

Effects of Exponential N Application on Soil Exchangeable Base Cations and the Growth and Nutrient Contents of Clonal Chinese Fir Seedlings
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Areas
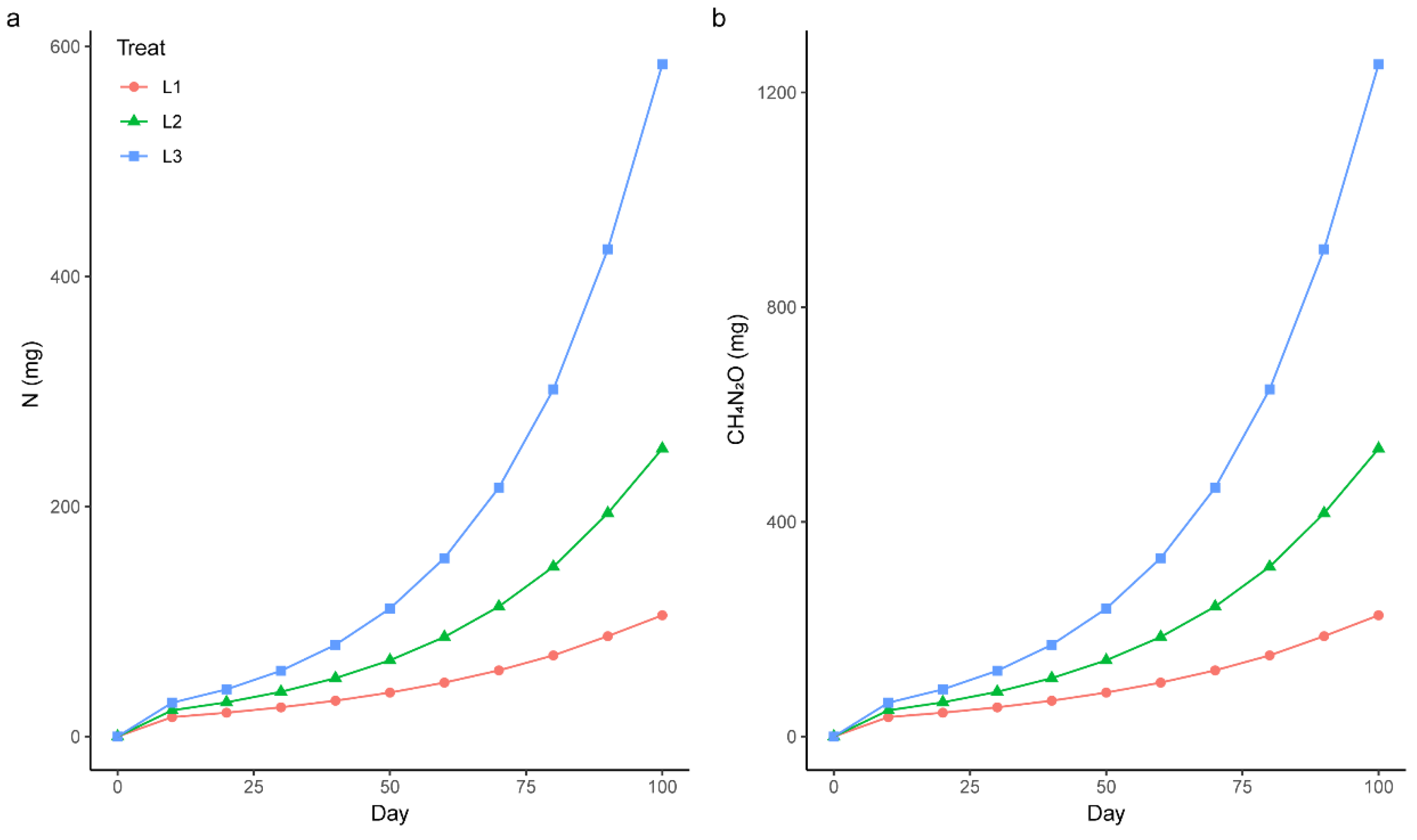
2.2. Experimental Design
2.3. Measurements
2.4. Statistical Analyses
3. Results
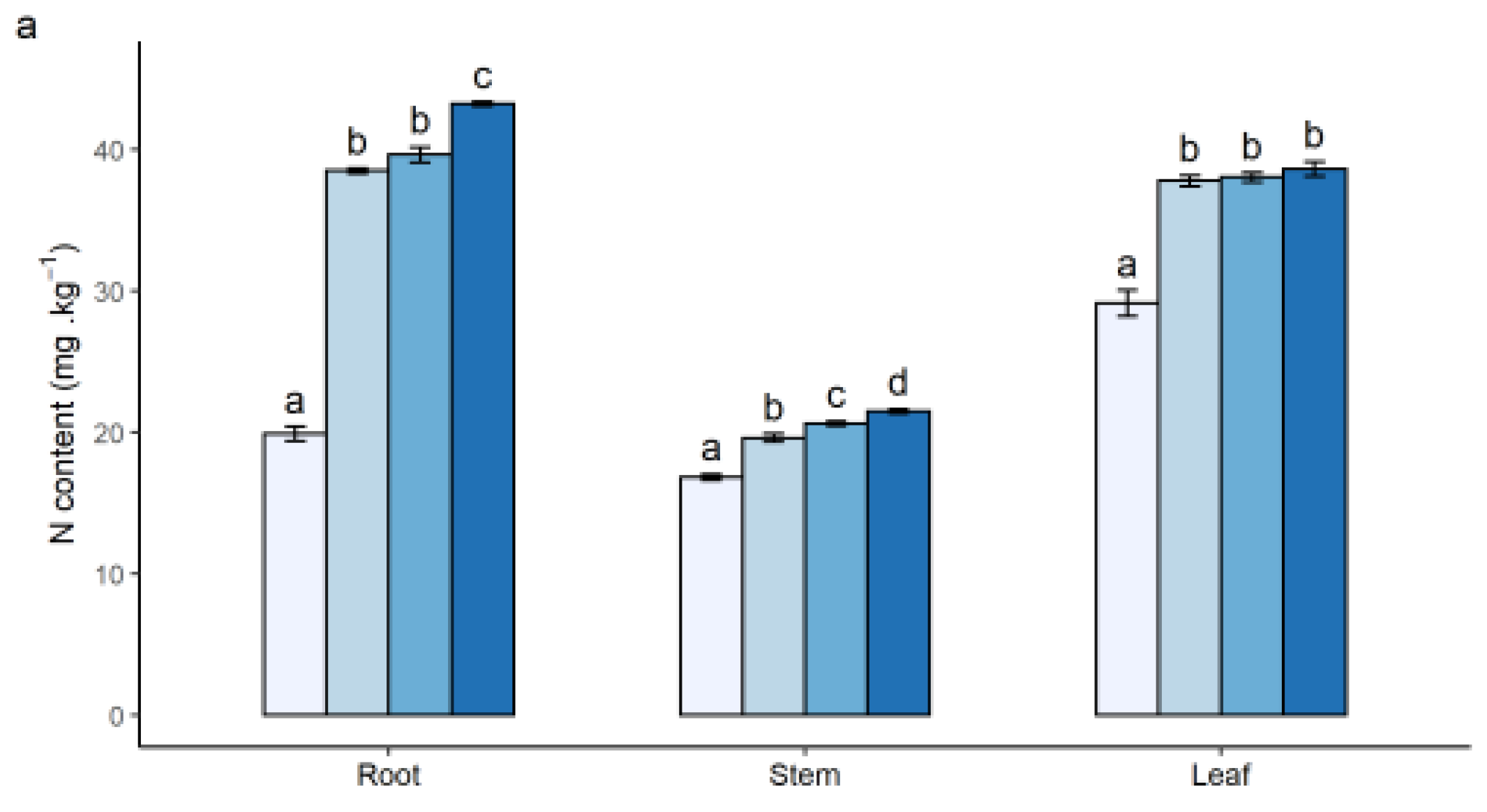
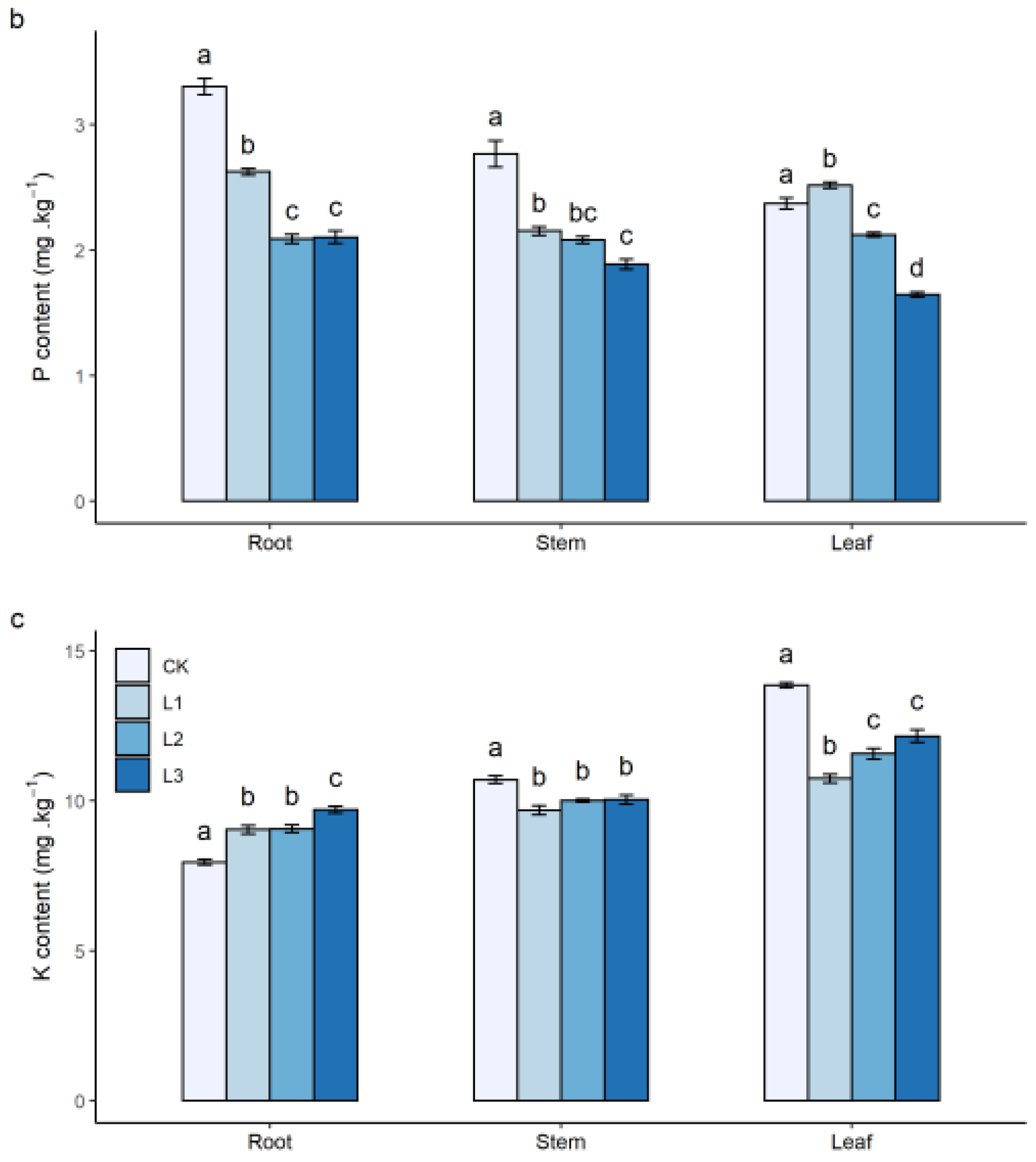
3.1. Effects of N Addition on Seedling Growth and Nutrient Content of Plants
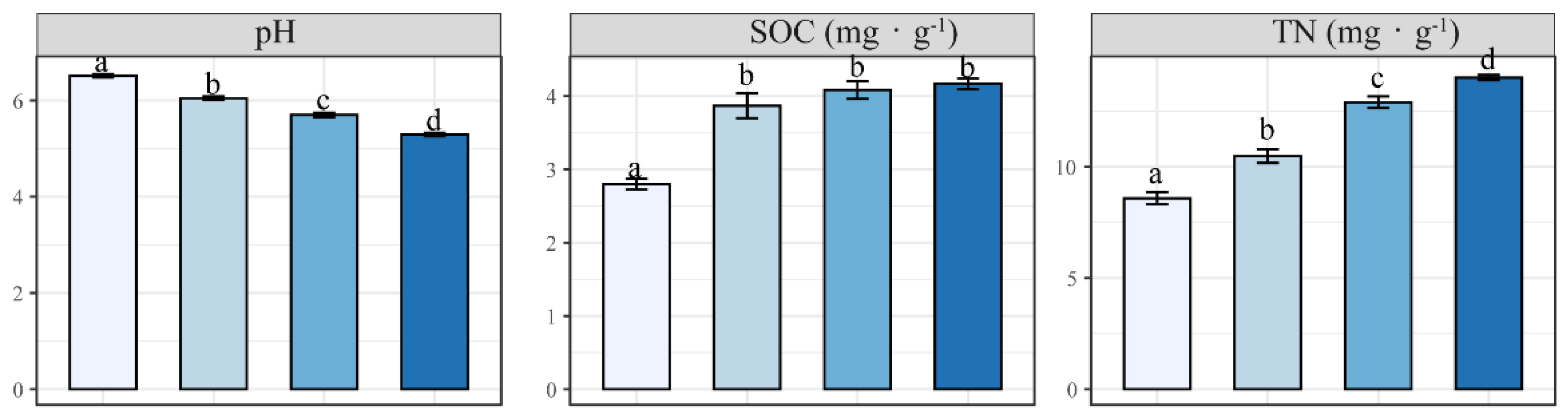
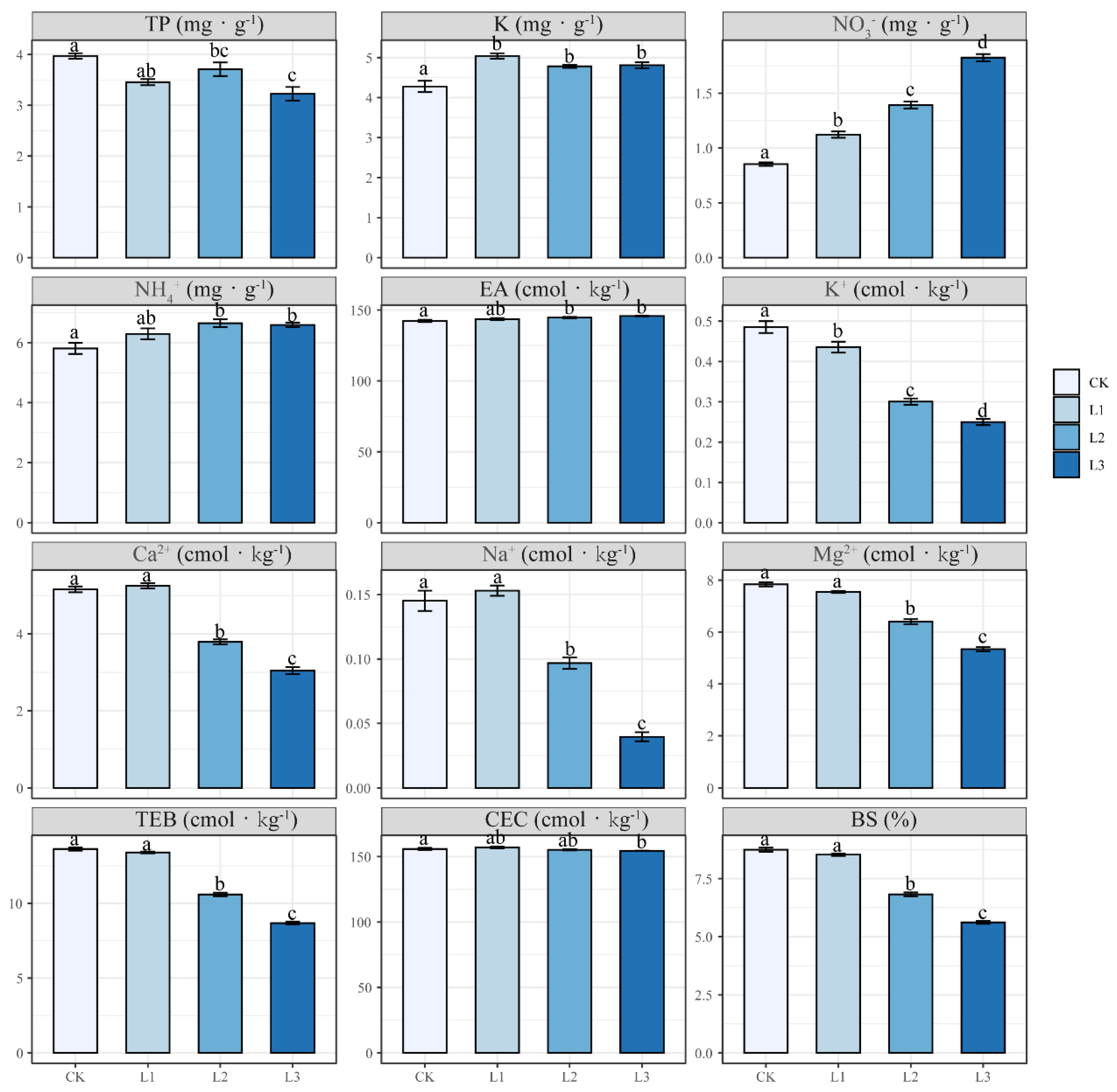
3.2. Effect of N Addition on Soil Physicochemical Properties
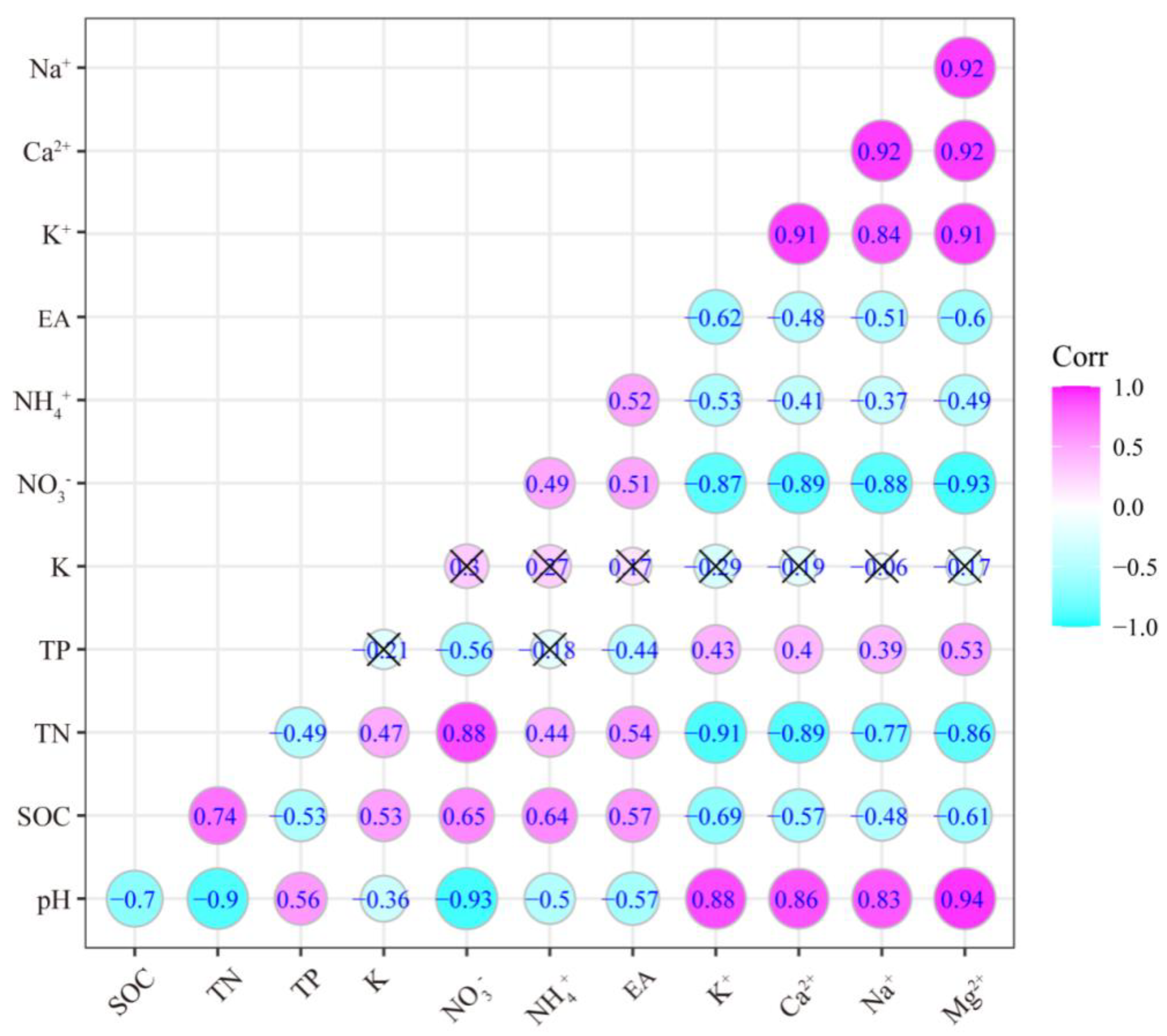
3.3. Relationships among Different Soil Parameters
3.4. The Correlations between Plant Growth and BCs
4. Discussion
4.1. Growth and Nutrient Contents of Seedlings
4.2. Response of Soil Parameters to N Addition
4.3. Effects of BCs on Chinese Fir Seedling Growth
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qaswar, M.; Li, D.C.; Huang, J.; Han, T.F.; Ahmed, W.; Abbas, M.; Zhang, L.; Du, J.X.; Khan, Z.H.; Ullah, S. Interaction of liming and long-term fertilization increased crop yield and phosphorus use efficiency (PUE) through mediating exchangeable cations in acidic soil under wheat-maize cropping system. Sci. Rep.-UK 2020, 10, 19828. [Google Scholar] [CrossRef] [PubMed]
- Pokharel, P.; Chang, S.X. Exponential fertilization promotes seedling growth by increasing nitrogen retranslocation in trembling aspen planted for oil sands reclamation. For. Ecol. Manag. 2016, 372, 35–43. [Google Scholar] [CrossRef]
- Fageria, N.K.; Dos Santos, A.B.; Moraes, M.F. Influence of Urea and Ammonium Sulfate on Soil Acidity Indices in Lowland Rice Production. Commun. Soil Sci. Plant Anal. 2010, 41, 1565–1575. [Google Scholar] [CrossRef]
- Munoz-Huerta, R.F.; Guevara-Gonzalez, R.G.; Contreras-Medina, L.M.; Torres-Pacheco, I.; Prado-Olivarez, J.; Ocampo-Velazquez, R.V. A Review of Methods for Sensing the Nitrogen Status in Plants: Advantages, Disadvantages and Recent Advances. Sensors. 2013, 13, 10823–10843. [Google Scholar] [CrossRef]
- Xu, X.J.; Timmer, V.R. Biomass and nutrient dynamics of Chinese fir seedlings under conventional and exponential fertilization regimes. Plant Soil. 1998, 203, 313–322. [Google Scholar] [CrossRef]
- Everett, K.T.; Hawkins, B.J.; Kiiskila, S. Growth and nutrient dynamics of Douglas-fir seedlings raised with exponential or conventional fertilization and planted with or without fertilizer. Can. J. For. Res. 2007, 37, 2552–2562. [Google Scholar] [CrossRef]
- Qu, L.Y.; Quoreshi, A.M.; Koike, T. Root growth characteristics, biomass and nutrient dynamics of seedlings of two larch species raised under different fertilization regimes. Plant Soil. 2003, 255, 293–302. [Google Scholar] [CrossRef]
- Timmer, V.R. Exponential nutrient loading: A new fertilization technique to improve seedling performance on competitive sites. New For. 1997, 13, 279–299. [Google Scholar] [CrossRef]
- Hawkins, B.J.; Burgess, D.; Mitchell, A.K. Growth and nutrient dynamics of western hemlock with conventional or exponential greenhouse fertilization and planting in different fertility conditions. Can. J. For. Res.-Rev. Can. Rech. For. 2005, 35, 1002–1016. [Google Scholar] [CrossRef]
- Tian, D.S.; Niu, S.L. A global analysis of soil acidification caused by nitrogen addition. Environ. Res. Lett. 2015, 10, 027019. [Google Scholar] [CrossRef]
- Bobbink, R.; Hicks, K.; Galloway, J.; Spranger, T.; Alkemade, R.; Ashmore, M.; Bustamante, M.; Cinderby, S.; Davidson, E.; Dentener, F.; et al. Global assessment of nitrogen deposition effects on terrestrial plant diversity: A synthesis. Ecological Applications 2010, 20, 30–59. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, X.H.; Jiang, Y.; Liu, X.S.; Zhu, F. Effects of N addition on soil exchangeable cations in a young Keteleeria fortunei var. cyclolepis forest. Ecoscience 2019, 26, 259–268. [Google Scholar] [CrossRef]
- Lucas, R.W.; Klaminder, J.; Futter, M.N.; Bishop, K.H.; Egnell, G.; Laudon, H.; Hogberg, P. A meta-analysis of the effects of nitrogen additions on base cations: Implications for plants, soils, and streams. For. Ecol. Manag. 2011, 262, 95–104. [Google Scholar] [CrossRef]
- McLaughlin, S.B.; Wimmer, R. Tansley Review No. 104—Calcium physiology and terrestrial ecosystem processes. New Phytol. 1999, 142, 373–417. [Google Scholar] [CrossRef]
- Likens, G.E.; Driscoll, C.T.; Buso, D.C.; Siccama, T.G.; Johnson, C.E.; Lovett, G.M.; Fahey, T.J.; Reiners, W.A.; Ryan, D.F.; Martin, C.W. The biogeochemistry of calcium at Hubbard Brook. Biogeochemistry 1998, 41, 89–173. [Google Scholar] [CrossRef]
- Bowman, W.D.; Cleveland, C.C.; Halada, L.; Hresko, J.; Baron, J.S. Negative impact of nitrogen deposition on soil buffering capacity. Nat. Geosci. 2008, 1, 767–770. [Google Scholar] [CrossRef]
- Cai, Z.J.; Wang, B.R.; Xu, M.G.; Zhang, H.M.; He, X.H.; Zhang, L.; Gao, S.D. Intensified soil acidification from chemical N fertilization and prevention by manure in an 18-year field experiment in the red soil of southern China. J. Soils Sediments 2015, 15, 260–270. [Google Scholar] [CrossRef]
- Priputina, I.; Zubkova, E.; Shanin, V.; Smirnov, V.; Komarov, A. Evidence of plant biodiversity changes as a result of nitrogen deposition in permanent pine forest plots in central Russia. Ecoscience 2014, 21, 286–300. [Google Scholar] [CrossRef]
- Bodirsky, B.L.; Popp, A.; Lotze-Campen, H.; Dietrich, J.P.; Rolinski, S.; Weindl, I.; Schmitz, C.; Muller, C.; Bonsch, M.; Humpenoder, F.; et al. Reactive nitrogen requirements to feed the world in 2050 and potential to mitigate nitrogen pollution. Nat. Commun. 2014, 5, 3858. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.L.; Zhang, H.Z.; Liu, T.; Zhang, W.X.; Shao, Y.H.; Ha, D.L.; Li, Y.Q.; Zhang, C.M.; Cai, X.A.; Rao, X.Q.; et al. Consistent effects of canopy vs. understory nitrogen addition on the soil exchangeable cations and microbial community in two contrasting forests. Sci. Total Environ. 2016, 553, 349–357. [Google Scholar] [CrossRef]
- Chen, D.M.; Lan, Z.C.; Bai, X.; Grace, J.B.; Bai, Y.F. Evidence that acidification-induced declines in plant diversity and productivity are mediated by changes in below-ground communities and soil properties in a semi-arid steppe. J. Ecol. 2013, 101, 1322–1334. [Google Scholar] [CrossRef]
- Stevens, C.J.; Thompson, K.; Grime, J.P.; Long, C.J.; Gowing, D.J. Contribution of acidification and eutrophication to declines in species richness of calcifuge grasslands along a gradient of atmospheric nitrogen deposition. Funct. Ecol. 2010, 24, 478–484. [Google Scholar] [CrossRef]
- Sheng, W.T.; Division of Forest Ecology, Chinese Society of Forestry (Eds.) Soil Degradation and its Control Techniques for Timber Plantations in China. In Research on Site Degradation of Timber Plantation; Chinese Press of Science and Technology: Beijing, China, 1992; pp. 1–7. [Google Scholar]
- Hu, R.; Sun, Y.; Wu, B.; Duan, H.; Zheng, H.; Hu, D.; Lin, H.; Tong, Z.; Xu, J.; Li, Y. Somatic Embryogenesis of Immature Cunninghamia lanceolata (Lamb.) Hook Zygotic Embryos. Sci. Rep. 2017, 7, 56. [Google Scholar] [CrossRef]
- Zhu, W.; Liu, T.; Liu, C.; Zhou, F.; Lai, X.E.; Hu, D.; Chen, J.; Huang, S. The Complete Chloroplast Genome Sequence of Cunninghamia lanceolata. Mitochondrial DNA Part A 2017, 28, 405–406. [Google Scholar] [CrossRef] [PubMed]
- Holland, E.A.; Dentener, F.J.; Braswell, B.H.; Sulzman, J.M. Contemporary and pre-industrial global reactive nitrogen budgets. Biogeochemistry 1999, 46, 7–43. [Google Scholar] [CrossRef]
- Zheng, M.H.; Zhang, W.; Luo, Y.Q.; Mori, T.K.; Mao, Q.Q.; Wang, S.H.; Huang, J.; Lu, X.K.; Mo, J.M. Different responses of asymbiotic nitrogen fixation to nitrogen addition between disturbed and rehabilitated subtropical forests. Sci. Total Environ. 2017, 601, 1505–1512. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.J.; Zhang, Y.; Han, W.X.; Tang, A.H.; Shen, J.L.; Cui, Z.L.; Vitousek, P.; Erisman, J.W.; Goulding, K.; Christie, P.; et al. Enhanced nitrogen deposition over China. Nature 2013, 494, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Yu, G.; He, N.; Zhan, X.; Fang, H.; Sheng, W.; Zuo, Y.; Zhang, D.; Wang, Q. Spatial and decadal variations in inorganic nitrogen wet deposition in China induced by human activity. Sci. Rep.-UK 2014, 4, 3763. [Google Scholar] [CrossRef]
- Stevens, C.J.; Dise, N.B.; Mountford, J.O.; Gowing, D.J. Impact of nitrogen deposition on the species richness of grasslands. Science 2013, 303, 1876–1879. [Google Scholar] [CrossRef]
- Elser, J.J.; Bracken, M.E.; Cleland, E.E.; Gruner, D.S.; Harpole, W.S.; Hillebrand, H.; Ngai, J.T.; Seabloom, E.W.; Shurin, J.B.; Smith, J.E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol. Lett. 2007, 10, 1135–1142. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.J.; Lei, P.F.; Xiang, W.H.; Ouyang, S.; Hui, X.R. Growth Variations of Tree Saplings in Relation to Species Diversity and Functional Traits in a Tree Diversity Pot Experiment. Forests 2018, 9, 380. [Google Scholar] [CrossRef]
- Gilliam, F.S.; May, J.D.; Adams, M.B. Response of foliar nutrients of Rubus allegheniensis to nutrient amendments in a central Appalachian hardwood forest. For. Ecol. Manag. 2018, 411, 101–107. [Google Scholar] [CrossRef]
- Lu, R.K. Analysis Methods of Soil Agricultural Chemistry; China Agricultural Science and Technology Press: Beijing, China, 1999; pp. 474–490. [Google Scholar]
- Reed, S.C.; Cleveland, C.C.; Townsend, A.R. Relationships among phosphorus, molybdenum and free-living nitrogen fixation in tropical rain forests: Results from observational and experimental analyses. Biogeochemistry 2013, 114, 135–147. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS Online Documentation, 8th ed.; SAS Institute Inc.: Cary, NC, USA, 2001. [Google Scholar]
- R Core Development Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2004. [Google Scholar]
- Bargali, K.; Bargali, S.S. Nutrient utilisation efficiencies of two central Himalayan tree species. J. Trop For. Sci. 2000, 12, 450–458. [Google Scholar]
- Singh, S.P.; Bargali, K.; Joshi, A.; Chaudhry, S. Nitrogen resorption in leaves of tree and shrub seedlings in response to increasing soil fertility. Curr. Sci. 2005, 89, 389–396. [Google Scholar]
- Manral, V.; Bargali, K.; Bargali, S.S.; Shahi, C. Changes in soil biochemical properties following replacement of Banj oak forest with Chir pine in Central Himalaya, India. Ecol. Process. 2020, 9, 30. [Google Scholar] [CrossRef]
- Brown, K.R.; Thompson, W.A.; Camm, E.L.; Hawkins, B.J.; Guy, R.D. Effects of N addition rates on the productivity of Picea sitchensis, Thuja plicata, and Tsuga heterophylla seedlings. 2. Photosynthesis, 13C discrimination and N partitioning in foliage. Trees-Struct. Funct. 1996, 10, 198–205. [Google Scholar] [CrossRef]
- Lu, X.; Mao, Q.; Mo, J.; Gilliam, F.S.; Zhou, G.; Luo, Y.; Zhang, W.; Huang, J. Divergent responses of soil buffering capacity to long-term N deposition in three typical tropical forests with different land-use history. Environ. Sci. Technol. 2015, 49, 4072–4080. [Google Scholar] [CrossRef]
- Lu, X.K.; Mao, Q.G.; Gilliam, F.S.; Luo, Y.Q.; Mo, J.M. Nitrogen deposition contributes to soil acidification in tropical ecosystems. Glob. Chang. Biol. 2014, 20, 3790–3801. [Google Scholar] [CrossRef]
- Tomlinson, G.H. Acidic deposition, nutrient leaching and forest growth. Biogeochemistry 2003, 65, 51–81. [Google Scholar] [CrossRef]
- Holmberg, M.; Aherne, J.; Austnes, K.; Beloica, J.; De Marco, A.; Dirnbock, T.; Fornasier, M.F.; Goergen, K.; Futter, M.; Lindroos, A.J.; et al. Modelling study of soil C, N and pH response to air pollution and climate change using European LTER site observations. Sci. Total Environ. 2018, 640–641, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Zarif, N.; Khan, A.; Wang, Q.C. Linking Soil Acidity to P Fractions and Exchangeable Base Cations under Increased N and P Fertilization of Mono and Mixed Plantations in Northeast China. Forests 2020, 11, 1274. [Google Scholar] [CrossRef]
- Rothwell, J.J.; Futter, M.N.; Dise, N.B. A classification and regression tree model of controls on dissolved inorganic nitrogen leaching from European forests. Environ. Pollut. 2008, 156, 544–552. [Google Scholar] [CrossRef]
- MacDonald, J.A.; Dise, N.B.; Matzner, E.; Armbruster, M.; Gundersen, P.; Forsius, M. Nitrogen input together with ecosystem nitrogen enrichment predict nitrate leaching from European forests. Glob. Chang. Biol. 2002, 8, 1028–1033. [Google Scholar] [CrossRef]
- Matschonat, G.; Matzner, E. Soil chemical properties affecting NH4+ sorption in forest soils. Z Pflanz Bodenkd. 1996, 159, 505–511. [Google Scholar] [CrossRef]
- Ai, Z.M.; Xue, S.; Wang, G.L.; Liu, G.B. Responses of Non-structural Carbohydrates and C:N:P Stoichiometry of Bothriochloa ischaemum to Nitrogen Addition on the Loess Plateau, China. J. Plant Growth Regul. 2017, 36, 714–722. [Google Scholar] [CrossRef]
- Eberwein, J.; Shen, W.J.; Jenerette, G.D. Michaelis-Menten kinetics of soil respiration feedbacks to nitrogen deposition and climate change in subtropical forests. Sci Rep.-UK 2017, 7, 1752. [Google Scholar] [CrossRef]
- Prajapati, K.; Modi, H.A. The importance of potassium in plant growth—A review. Indian J. Plant Sci. 2012, 1, 177–186. [Google Scholar]
- Pilon-Smits, E.A.; Quinn, C.F.; Tapken, W.; Malagoli, M.; Schiavon, M. Physiological functions of beneficial elements. Curr. Opin. Plant Biol. 2009, 12, 267–274. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treat | Diameter (cm) | Height (cm) | Root (g) | Stem (g) | Leaf (g) | Total Biomass (g) |
|---|---|---|---|---|---|---|
| CK | 4.13 ± 0.12a | 15.90 ± 0.34a | 5.69 ± 0.06a | 5.31 ± 0.05a | 5.37 ± 0.06a | 16.37 ± 0.13a |
| L1 | 4.64 ± 0.10b | 19.47 ± 0.29b | 7.34 ± 0.10b | 5.86 ± 0.06b | 6.09 ± 0.05b | 19.29 ± 0.15b |
| L2 | 5.62 ± 0.06c | 23.26 ± 0.38c | 7.67 ± 0.09b | 6.28 ± 0.08c | 6.20 ± 0.11b | 20.14 ± 0.15c |
| L3 | 4.88 ± 0.13b | 20.59 ± 0.37b | 5.89 ± 0.13a | 5.90 ± 0.07b | 5.76 ± 0.07c | 17.54 ± 0.21d |
| Variate | Fixed | Estimate | SE | t-Value | p-Value | AIC |
|---|---|---|---|---|---|---|
| Diameter | Intercept | 8.00 | 1.05 | 7.60 | <0.001 | −50.45 |
| K+ | −2.95 | 1.86 | −1.58 | 0.12 | ||
| Na+ | 9.23 | 4.33 | 2.13 | 0.04 | ||
| Mg2+ | −0.46 | 0.26 | −1.75 | 0.09 | ||
| Height | Intercept | 33.43 | 3.97 | 8.42 | <0.001 | 55.93 |
| K+ | −24.61 | 7.07 | −3.48 | 0.001 | ||
| Na+ | 44.55 | 16.03 | 2.78 | 0.01 | ||
| Mg2+ | −1.39 | 0.98 | −1.42 | 0.16 | ||
| Biomass | Intercept | 20.90 | 0.87 | 24.12 | <0.001 | 28.02 |
| K+ | −14.92 | 3.91 | −3.82 | <0.001 | ||
| Na+ | 26.89 | 8.30 | 3.24 | 0.003 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Wang, Y.; Zhang, Z.; Pan, H.; Lan, L.; Huang, R.; Deng, X.; Peng, Y. Effects of Exponential N Application on Soil Exchangeable Base Cations and the Growth and Nutrient Contents of Clonal Chinese Fir Seedlings. Plants 2023, 12, 851. https://doi.org/10.3390/plants12040851
Wang R, Wang Y, Zhang Z, Pan H, Lan L, Huang R, Deng X, Peng Y. Effects of Exponential N Application on Soil Exchangeable Base Cations and the Growth and Nutrient Contents of Clonal Chinese Fir Seedlings. Plants. 2023; 12(4):851. https://doi.org/10.3390/plants12040851
Chicago/Turabian StyleWang, Renjie, Yong Wang, Zeyao Zhang, Huibiao Pan, Liufeng Lan, Ronglin Huang, Xiaojun Deng, and Yuanying Peng. 2023. "Effects of Exponential N Application on Soil Exchangeable Base Cations and the Growth and Nutrient Contents of Clonal Chinese Fir Seedlings" Plants 12, no. 4: 851. https://doi.org/10.3390/plants12040851