
Soil Nutrients, Enzyme Activities, and Microbial Communities along a Chronosequence of Chinese Fir Plantations in Subtropical China
 , ,
, ,
Abstract
:1. Introduction
2. Results
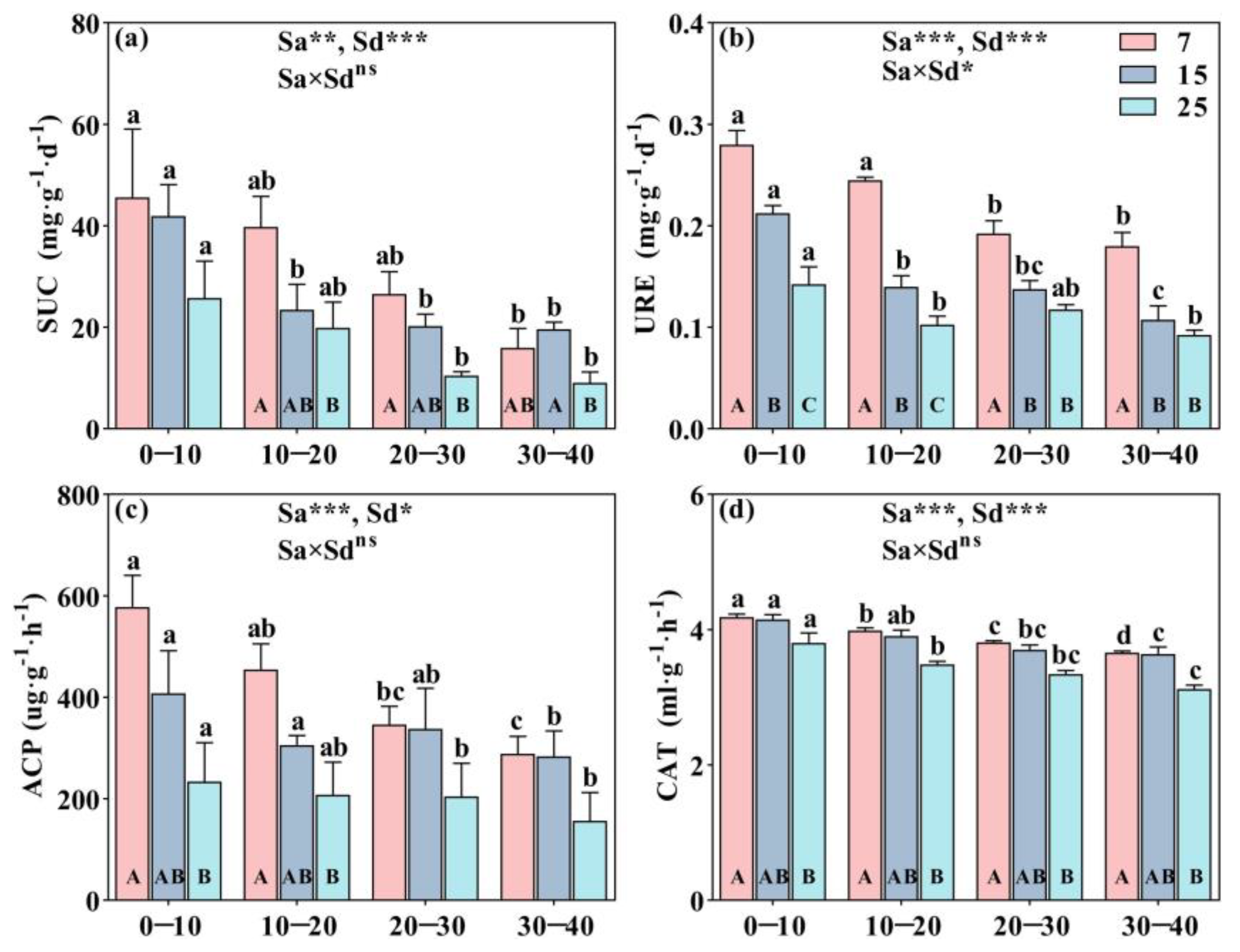
2.1. The Characteristics of Soil Physicochemical Properties, Microbial Biomass, and Enzyme Activities in Stand Ages and Soil Depth
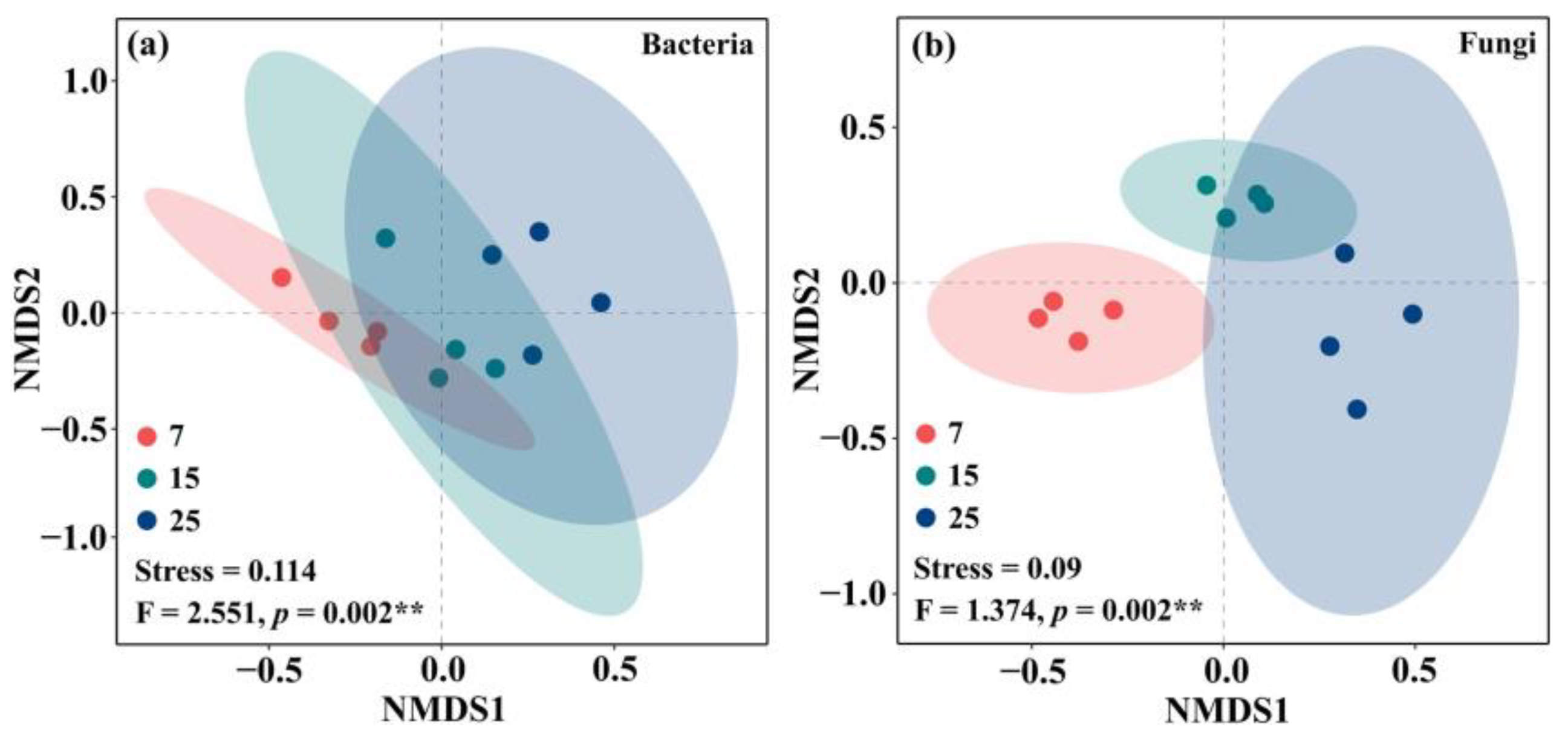
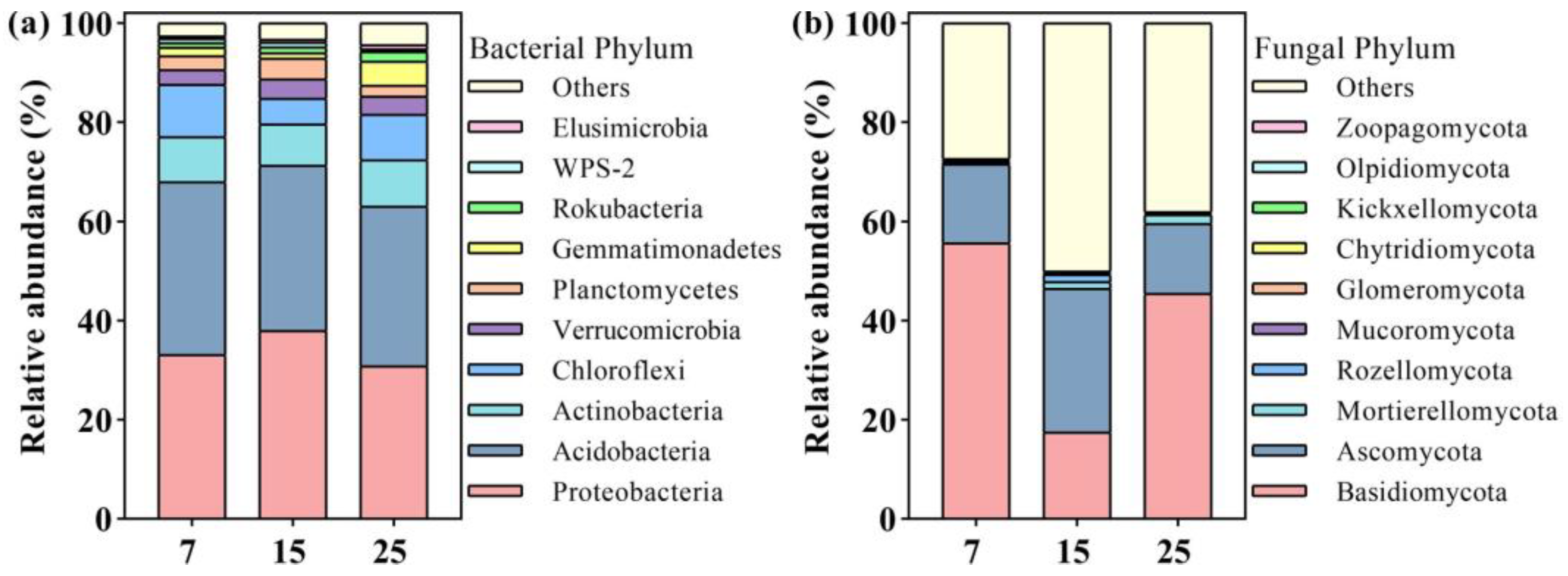
2.2. Soil Microbial Diversity and Community Composition
2.3. Effects of Soil Abiotic and Biotic Factors on Soil Microbial Community
3. Materials and Methods
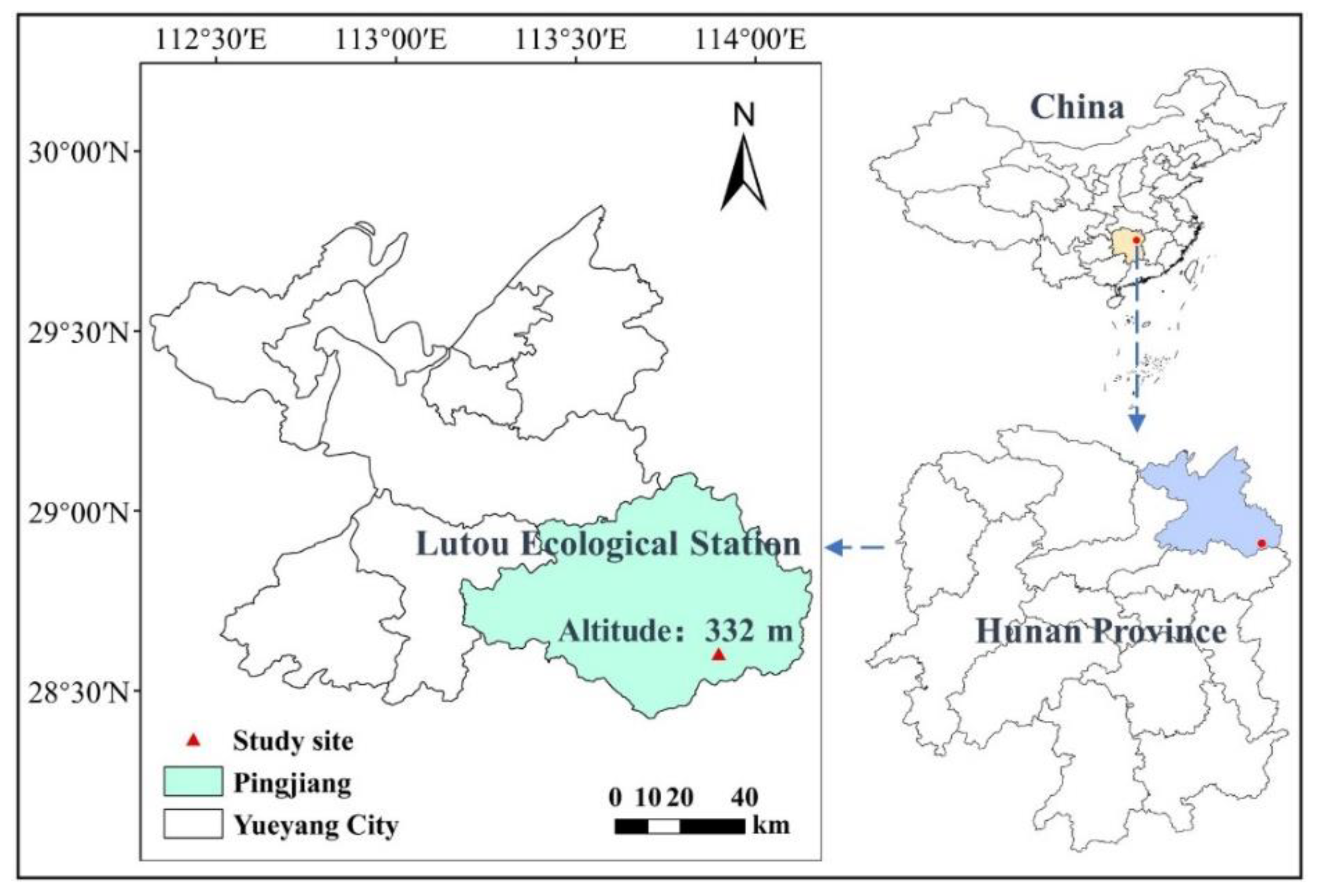
3.1. Study Site
3.2. Experimental Design and Soil Sampling
3.3. Soil Sample Analysis
3.4. DNA Extraction and High-Throughput Sequencing
3.5. Statistical Analysis
4. Discussion
4.1. Variation of Soil Nutrients and Enzyme Activities in Different Stand Ages of CFP
4.2. Variations of Soil Microbial Diversity and Composition in Different Stand Ages of CFP
4.3. Effect of Soil Environmental Factors on Microbial Communities in CFP
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Colombo, F.; Macdonald, C.A.; Jeffries, T.C.; Powell, J.R.; Singh, B.K. Impact of forest management practices on soil bacterial diversity and consequences for soil processes. Soil Biol. Biochem. 2016, 94, 200–210. [Google Scholar] [CrossRef]
- Record, S.; Kobe, R.K.; Vriesendorp, C.F.; Finley, A.O. Seedling survival responses to conspecific density, soil nutrients, and irradiance vary with age in a tropical forest. Ecology 2016, 97, 2406–2415. [Google Scholar] [CrossRef] [PubMed]
- Cline, L.C.; Zak, D.R. Soil microbial communities are shaped by plant-driven changes in resource availability during secondary succession. Ecology 2015, 96, 3374–3385. [Google Scholar] [CrossRef] [PubMed]
- Bright, R.M.; Antón-Fernández, C.; Astrup, R.; Cherubini, F.; Kvalevåg, M.; Strømman, A.H. Climate change implications of shifting forest management strategy in a boreal forest ecosystem of Norway. Glob. Chang. Biol. 2014, 20, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Zhao, N.; Liu, C.C.; Yang, H.; Li, M.L.; Yu, G.R.; Wilcox, K.; Yu, Q.; He, N.P. C:N:P stoichiometry in China’s forests: From organs to ecosystems. Funct. Ecol. 2018, 32, 50–60. [Google Scholar] [CrossRef]
- Brown, S.P.; Jumpponen, A. Phylogenetic diversity analyses reveal disparity between fungal and bacterial communities during microbial primary succession. Soil Biol. Biochem. 2015, 89, 52–60. [Google Scholar] [CrossRef]
- Shen, F.; Wu, J.; Fan, H.; Liu, W.; Guo, X.; Duan, H.; Hu, L.; Lei, X.; Wei, X. Soil N/P and C/P ratio regulate the responses of soil microbial community composition and enzyme activities in a long-term nitrogen loaded Chinese fir forest. Plant Soil 2019, 436, 91–107. [Google Scholar] [CrossRef]
- Nottingham, A.T.; Whitaker, J.; Ostle, N.J.; Bardgett, R.D.; McNamara, N.P.; Fierer, N.; Salinas, N.; Ccahuana, A.J.Q.; Turner, B.L.; Meir, P. Microbial responses to warming enhance soil carbon loss following translocation across a tropical forest elevation gradient. Ecol. Lett. 2019, 22, 1889–1899. [Google Scholar] [CrossRef]
- Zhang, M.; O’Connor, P.J.; Zhang, J.; Ye, X. Linking soil nutrient cycling and microbial community with vegetation cover in riparian zone. Geoderma 2021, 384, 114801. [Google Scholar] [CrossRef]
- Bergmann, J.; Verbruggen, E.; Heinze, J.; Xiang, D.; Chen, B.; Joshi, J.; Rillig, M.C. The interplay between soil structure, roots, and microbiota as a determinant of plant-soil feedback. Ecol. Evol. 2016, 6, 7633–7644. [Google Scholar] [CrossRef]
- Semchenko, M.; Barry, K.E.; de Vries, F.T.; Mommer, L.; Moora, M.; Maciá-Vicente, J.G. Deciphering the role of specialist and generalist plant–microbial interactions as drivers of plant–soil feedback. New Phytol. 2022, 234, 1929–1944. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xue, L.; Jiao, R. Soil organic carbon fractions, C-cycling associated hydrolytic enzymes, and microbial carbon metabolism vary with stand age in Cunninghamia lanceolate (Lamb.) Hook plantations. For. Ecol. Manag. 2021, 482, 118887. [Google Scholar] [CrossRef]
- Guo, J.; Feng, H.; Roberge, G.; Feng, L.; Pan, C.; McNie, P.; Yu, Y. The negative effect of Chinese fir (Cunninghamia lanceolata) monoculture plantations on soil physicochemical properties, microbial biomass, fungal communities, and enzymatic activities. For. Ecol. Manag. 2022, 519, 120297. [Google Scholar] [CrossRef]
- Yang, Z.; Chen, S.; Liu, X.; Xiong, D.; Xu, C.; Arthur, M.A.; McCulley, R.L.; Shi, S.; Yang, Y. Loss of soil organic carbon following natural forest conversion to Chinese fir plantation. For. Ecol. Manag. 2019, 449, 117476. [Google Scholar] [CrossRef]
- Guillaume, T.; Kotowska, M.M.; Hertel, D.; Knohl, A.; Krashevska, V.; Murtilaksono, K.; Scheu, S.; Kuzyakov, Y. Carbon costs and benefits of Indonesian rainforest conversion to plantations. Nat. Commun. 2018, 9, 2388. [Google Scholar] [CrossRef]
- Kumar, S.; Garkoti, S.C. Rhizosphere influence on soil microbial biomass and enzyme activity in banj oak, chir pine and banj oak regeneration forests in the central Himalaya. Geoderma 2022, 409, 115626. [Google Scholar] [CrossRef]
- Ren, C.; Wang, J.; Bastida, F.; Delgado-Baquerizo, M.; Yang, Y.; Wang, J.; Zhong, Z.; Zhou, Z.; Zhang, S.; Guo, Y.; et al. Microbial traits determine soil C emission in response to fresh carbon inputs in forests across biomes. Glob. Chang. Biol. 2022, 28, 1516–1528. [Google Scholar] [CrossRef]
- Ananbeh, H.; Stojanovic, M.; Pompeiano, A.; Voberkova, S.; Trasar-Cepeda, C. Use of soil enzyme activities to assess the recovery of soil functions in abandoned coppice forest systems. Sci. Total Environ. 2019, 694, 133692. [Google Scholar] [CrossRef]
- Liu, X.; Wang, Y.; Liu, Y.; Chen, H.; Hu, Y. Response of Bacterial and Fungal Soil Communities to Chinese Fir (Cunninghamia lanceolate) Long-Term Monoculture Plantations. Front. Microbiol. 2020, 11, 181. [Google Scholar] [CrossRef]
- Sun, S.; Li, S.; Avera, B.N.; Strahm, B.D.; Badgley, B.D. Soil Bacterial and Fungal Communities Show Distinct Recovery Patterns during Forest Ecosystem Restoration. Appl. Environ. Microbiol. 2017, 83, e00966-17. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, G.; Zhu, P.; Xing, S.; Wang, C. Soil C, N and P contents and their stoichiometry as affected by typical plant communities on steep gully slopes of the Loess Plateau, China. Catena 2022, 208, 105740. [Google Scholar] [CrossRef]
- Ortiz, C.; Fernández-Alonso, M.J.; Kitzler, B.; Díaz-Pinés, E.; Saiz, G.; Rubio, A.; Benito, M. Variations in soil aggregation, microbial community structure and soil organic matter cycling associated to long-term afforestation and woody encroachment in a Mediterranean alpine ecotone. Geoderma 2022, 405, 115450. [Google Scholar] [CrossRef]
- Yang, M.; Yang, D.; Yu, X. Soil microbial communities and enzyme activities in sea-buckthorn (Hippophae rhamnoides) plantation at different ages. PLoS ONE 2018, 13, e0190959. [Google Scholar] [CrossRef] [PubMed]
- He, X.X.; Huang, Y.Z.; Zhang, Q.C.; Ye, S.M.; Wang, S.Q. Distribution of organic carbon fractions in soil aggregates in Chinese fir plantations with different stand ages. Ecol. Process. 2021, 10, 49. [Google Scholar] [CrossRef]
- Xu, M.; Lu, X.; Xu, Y.; Zhong, Z.; Zhang, W.; Ren, C.; Han, X.; Yang, G.; Feng, Y. Dynamics of bacterial community in litter and soil along a chronosequence of Robinia pseudoacacia plantations. Sci. Total Environ. 2020, 703, 135613. [Google Scholar] [CrossRef]
- Qiao, Y.; Wang, J.; Liu, H.; Huang, K.; Yang, Q.; Lu, R.; Yan, L.; Wang, X.; Xia, J. Depth-dependent soil C-N-P stoichiometry in a mature subtropical broadleaf forest. Geoderma 2020, 370, 114357. [Google Scholar] [CrossRef]
- Li, J.; Sang, C.; Yang, J.; Qu, L.; Xia, Z.; Sun, H.; Jiang, P.; Wang, X.; He, H.; Wang, C. Stoichiometric imbalance and microbial community regulate microbial elements use efficiencies under nitrogen addition. Soil Biol. Biochem. 2021, 156, 108207. [Google Scholar] [CrossRef]
- Yu, Z.; Guoyi, Z.; Liu, L.; Manzoni, S.; Ciais, P.; Goll, D.; Peñuelas, J.; Sardans, J.; Wang, W.; Zhu, J.; et al. Natural forests promote phosphorus retention in soil. Glob. Chang. Biol. 2022, 28, 1678–1689. [Google Scholar] [CrossRef]
- Cenini, V.L.; Fornara, D.A.; McMullan, G.; Ternan, N.; Carolan, R.; Crawley, M.J.; Clément, J.-C.; Lavorel, S. Linkages between extracellular enzyme activities and the carbon and nitrogen content of grassland soils. Soil Biol. Biochem. 2016, 96, 198–206. [Google Scholar] [CrossRef]
- Prescott, C.E.; Grayston, S.J. Tree species influence on microbial communities in litter and soil: Current knowledge and research needs. For. Ecol. Manag. 2013, 309, 19–27. [Google Scholar] [CrossRef]
- Urbanova, M.; Snajdr, J.; Baldrian, P. Composition of fungal and bacterial communities in forest litter and soil is largely determined by dominant trees. Soil Biol. Biochem. 2015, 84, 53–64. [Google Scholar] [CrossRef]
- Waymouth, V.; Miller, R.E.; Ede, F.; Bissett, A.; Aponte, C. Variation in soil microbial communities: Elucidating relationships with vegetation and soil properties, and testing sampling effectiveness. Plant Ecol. 2020, 221, 837–851. [Google Scholar] [CrossRef]
- Toju, H.; Kishida, O.; Katayama, N.; Takagi, K. Networks Depicting the Fine-Scale Co-Occurrences of Fungi in Soil Horizons. PLoS ONE 2016, 11, e0165987. [Google Scholar] [CrossRef] [PubMed]
- Bayranvand, M.; Akbarinia, M.; Jouzani, G.S.; Gharechahi, J.; Kooch, Y.; Baldrian, P. Composition of soil bacterial and fungal communities in relation to vegetation composition and soil characteristics along an altitudinal gradient. FEMS Microbiol. Ecol. 2021, 97, fiaa201. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Xing, Y.; Liu, G.; Hu, C.; Wang, X.; Yan, G.; Wang, Q. Changes in soil bacterial and fungal community composition and functional groups during the succession of boreal forests. Soil Biol. Biochem. 2021, 161, 108393. [Google Scholar] [CrossRef]
- Wang, J.; Shi, X.; Zheng, C.; Suter, H.; Huang, Z. Different responses of soil bacterial and fungal communities to nitrogen deposition in a subtropical forest. Sci. Total Environ. 2021, 755, 142449. [Google Scholar] [CrossRef]
- Rodriguez-Ramos, J.C.; Cale, J.A.; Cahill, J.F.; Simard, S.W.; Karst, J.; Erbilgin, N. Changes in soil fungal community composition depend on functional group and forest disturbance type. New Phytol. 2021, 229, 1105–1117. [Google Scholar] [CrossRef]
- Singh, J.S.; Gupta, V.K. Soil microbial biomass: A key soil driver in management of ecosystem functioning. Sci. Total Environ. 2018, 634, 497–500. [Google Scholar] [CrossRef]
- Wardle, D.A.; Gundale, M.J.; Kardol, P.; Nilsson, M.; Fanin, N. Impact of plant functional group and species removals on soil and plant nitrogen and phosphorus across a retrogressive chronosequence. J. Ecol. 2020, 108, 561–573. [Google Scholar] [CrossRef]
- Liu, J.L.; Wang, Q.Q.; Ku, Y.L.; Zhang, W.W.; Zhu, H.L.; Zhao, Z. Precipitation and soil pH drive the soil microbial spatial patterns in the Robinia pseudoacacia forests at the regional scale. Catena 2022, 212, 106120. [Google Scholar] [CrossRef]
- Manral, V.; Bargali, K.; Bargali, S.S.; Jhariya, M.K.; Padalia, K. Relationships between soil and microbial biomass properties and annual flux of nutrients in Central Himalaya forests, India. Land Degrad. Dev. 2022, 33, 2014–2025. [Google Scholar] [CrossRef]
- Wang, C.; Liu, D.; Bai, E. Decreasing soil microbial diversity is associated with decreasing microbial biomass under nitrogen addition. Soil Biol. Biochem. 2018, 120, 126–133. [Google Scholar] [CrossRef]
- Grant, T.; Sethuraman, A.; Escobar, M.A.; Vourlitis, G.L. Chronic dry nitrogen inputs alter soil microbial community composition in Southern California semi-arid shrublands. Appl. Soil Ecol. 2022, 176, 104496. [Google Scholar] [CrossRef]
- Cai, H.; Li, F.; Jin, G. Soil nutrients, forest structure and species traits drive aboveground carbon dynamics in an old-growth temperate forest. Sci. Total Environ. 2020, 705, 135874. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stand Age (Year) | 7 | 15 | 25 |
|---|---|---|---|
| Overstory | |||
| Mean DBH (cm) | 6.5 | 17.3 | 20.8 |
| Mean height (m) | 4.5 | 16.3 | 18 |
| Canopy closure (%) | 0.5 | 0.7 | 0.85 |
| Shrubs coverage (%) | 10% | 28% | 70% |
| Herbs coverage (%) | 82% | 60% | 75% |
| Slope aspect | WS | ES | WS |
| Gradient (°) | 29.6 | 29.2 | 45.2 |
| Altitude (m) | 438.4 | 281.3 | 278 |
| Soil properties | |||
| SWC (%) | 28.63 ± 0.96 A | 21.50 ± 1.91 B | 20.17 ± 1.93 B |
| SBD (g/cm3) | 1.08 ± 0.08 | 1.32 ± 0.04 | 1.28 ± 0.03 |
| pH | 4.96 ± 0.01 A | 4.67 ± 0.04 B | 4.66 ± 0.04 B |
| SOC (g/kg) | 15.71 ± 1.28 A | 9.11 ± 0.93 B | 11.59 ± 0.83 B |
| TN (g/kg) | 1.96 ± 0.13 | 1.58 ± 0.10 | 1.70 ± 0.06 |
| TP (g/kg) | 0.39 ± 0.03 A | 0.27 ± 0.01 B | 0.34 ± 0.01 A |
| C/N | 7.86 ± 0.29 A | 5.55 ± 0.27 B | 6.48 ± 0.35 B |
| C/P | 39.84 ± 1.52 | 33.59 ± 2.95 | 33.91 ± 2.79 |
| N/P | 5.01 ± 0.19 | 5.85 ± 0.28 | 5.06 ± 0.26 |
| MBC (mg/kg) | 505.52 ± 30.04 A | 330.03 ± 15.60 B | 409.79 ± 40.60 AB |
| MBN (mg/kg) | 190.44 ± 7.76 A | 124.08 ± 9.77 B | 101.74 ± 7.60 B |
| MBP (mg/kg) | 7.27 ± 0.83 | 4.66 ± 0.55 | 5.12 ± 0.73 |
| MBC/MBN | 3.05 ± 0.19 B | 2.80 ± 0.23 B | 4.43 ± 0.46 A |
| MBC/MBP | 99.03 ± 15.78 | 77.43 ± 9.96 | 111.08 ± 18.54 |
| MBN/MBP | 31.59 ± 3.56 | 28.96 ± 3.64 | 25.35 ± 1.34 |
| MBC/MBN | 3.05 ± 0.19 B | 2.80 ± 0.23 B | 4.43 ± 0.46 A |
| MBC/MBP | 99.03 ± 15.78 | 77.43 ± 9.96 | 111.08 ± 18.54 |
| MBN/MBP | 31.59 ± 3.56 | 28.96 ± 3.64 | 25.35 ± 1.34 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, J.; Cao, Y.; Wang, J.; Chen, Y.; Peng, Y.; Shao, Q.; Dan, Q.; Xu, Y.; Chen, X.; Dang, P.; et al. Soil Nutrients, Enzyme Activities, and Microbial Communities along a Chronosequence of Chinese Fir Plantations in Subtropical China. Plants 2023, 12, 1931. https://doi.org/10.3390/plants12101931
Lei J, Cao Y, Wang J, Chen Y, Peng Y, Shao Q, Dan Q, Xu Y, Chen X, Dang P, et al. Soil Nutrients, Enzyme Activities, and Microbial Communities along a Chronosequence of Chinese Fir Plantations in Subtropical China. Plants. 2023; 12(10):1931. https://doi.org/10.3390/plants12101931
Chicago/Turabian StyleLei, Junjie, Yixuan Cao, Jun Wang, Yazhen Chen, Yuanying Peng, Qiwen Shao, Qing Dan, Yichen Xu, Xiaoyong Chen, Peng Dang, and et al. 2023. "Soil Nutrients, Enzyme Activities, and Microbial Communities along a Chronosequence of Chinese Fir Plantations in Subtropical China" Plants 12, no. 10: 1931. https://doi.org/10.3390/plants12101931