
CuO Nanoparticle-Mediated Seed Priming Improves Physio-Biochemical and Enzymatic Activities of Brassica juncea

 ,
,  ,
,  , ,
, ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Phenotypic Character
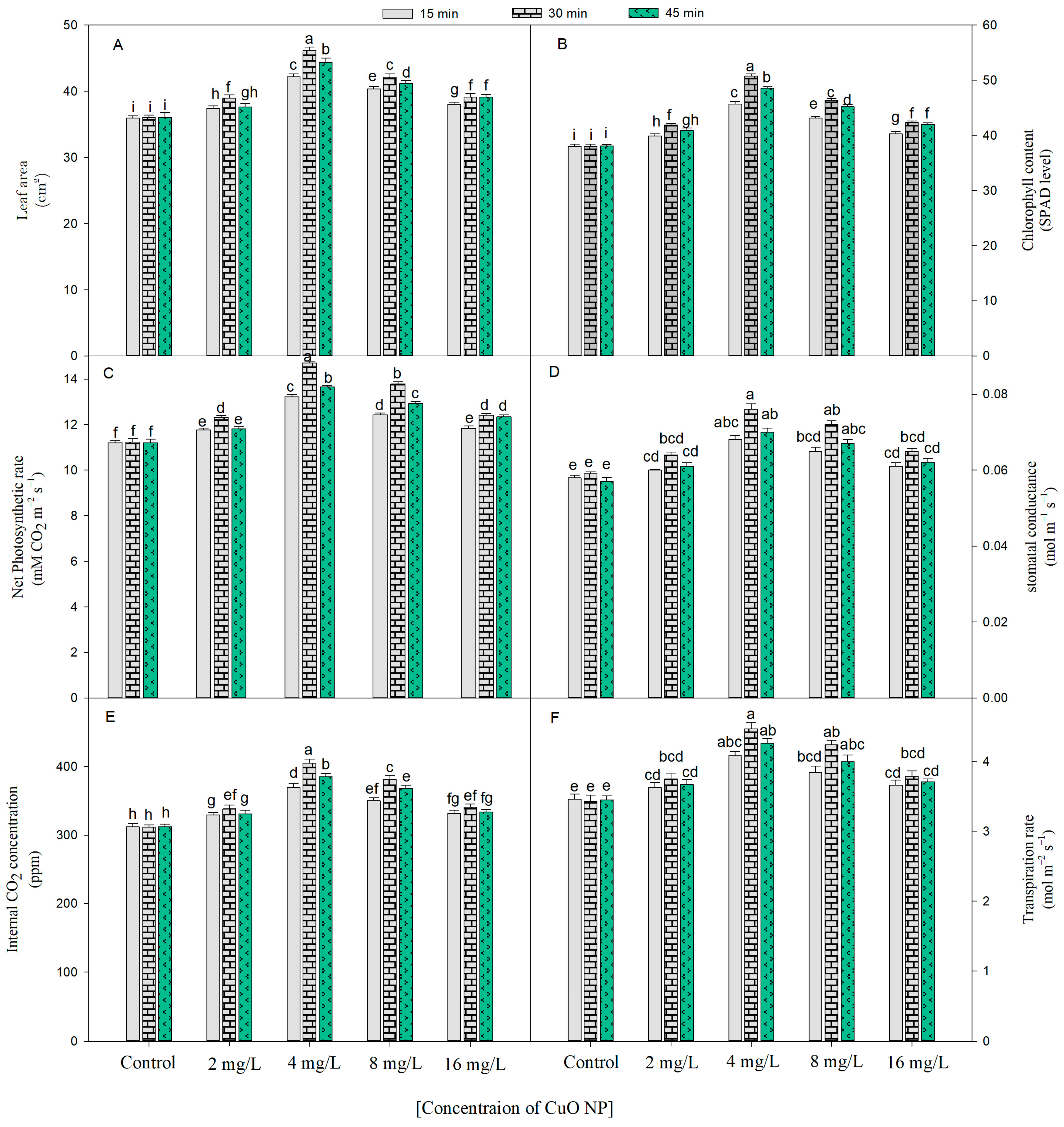
2.2. Effect of CuO NPs on Physiological Indices
2.2.1. Chlorophyll Content (SPAD Value)
2.2.2. Photosynthetic Parameters
2.3. Effects of CuO NPs on Biochemical Parameters
2.3.1. Activity of Carbonic Anhydrase (CA) and Nitrate Reductase (NR)
2.3.2. Enzymatic Activity
2.3.3. Proline Content
3. Discussion
4. Materials and Methods
4.1. Nanoparticles and Seed Priming
4.2. Experimental Setup and Plant Growth Performance
4.3. Determination of Phenotypic Characteristics
4.4. Observation of Physiological Indices
4.4.1. Leaf Area and Chlorophyll Content (SPAD Value)
4.4.2. Photosynthetic Rate and Their Related Parameters
4.5. Biochemical Parameters Estimation
4.5.1. NR and CA Activity
4.5.2. Estimation of Antioxidant Enzymes
4.5.3. Content of Proline
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Singh, P.; Kim, Y.; Zhang, D.; Yang, D. Biological Synthesis of Nanoparticles from Plants and Microorganisms. Trends Biotechnol. 2016, 34, 588–599. [Google Scholar] [CrossRef]
- Bernhardt, E.S.; Colman, B.P.; Hochella, M.F.; Cardinale, B.J.; Nisbet, R.M.; Richardson, C.J.; Yin, L. An Ecological Perspective on Nanomaterial Impacts in the Environment. J. Environ. Qual. 2010, 39, 1954. [Google Scholar] [CrossRef]
- Rajput, V.D.; Singh, A.; Minkina, T.; Rawat, S.; Mandzhieva, S.; Sushkova, S.; Shuvaeva, V.; Nazarenko, O.; Rajput, P.; Verma, K.K. Nano-Enabled Products: Challenges and Opportunities for Sustainable Agriculture. Plants 2021, 10, 2727. [Google Scholar] [CrossRef]
- Sashidhar, P.; Arya, S.; Das, R.K.; Dubey, M.K.; Lenka, S.K. Nanobiotechnology for Plant Genome Engineering and Crop Protection. In Genetically Modified Crops in Asia Pacific; CSIRO Australia: Canberra, Autralia, 2021; pp. 279–310. [Google Scholar]
- Aleksandrowicz-Trzcińska, M.; Olchowik, J.; Studnicki, M.; Urban, A. Do Silver Nanoparticles Stimulate the Formation of Ectomycorrhizae in Seedlings of Pedunculate Oak (Quercus Robur L.)? Symbiosis 2019, 79, 89–97. [Google Scholar] [CrossRef]
- Arya, S.; Rookes, J.E.; Cahill, D.M.; Lenka, S.K. Reduced Genotoxicity of Gold Nanoparticles With Protein Corona in Allium Cepa. Front. Bioeng. Biotechnol. 2022, 10, 849464. [Google Scholar] [CrossRef]
- Arya, S.; Tanwar, N.; Lenka, S.K. Prospects of Nano-and Peptide-Carriers to Deliver CRISPR Cargos in Plants to Edit across and beyond Central Dogma. Nanotechnol. Environ. Eng. 2021, 6, 22. [Google Scholar] [CrossRef]
- Croteau, M.-N.; Misra, S.K.; Luoma, S.N.; Valsami-Jones, E. Bioaccumulation and Toxicity of CuO Nanoparticles by a Freshwater Invertebrate after Waterborne and Dietborne Exposures. Environ. Sci. Technol. 2014, 48, 10929–10937. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Han, H.; Liu, X.; Gu, X.; Chen, K.; Lu, D. Multi-Walled Carbon Nanotubes Can Enhance Root Elongation of Wheat (Triticum Aestivum) Plants. J. Nanoparticle Res. 2012, 14, 841. [Google Scholar] [CrossRef]
- Xiong, T.; Dumat, C.; Dappe, V.; Vezin, H.; Schreck, E.; Shahid, M.; Pierart, A.; Sobanska, S. Copper Oxide Nanoparticle Foliar Uptake, Phytotoxicity, and Consequences for Sustainable Urban Agriculture. Environ. Sci. Technol. 2017, 51, 5242–5251. [Google Scholar] [CrossRef] [PubMed]
- Rehman, R.U.; Khan, B.; Aziz, T.; Gul, F.Z.; Nasreen, S.; Zia, M. Postponement Growth and Antioxidative Response of Brassica Nigra on CuO and ZnO Nanoparticles Exposure under Soil Conditions. IET Nanobiotechnol. 2020, 14, 423–427. [Google Scholar] [PubMed]
- Wang, Y.; Lin, Y.; Xu, Y.; Yin, Y.; Guo, H.; Du, W. Divergence in Response of Lettuce (Var. Ramosa Hort.) to Copper Oxide Nanoparticles/Microparticles as Potential Agricultural Fertilizer. Environ. Pollut. Bioavailab. 2019, 31, 80–84. [Google Scholar] [CrossRef]
- Shende, S.; Rathod, D.; Gade, A.; Rai, M. Biogenic Copper Nanoparticles Promote the Growth of Pigeon Pea (Cajanus Cajan L.). IET Nanobiotechnol. 2017, 11, 773–781. [Google Scholar] [CrossRef]
- Zafar, H.; Ali, A.; Zia, M. CuO Nanoparticles Inhibited Root Growth from Brassica Nigra Seedlings but Induced Root from Stem and Leaf Explants. Appl. Biochem. Biotechnol. 2017, 181, 365–378. [Google Scholar] [CrossRef] [PubMed]
- Adhikari, T.; Kundu, S.; Biswas, A.K.; Tarafdar, J.C.; Rao, A.S. Effect of Copper Oxide Nano Particle on Seed Germination of Selected Crops. J. Agric. Sci. Technol. 2012, 2, 815–823. [Google Scholar]
- Keller, A.A.; Adeleye, A.S.; Conway, J.R.; Garner, K.L.; Zhao, L.; Cherr, G.N.; Hong, J.; Gardea-Torresdey, J.L.; Godwin, H.A.; Hanna, S. Comparative Environmental Fate and Toxicity of Copper Nanomaterials. NanoImpact 2017, 7, 28–40. [Google Scholar] [CrossRef]
- Sonawane, H.; Arya, S.; Math, S.; Shelke, D. Myco-Synthesized Silver and Titanium Oxide Nanoparticles as Seed Priming Agents to Promote Seed Germination and Seedling Growth of Solanum Lycopersicum: A Comparative Study. Int. Nano Lett. 2021, 11, 371–379. [Google Scholar] [CrossRef]
- An, J.; Hu, P.; Li, F.; Wu, H.; Shen, Y.; White, J.C.; Tian, X.; Li, Z.; Giraldo, J.P. Emerging Investigator Series: Molecular Mechanisms of Plant Salinity Stress Tolerance Improvement by Seed Priming with Cerium Oxide Nanoparticles. Environ. Sci. Nano 2020, 7, 2214–2228. [Google Scholar] [CrossRef]
- Munir, T.; Rizwan, M.; Kashif, M.; Shahzad, A.; Ali, S.; Amin, N.; Zahid, R.; Alam, M.F.E.; Imran, M. Effect of Zinc Oxide Nanoparticles on the Growth and Zn Uptake in Wheat (Triticum aestivum L.) by seed priming method. Dig. J. Nanomater. Biostruct. 2018, 13, 315–323. [Google Scholar]
- Salam, A.; Khan, A.R.; Liu, L.; Yang, S.; Azhar, W.; Ulhassan, Z.; Zeeshan, M.; Wu, J.; Fan, X.; Gan, Y. Seed Priming with Zinc Oxide Nanoparticles Downplayed Ultrastructural Damage and Improved Photosynthetic Apparatus in Maize under Cobalt Stress. J. Hazard. Mater. 2022, 423, 127021. [Google Scholar] [CrossRef]
- Ramkumar, T.R.; Arya, S.; Kumari, D.D.; Lenka, S.K. Brassica Juncea Genome Sequencing: Structural and Functional Insights. In The Brassica Juncea Genome; Springer: Berlin/Heidelberg, Germany, 2022; pp. 221–240. [Google Scholar]
- Kumar, P.R. Rapeseed Mustard Research in India: 21st Century Strategies. In Proceedings of the 10th International Rapeseed Congress, Canberra, Australia, 26–29 September 1999. [Google Scholar]
- Singh, D.; Arya, R.K.; Chandra, N.; Niwas, R.; Salisbury, P. Genetic Diversity Studies in Relation to Seed Yield and Its Component Traits in Indian Mustard (Brassica Juncea L. Czern & Coss.). J. Oilseed Brassica 2010, 1, 19–22. [Google Scholar]
- Zheng, L.; Hong, F.; Lu, S.; Liu, C. Effect of Nano-TiO2 on Strength of Naturally Aged Seeds and Growth of Spinach. Biol. Trace Elem. Res. 2005, 104, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Mondal, A.; Basu, R.; Das, S.; Nandy, P. Beneficial Role of Carbon Nanotubes on Mustard Plant Growth: An Agricultural Prospect. J. Nanoparticle Res. 2011, 13, 4519. [Google Scholar] [CrossRef]
- Dhoke, S.K.; Mahajan, P.; Kamble, R.; Khanna, A. Effect of Nanoparticles Suspension on the Growth of Mung (Vigna Radiata) Seedlings by Foliar Spray Method. Nanotechnol. Dev. 2013, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Burman, U.; Saini, M.; Kumar, P. Effect of Zinc Oxide Nanoparticles on Growth and Antioxidant System of Chickpea Seedlings. Toxicol. Environ. Chem. 2013, 95, 605–612. [Google Scholar] [CrossRef]
- Razzaq, A.; Ammara, R.; Jhanzab, H.M.; Mahmood, T.; Hafeez, A.; Hussain, S. A Novel Nanomaterial to Enhance Growth and Yield of Wheat. J. Nanosci. Technol. 2016, 2, 55–58. [Google Scholar]
- Jasim, B.; Thomas, R.; Mathew, J.; Radhakrishnan, E.K. Plant Growth and Diosgenin Enhancement Effect of Silver Nanoparticles in Fenugreek (Trigonella Foenum-Graecum L.). Saudi Pharm. J. 2016, 25, 4–8. [Google Scholar] [CrossRef]
- Fathi, A.; Zahedi, M.; Torabian, S.; Khoshgoftar, A. Response of Wheat Genotypes to Foliar Spray of ZnO and Fe2O3 Nanoparticles under Salt Stress. J. Plant Nutr. 2017, 40, 1376–1385. [Google Scholar] [CrossRef]
- Rajak, J.; Bawaskar, M.; Rathod, D.; Agarkar, G.; Nagaonkar, D.; Gade, A.; Rai, M. Interaction of Copper Nanoparticles and an Endophytic Growth Promoter Piriformospora Indica with Cajanus Cajan. J. Sci. Food Agric. 2017, 97, 4562–4570. [Google Scholar] [CrossRef]
- Liu, H.; Ma, C.; Chen, G.; White, J.C.; Wang, Z.; Xing, B.; Dhankher, O.P. Titanium Dioxide Nanoparticles Alleviate Tetracycline Toxicity to Arabidopsis Thaliana (L.). ACS Sustain. Chem. Eng. 2017, 5, 3204–3213. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E.; Møller, I.M.; Murphy, A. Plant Physiology and Development; Sinauer Associates Incorporated: Sunderland, MA, USA, 2015; ISBN 1605353531. [Google Scholar]
- Abdel Latef, A.A.H.; Srivastava, A.K.; El-sadek, M.S.A.; Kordrostami, M.; Tran, L.P. Titanium Dioxide Nanoparticles Improve Growth and Enhance Tolerance of Broad Bean Plants under Saline Soil Conditions. Land Degrad. Dev. 2017, 1073, 1065–1073. [Google Scholar] [CrossRef]
- Faizan, M.; Faraz, A.; Yusuf, M.; Khan, S.T.; Hayat, S. Zinc Oxide Nanoparticle-Mediated Changes in Photosynthetic Efficiency and Antioxidant System of Tomato Plants. Photosynthetica 2018, 56, 678–686. [Google Scholar] [CrossRef]
- Faraz, A.; Faizan, M.; Hayat, S. Effects of Copper Oxide Nanoparticles on the Photosynthesis and Antioxidant Levels of Mustard Plants (Brassica Juncea). J. Biol. Chem. Res. 2018, 35, 418–426. [Google Scholar]
- Lei, Z.; Mingyu, S.; Chao, L.; Liang, C.; Hao, H.; Xiao, W.; Xiaoqing, L.; Fan, Y.; Fengqing, G.; Fashui, H. Effects of Nanoanatase TiO2 on Photosynthesis of Spinach Chloroplasts under Different Light Illumination. Biol. Trace Elem. Res. 2007, 119, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Xu, G.; Qian, H.; Liu, P.; Zhao, P.; Hu, Y. Effects of Nano-TiO2 on Photosynthetic Characteristics of Ulmus Elongata Seedlings. Environ. Pollut. 2013, 176, 63–70. [Google Scholar] [CrossRef]
- Hong, F.; Zhou, J.; Liu, C.; Yang, F.; Wu, C.; Zheng, L.; Yang, P. Effect of Nano-TiO2 on Photochemical Reaction of Chloroplasts of Spinach. Biol. Trace Elem. Res. 2005, 105, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Li, B.; Zhang, Q.; Zhang, C. Effects of Nano-Silicon Dioxide on Photosynthetic Fluorescence Characteristics of Indocalamus Barbatus McClure. J. Nanjing For. Univ. 2012, 2, 59–63. [Google Scholar]
- Giraldo, J.P.; Landry, M.P.; Faltermeier, S.M.; McNicholas, T.P.; Iverson, N.M.; Boghossian, A.A.; Reuel, N.F.; Hilmer, A.J.; Sen, F.; Brew, J.A.; et al. Plant Nanobionics Approach to Augment Photosynthesis and Biochemical Sensing. Nat. Mater. 2014, 13, 400–408. [Google Scholar] [CrossRef]
- Singh, A.; Singh, N.B.; Hussain, I.; Singh, H. Effect of Biologically Synthesized Copper Oxide Nanoparticles on Metabolism and Antioxidant Activity to the Crop Plants Solanum Lycopersicum and Brassica Oleracea Var. Botrytis. J. Biotechnol. 2017, 262, 11–27. [Google Scholar] [CrossRef]
- Faizan, M.; Bhat, J.A.; Noureldeen, A.; Ahmad, P.; Yu, F. Zinc Oxide Nanoparticles and 24-Epibrassinolide Alleviates Cu Toxicity in Tomato by Regulating ROS Scavenging, Stomatal Movement and Photosynthesis. Ecotoxicol. Environ. Saf. 2021, 218, 112293. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Xuming, W.; Fengqing, G.; Linglan, M.; Jie, L.; Sitao, Y.; Ping, Y.; Fashui, H. Effects of Nano-Anatase on Ribulose-1, 5-Bisphosphate Carboxylase/Oxygenase Mrna Expression in Spinach. Biol. Trace Elem. Res. 2008, 126, 280–289. [Google Scholar] [CrossRef]
- Yang, H.; Yang, X.; Zhang, Y.; Heskel, M.A.; Lu, X.; Munger, J.W.; Sun, S.; Tang, J. Chlorophyll Fluorescence Tracks Seasonal Variations of Photosynthesis from Leaf to Canopy in a Temperate Forest. Glob. Chang. Biol. 2017, 23, 2874–2886. [Google Scholar] [CrossRef] [PubMed]
- Yanga, J.; Cao, W.; Rui, Y. Interactions between Nanoparticles and Plants: Phytotoxicity and Defense Mechanisms. J. Plant Interact. 2017, 12, 158–169. [Google Scholar] [CrossRef]
- Xinbin, D.; Rongxian, Z.; Wei, L. Effects of Carbonic Anhydrase in Wheat Leaf on Photosynthetic Function under Low CO2 Concentration. Zhongguo Nongye Kexue 2001, 34, 97–100. [Google Scholar]
- Tashian, R.E. Genetics of the Mammalian Carbonic Anhydrases. In Advances in Genetics; Elsevier: Amsterdam, The Netherlands, 1992; Volume 30, pp. 321–356. ISBN 0065-2660. [Google Scholar]
- Siddiqui, M.H.; Al-Whaibi, M.H. Role of Nano-SiO2 in Germination of Tomato (Lycopersicum Esculentum Seeds Mill.). Saudi J. Biol. Sci. 2014, 21, 13–17. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Faisal, M.; Al Sahli, A.A. Nano-silicon Dioxide Mitigates the Adverse Effects of Salt Stress on Cucurbita Pepo L. Environ. Toxicol. Chem. 2014, 33, 2429–2437. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, B.; Shabbir, A.; Jaleel, H.; Khan, M.M.A.; Sadiq, Y. Efficacy of Titanium Dioxide Nanoparticles in Modulating Photosynthesis, Peltate Glandular Trichomes and Essential Oil Production and Quality in Mentha Piperita L. Curr. Plant Biol. 2018, 13, 6–15. [Google Scholar] [CrossRef]
- Sivasankar, S.; Oaks, A.; Science, N. Regulation of Nitrate Reductase during Early Seedling Growth. Plant Physiol. 1995, 107, 1225–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, W.H. Nitrate Reductase Structure, Function and Regulation: Bridging the Gap between Biochemistry and Physiology. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 277–303. [Google Scholar] [CrossRef]
- Sivasankar, S.; Oaks, A. Nitrate Assimilation in Higher Plants: The Effects of Metabolites and Light. Plant Physiol. Biochem. 1996, 34, 609–620. [Google Scholar]
- Jaberzadeh, A.; Moaveni, P.; Moghadam, H.R.T.; Zahedi, H. Influence of Bulk and Nanoparticles Titanium Foliar Application on Some Agronomic Traits, Seed Gluten and Starch Contents of Wheat Subjected to Water Deficit Stress. Not. Bot. Horti Agrobot. Cluj-Napoca 2013, 41, 201. [Google Scholar] [CrossRef]
- Khan, M.N.; Mobin, M.; Abbas, Z.K.; AlMutairi, K.A.; Siddiqui, Z.H. Role of Nanomaterials in Plants under Challenging Environments. Plant Physiol. Biochem. 2017, 110, 194–209. [Google Scholar] [CrossRef] [PubMed]
- Das, P.; Barua, S.; Sarkar, S.; Karak, N.; Bhattacharyya, P.; Raza, N.; Kim, K.H.; Bhattacharya, S.S. Plant Extract–Mediated Green Silver Nanoparticles: Efficacy as Soil Conditioner and Plant Growth Promoter. J. Hazard. Mater. 2018, 346, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.D.; Huang, B.-W.; Tsuji, Y. Reactive Oxygen Species (ROS) Homeostasis and Redox Regulation in Cellular Signaling. Cell. Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef]
- Tripathy, B.C.; Oelmüller, R. Reactive Oxygen Species Generation and Signaling in Plants. Plant Signal. Behav. 2012, 7, 1621–1633. [Google Scholar] [CrossRef]
- Buchanan, B.B.; Gruissem, W.; Jones, R.L. Biochemistry & Molecular Biology of Plants; John Wiley & Sons: Hoboken, NJ, USA, 2015; Volume 53, ISBN 9788578110796. [Google Scholar]
- Andre, C.M.; Larondelle, Y.; Evers, D. Dietary Antioxidants and Oxidative Stress from a Human and Plant Perspective: A Review. Curr. Nutr. Food Sci. 2010, 6, 2–12. [Google Scholar] [CrossRef]
- Rastogi, A.; Zivcak, M.; Sytar, O.; Kalaji, H.M.; He, X.; Mbarki, S.; Brestic, M. Impact of Metal and Metal Oxide Nanoparticles on Plant: A Critical Review. Front. Chem. 2017, 5, 78. [Google Scholar] [CrossRef]
- Liu, R.; Lal, R. Potentials of Engineered Nanoparticles as Fertilizers for Increasing Agronomic Productions. Sci. Total Environ. 2015, 514, 131–139. [Google Scholar] [CrossRef]
- Nair, P.M.G.; Chung, I.M. Impact of Copper Oxide Nanoparticles Exposure on Arabidopsis Thaliana Growth, Root System Development, Root Lignificaion, and Molecular Level Changes. Environ. Sci. Pollut. Res. 2014, 21, 12709–12722. [Google Scholar] [CrossRef] [PubMed]
- Hossain, Z.; Mustafa, G.; Komatsu, S. Plant Responses to Nanoparticle Stress. Int. J. Mol. Sci. 2015, 16, 26644–26653. [Google Scholar] [CrossRef]
- Rani, P.U.; Yasur, J.; Loke, K.S.; Dutta, D. Effect of Synthetic and Biosynthesized Silver Nanoparticles on Growth, Physiology and Oxidative Stress of Water Hyacinth: Eichhornia Crassipes (Mart) Solms. Acta Physiol. Plant. 2016, 38, 58. [Google Scholar] [CrossRef]
- Amist, N.; Singh, N.B.; Yadav, K.; Singh, S.C.; Pandey, J.K. Comparative Studies of Al3+ ions and Al2O3 nanoparticles on Growth and Metabolism of Cabbage Seedlings. J. Biotechnol. 2017, 254, 1–8. [Google Scholar] [CrossRef]
- Yasmeen, F.; Raja, N.I.; Razzaq, A.; Komatsu, S. Proteomic and Physiological Analyses of Wheat Seeds Exposed to Copper and Iron Nanoparticles. Biochim. Biophys. Acta Proteins Proteom. 2017, 1865, 28–42. [Google Scholar] [CrossRef]
- Faizan, M.; Bhat, J.A.; Hessini, K.; Yu, F.; Ahmad, P. Zinc Oxide Nanoparticles Alleviates the Adverse Effects of Cadmium Stress on Oryza Sativa via Modulation of the Photosynthesis and Antioxidant Defense System. Ecotoxicol. Environ. Saf. 2021, 220, 112401. [Google Scholar] [CrossRef]
- Murshed, R.; Lopez-Lauri, F.; Sallanon, H. Effect of Salt Stress on Tomato Fruit Antioxidant Systems Depends on Fruit Development Stage. Physiol. Mol. Biol. Plants 2014, 20, 15–29. [Google Scholar] [CrossRef]
- Bohnert, H.J.; Jensen, R.G. Strategies for Engineering Water-Stress Tolerance in Plants. Trends Biotechnol. 1996, 14, 89–97. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of Glycine Betaine and Proline in Improving Plant Abiotic Stress Resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Alia; Saradhi, P.P.; Mohanty, P. Involvement of Proline in Protecting Thylakoid Membranes against Free Radical-Induced Photodamage. J. Photochem. Photobiol. B Biol. 1997, 38, 253–257. [Google Scholar] [CrossRef]
- Faraz, A.; Faizan, M.; Hayat, S.; Alam, P. Foliar application of copper oxide nanoparticles increases the photosynthetic efficiency and antioxidant activity in Brassica juncea. J. Food Qual. 2022, 2022, 5535100. [Google Scholar] [CrossRef]
- Zhao, L.; Huang, Y.; Hu, J.; Zhou, H.; Adeleye, A.S.; Keller, A.A. 1H NMR and GC-MS Based Metabolomics Reveal Defense and Detoxification Mechanism of Cucumber Plant under Nano-Cu Stress. Environ. Sci. Technol. 2016, 50, 2000–2010. [Google Scholar] [CrossRef]
- Zahra, Z.; Waseem, N.; Zahra, R.; Lee, H.; Badshah, M.A.; Mehmood, A.; Choi, H.-K.; Arshad, M. Growth and Metabolic Responses of Rice (Oryza Sativa L.) Cultivated in Phosphorus-Deficient Soil Amended with TiO2 Nanoparticles. J. Agric. Food Chem. 2017, 65, 5598–5606. [Google Scholar] [CrossRef]
- Doğaroğlu, Z.G.; Köleli, N. TiO2 and ZnO Nanoparticles Toxicity in Barley (Hordeum Vulgare L.). Clean–Soil Air Water 2017, 45, 1700096. [Google Scholar] [CrossRef]
- Faizan, M.; Faraz, A.; Hayat, S. Effective Use of Zinc Oxide Nanoparticles through Root Dipping on the Performance of Growth, Quality, Photosynthesis and Antioxidant System in Tomato. J. Plant Biochem. Biotechnol. 2020, 29, 553–567. [Google Scholar] [CrossRef]
- Jaworski, E.G. Nitrate Reductase Assay in Intact Plant Tissues. Biochem. Biophys. Res. Commun. 1971, 43, 1274–1279. [Google Scholar] [CrossRef]
- Dwivedi, R.S.; Randhawa, N.S. Evaluation of a Rapid Test for the Hidden Hunger of Zinc in Plants. Plant Soil 1974, 40, 445–451. [Google Scholar] [CrossRef]
- Arya, S.; Rookes, J.E.; Cahill, D.M.; Lenka, S.K. Chitosan Nanoparticles and Their Combination with Methyl Jasmonate for the Elicitation of Phenolics and Flavonoids in Plant Cell Suspension Cultures. Int. J. Biol. Macromol. 2022, 214, 632–641. [Google Scholar] [CrossRef]
- Faraz, A. Effects of Nanoparticles and Their Mode of Application on the Performance of Mustard. Ph.D. Thesis, Aligarh Muslim University, Aligarh, India, 2018. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid Determination of Free Proline for Water-Stress Studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faraz, A.; Faizan, M.; D. Rajput, V.; Minkina, T.; Hayat, S.; Faisal, M.; Alatar, A.A.; Abdel-Salam, E.M. CuO Nanoparticle-Mediated Seed Priming Improves Physio-Biochemical and Enzymatic Activities of Brassica juncea. Plants 2023, 12, 803. https://doi.org/10.3390/plants12040803
Faraz A, Faizan M, D. Rajput V, Minkina T, Hayat S, Faisal M, Alatar AA, Abdel-Salam EM. CuO Nanoparticle-Mediated Seed Priming Improves Physio-Biochemical and Enzymatic Activities of Brassica juncea. Plants. 2023; 12(4):803. https://doi.org/10.3390/plants12040803
Chicago/Turabian StyleFaraz, Ahmad, Mohammad Faizan, Vishnu D. Rajput, Tatiana Minkina, Shamsul Hayat, Mohammad Faisal, Abdulrahman A. Alatar, and Eslam M. Abdel-Salam. 2023. "CuO Nanoparticle-Mediated Seed Priming Improves Physio-Biochemical and Enzymatic Activities of Brassica juncea" Plants 12, no. 4: 803. https://doi.org/10.3390/plants12040803