid growth and opportunism [
57]. This hypothesis should be tested and evaluated to see if these isolates are able to resist different herbicides. Furthermore, no tillage was found to help increase the population of native
Trichoderma strains. Tillage destroys fungal hyphae and reduces the presence of fungal communities [
58]. Another study shows that non-tillage favors fungi and more diverse fungal communities [
59]. In addition, in another study where the presence of
Trichoderma was evaluated in comparison to practice management, a higher presence of this fungus was found when no tillage was performed in the field [
19]. Finally, fertilization did not have a positive correlation with the presence of
Trichoderma in soils evaluated. Several studies show that in fertilized soils there is an increase in the microbial biomass, crop yield and crop quality compared to non-fertilized soils [
19,
21,
60,
61]. An increase in the
Trichoderma population was also found after applying fertilizers [
62]. In this study, no differences in terms of the type of fertilization (manure or chemical fertilizers) were described to facilitate a reliable statistical analysis. As significant differences were achieved when applying any type of fertilization, it could also be due to an alteration of the ratio of carbon to nitrogen in the soil (C/N) or to other nutrients that alter the composition of the microbiota in soil [
63]. To sum up, the highest presence of
Trichoderma was found in acid soils (pH 5.5–6.5) where non-tillage, fertilization and herbicide spraying are performed. For future perspectives, further assays are needed to separately confirm the factor that has a crucial incidence in this interaction between factors.
In terms of biocontrol, for an eco-sustainable agriculture,
Trichoderma is one of the most suitable biological control agents for this aim [
4]. In this study, fungi of the
Trichoderma genus were isolated in an attempt to reduce the incidence of fungi that cause GTDs through the roots. In this case,
Phaeoacremonium minimum was used as a very aggressive pathogen that can attack the plant via pruning wounds but also in the roots [
8]. In previous complementary studies,
Trichoderma strains were isolated from grapevine bark and tested against
Phaeoacremonium minimum with the aim of controlling infection in pruning wounds [
33], and
Trichoderma strains were selected to improve its performance—related to climatic factors [
64]—on these pruning wounds. In our study we selected
Trichoderma strains from the soil due to its capacity to produce secondary metabolites. Synthetic chemical pesticides have been used for years but their impact on human health and the environment has led to a change and a new approach to search for agriculturally important microorganisms such as
Trichoderma [
2]. Secondary metabolites of
Trichoderma have different biological roles, some of them exhibiting direct activity against plant pathogens, but also triggering plant defenses or enhancing vegetal growth [
65]. In our research,
in vitro cellophane membrane assays led us to select the
Trichoderma strains able to produce metabolites with antibiotic activity against
P. minimum. We obtained two strains that significantly reduced the growth of this pathogen. The first was T065, identified as
T. gamsii [
31]. Another fungus of this species (
T. gamsii strain ICC080), along with other
T. asperellum, have been used as biocontrol agents of esca and grapevine trunk diseases, and they have been able to reduce GTD incidence and protect grapevine plants [
66,
67]. Secondly, T087 was identified as
T. harzianum. In this case,
T. harzianum and
T. atroviride are the two species most widely used in the biocontrol of GTDs [
10].
T. harzianum has been used for protecting vine cuttings (
T. harzianum T39 Trichodex
®) [
68], and selected strains have been used to avoid the decline in pathogens in nursery grapevine plants using Trichoflow-T™ [
69]. Molecular identification is an important factor in unravelling the great disparity of results in
Trichoderma field assays and trying to fix the factors that govern each genotype’s function [
70]. One of the first steps is to reach the strain level because even
Trichoderma from the same species present differences [
71]. Moreover, merely observing molecular identification may lead to some problems, especially in the
harzianum–virens clade [
72]. If the whole genome of some biological control agents is sequenced and its genes are identified, interesting new synthetic pathways and other relations can be found, as described by Schmoll et al. [
73]. However, a final step is also necessary in order to confirm this effectiveness. It is important to test a biological control agent in a three-way interaction (
Trichoderma–plant–pathogen) [
7] before being sold on the market. Proteomic assays [
74], microscopic assays [
75] or metabolomic assays [
76] must be conducted in planta to confirm a proper control of pathogens and to reduce the great disparity in results described previously. Moreover, other strains of
T. harzianum have been described to protect plants from pruning-wound infections [
75,
77,
78]. In addition, the application of a major secondary metabolite (6-pentyl-α-pyrone (6PP)) produced by
Trichoderma strains (one of them was
T. harzianum T77) against
Eutypa lata,
Neofussicocum australe,
Neofusiccocum parvum and
Phaeomoniella chlamydospora (pathogens involved in GTDs) triggered the inhibition of ascospore germination and a reduction in mycelial growth [
79]. In our research, only diffusible antifungal compounds (DACs) have been evaluated. It could also be interesting to identify volatile organic compounds (VOCs), as has been described by Van Jaarsveld et al. [
80] against black foot disease pathogens. Our future work will evaluate the biocontrol mechanism of mycoparasitism, which would most likely be one of the ancestral lifestyles of
Trichoderma [
81]. We assayed our selected
Trichoderma strains (T065 and T087) against
P. minimum in a dual-culture assay and saw that both of them stopped the growth of this pathogen, as in another study described by Carro-Huerga et al. [
33] where native
Trichoderma strains isolated from the bark of grapevine plants could mycoparasitize
P. minimum. In dual-confrontation assays two types of mechanisms are used: antibiosis (as previously assayed in the membrane growth assay in this research) [
82] and mycoparasitism [
83,
84]. Therefore, we confirmed the biocontrol by performing an
in vitro test of two strains (T065 and T087). T087 could especially be a potential microorganism for mass production due to its capacity for high production spores, as most
Trichoderma-based products are commercialized in spores [
6].
To sum up, our results indicate that (i) physicochemical soil properties and farming management practices affect native Trichoderma populations and (ii) native Trichoderma strains isolated from vineyard soil biocontrol P. minimum. According to the former, the pH and iron content of soils are correlated to Trichoderma abundance and farming management practices also affect its presence. Regarding the second hypothesis, two Trichoderma strains (T065 Trichoderma gamsii and T087 Trichoderma harzianum) exhibited a significant biocontrol activity against P. minimum using antibiosis as the main mechanism. Future assays should evaluate siderophore production of the isolated Trichoderma strains, test the resistance to biodegrade herbicides, analyze secondary metabolite profiles of both Trichoderma strains selected and determine their major compounds to evaluate the suitability of their application in field.

 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}