
The Genetic Mechanism of the Immune Response to the Rice False Smut (RFS) Fungus Ustilaginoidea virens

Abstract
:1. Introduction
2. Genomics and Effectors in Ustilaginoidea virens
3. Symptoms and Resistance-Related Genes of RFS
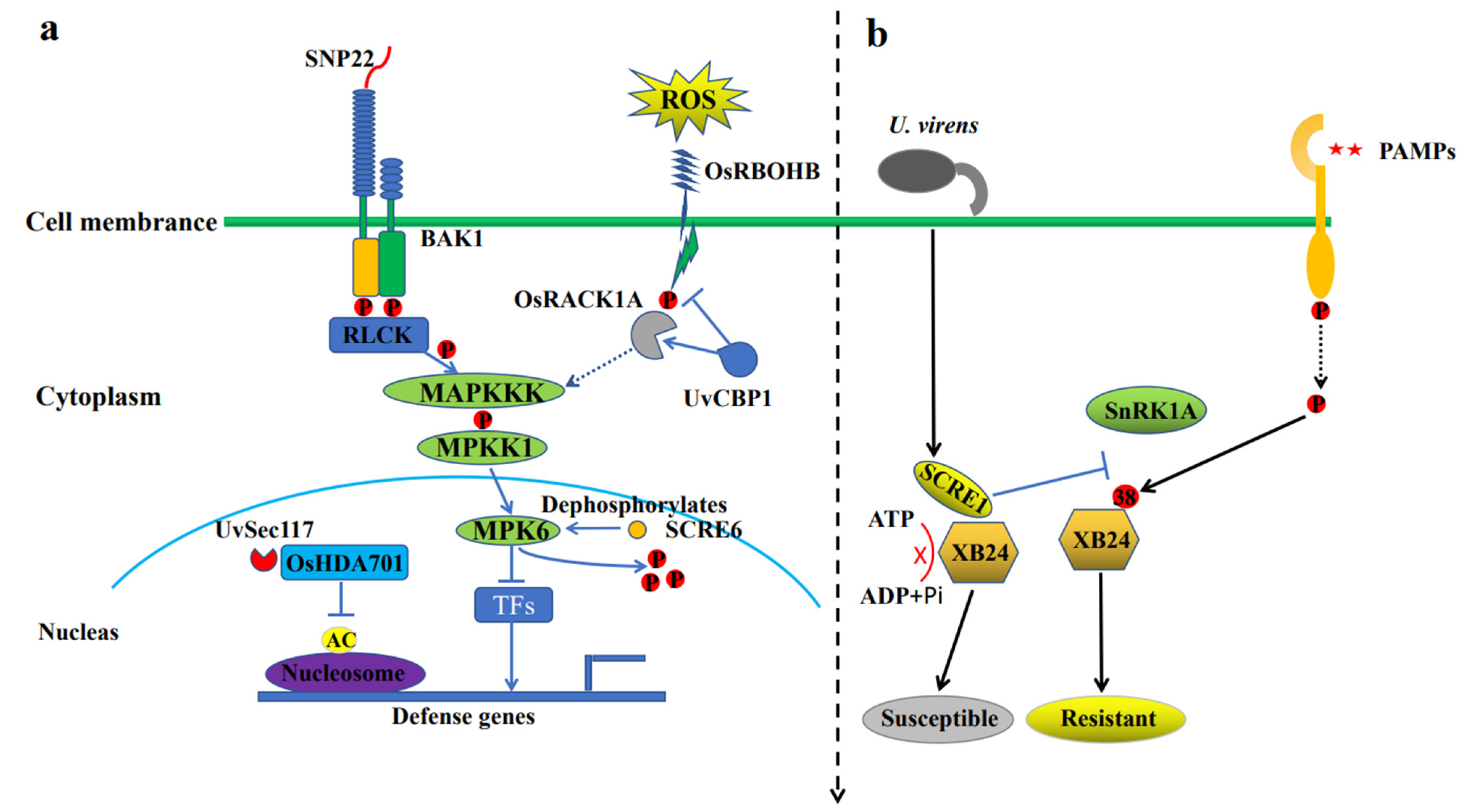
4. Genetic Mechanism of the Immune Response to RFS
5. Discussion
5.1. Methods for the Identification of Resistance to RFS
5.2. Identification of Resistance Genes and Molecular Breeding for RFS
5.3. Mechanism of Interaction between Rice R Proteins and Effector Proteins in RFS
6. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| QTL | Quantitative trait locus |
| RFS | Rice false smut |
| U. virens | Ustilaginoidea virens |
| ROS | Reactive oxygen species |
| MAMP | Microbe-associated molecular pattern |
| PTI | PIMP-triggered immunity |
| ETI | Effector-triggered immunity |
| PAMP | Pattern molecular patterns |
| PRR | Pattern recognition receptors |
| LRR-RLK | Composed of some leucine-rich repeating |
| Cys | Cysteine |
| IRRI | International Rice Research Institute |
References
- Cooke, M.C. Some extra–European fungi. Grevillea 1878, 7, 13–15. [Google Scholar]
- Lu, M.H.; Liu, W.C.; Zhu, F. Epidemic law and control technique of rice false smut in recent years. China Plant Prot. 2018, 38, 44–47. [Google Scholar]
- Kumari, S.; Kumar, J. Evaluation of yield losses and management practices of false smut in rice (Oryza sativa). Indian Phytopathol. 2015, 68, 45–49. [Google Scholar]
- Ladhalakshmi, D.; Laha, G.S.; Singh, R.; Karthikeyan, A.; Mangrauthia, S.K.; Sundaram, R.M.; Thukkaiyannan, P.; Viraktamath, B.C. Isolation and characterization of Ustilaginoidea virens and survey of false smut disease of rice in India. Phytoparasitica 2012, 40, 171–176. [Google Scholar] [CrossRef]
- Rush, M.C.A.; Shahjahan, K.M.; Jones, J.P. Outbreak of false smut of rice in Louisiana. Plant Dis. 2000, 84, 100. [Google Scholar] [CrossRef] [PubMed]
- Yong, M.L.; Deng, Q.D.; Fan, L.L.; Miao, J.K.; Lai, C.H.; Chen, H.M.; Yang, X.; Wang, S.; Chen, F.; Jin, L.; et al. The role of Ustilaginoidea virens sclerotia in increasing incidence of rice false smut disease in the subtropical zone in China. Eur. J. Plant Pathol. 2018, 150, 669–677. [Google Scholar] [CrossRef]
- Honkura, R.; Kimura, Y. Some notes on the spore disperse and the period of infection of rice false smut pathogenic fungus. Annu. Rep. Soc. Plant Prot. North Jpn. 1988, 39, 88–91. [Google Scholar]
- Jin, M.Z.; Li, Y. Survey on the sclerotia of Ustilaginoidea virens in the paddy field. J. Zhejiang Agric. Sci. 1987, 5, 238–239. [Google Scholar]
- Miao, Q.M. Studies on the sclerotium of Ustilaginoidea virens (Cooke) Tak. J. Yunnan Agric. Univ. 1994, 9, 101–104. [Google Scholar]
- Singh, R.A.; Dubey, K.S. Effect of different treatments on the dormancy of sclerotia of Claviceps oryzae-sativae. Curr. Sci. 1980, 49, 115–116. [Google Scholar]
- Ikegami, H. Studies on the false smut of rice IV. Infection of the false smut due to inoculation with chlamydospores and ascospores at the booting stage of rice plants. Res. Bull. Fac. Agric. Gifu Univ. 1960, 12, 45–51. [Google Scholar]
- Wang, G. The sexual stage of Ustilaginoidea virens and the infection process of ascospores on rice. J. Zhejiang Wanli Univ. 1995, Z1, 1–7. [Google Scholar]
- Fan, J.; Liu, J.; Gong, Z.Y.; Xu, P.Z.; Hu, X.H.; Wu, J.L.; Li, G.B.; Yang, J.; Wang, Y.Q.; Zhou, Y.F.; et al. The false smut pathogen Ustilaginoidea virens requires rice stamens for false smut ball formation. Environ. Microbiol. 2020, 22, 646–659. [Google Scholar] [CrossRef]
- Tanaka, E.; Ashizawa, T.; Sonoda, R.; Tanaka, C. Villosiclava virens gen. nov., comb. nov., teleomorph of Ustilaginoidea virens, the causal agent of rice false smut. Mycotaxon 2008, 106, 491–501. [Google Scholar]
- Fan, L.L.; Yong, M.L.; Li, D.Y.; Liu, Y.J.; Lai, C.H.; Chen, H.M.; Cheng, F.M.; Hu, D.W. Effect of temperature on the development of sclerotia in Villosiclava virens. J. Integr. Agric. 2016, 15, 2550–2555. [Google Scholar] [CrossRef]
- Fan, J.; Guo, X.Y.; Huang, F.; Li, Y.; Liu, Y.F.; Huang, F.; Wang, W.M. Epiphytic colonization of Ustilaginoidea virens on biotic and abiotic surfaces implies the widespread presence of primary inoculum for rice false smut disease. Plant Pathol. 2014, 63, 937–945. [Google Scholar] [CrossRef]
- Kim, K.W.; Park, E.W. Ultrastructure of spined conidia and hyphae of the rice false smut fungus Ustilaginoidea virens. Micron 2007, 38, 626–631. [Google Scholar] [CrossRef]
- Dhua, U.; Dhua, S.R.; Sahu, R.K. Precise disease severity assessment for false smut disease of rice. J. Phytopathol. 2015, 163, 931–940. [Google Scholar] [CrossRef]
- Nakamura, K.; Izumiyama, N.; Ohtsubo, K.; Koiso, Y.; Iwasaki, S.; Sonoda, R.; Fujita, Y.; Yaegashi, H.; Sato, Z. “Lupinosis”-like lesions in mice caused by ustiloxin, produced by Ustilaginoidea virens: A morphological study. Nat. Toxins 1994, 2, 22–28. [Google Scholar] [CrossRef]
- Zhou, L.; Lu, S.; Shan, T.; Wang, P.; Sun, W.; Chen, Z.; Wang, S. Chemistry and biology of mycotoxins from rice false smut pathogen. In Mycotoxins: Properties, Applications and Hazards; Melborn, B.J., Greene, J.C., Eds.; Nova Science Publishers Inc.: Hauppauge, NY, USA, 2012; Chapter 4; pp. 109–130. [Google Scholar]
- Lee, D.; Lal, N.K.; Lin, Z.D.; Ma, S.; Liu, J.; Castro, B.; Toruño, T.; Dinesh-Kumar, S.P.; Coaker, G. Regulation of reactive oxygen species during plant immunity through phosphorylation and ubiquitination of RBOHD. Nat. Commun. 2020, 11, 1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Liu, R.; Pang, J.; Ren, B.; Zhou, H.; Wang, G.; Wang, E.; Liu, J. Poaceae-specific cell wall-derived oligosaccharides activate plant immunity via OsCERK1 during Magnaporthe oryzae infection in rice. Nat. Commun. 2021, 12, 2178. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Qu, X.; Dong, Z.; Luo, L.; Shao, C.; Forner, J.; Lohmann, J.U.; Su, M.; Xu, M.; Liu, X.; et al. Wuschel triggers innate antiviral immunity in plant stem cells. Science 2020, 370, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.M.; Zhang, Y. Plant immunity: Danger perception and signaling. Cell 2020, 181, 978–989. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Tsuda, K. Intimate Association of PRR- and NLR-Mediated Signaling in Plant Immunity. Mol. Plant Microbe Interact. 2021, 34, 3–14. [Google Scholar] [CrossRef]
- Lüdke, D.; Yan, Q.; Rohmann, P.F.W.; Wiermer, M. NLR we there yet? Nucleocytoplasmic coordination of NLR-mediated immunity. New Phytol. 2022, 236, 24–42. [Google Scholar] [CrossRef]
- Wang, W.; Feng, B.M.; Zhou, J.M.; Tang, D.Z. Plant immune signaling: Advancing on two frontiers. J. Integr. Plant Biol. 2020, 62, 2–24. [Google Scholar] [CrossRef]
- Cui, H.; Tsuda, K.; Parker, J.E. Effector-triggered immunity: From pathogen perception to robust defense. Annu. Rev. Plant Biol. 2015, 66, 487–511. [Google Scholar]
- Yuan, M.; Jiang, Z.; Bi, G.; Nomura, K.; Liu, M.; Wang, Y.; Cai, B.; Zhou, J.M.; He, S.Y.; Xin, X.F. Pattern-recognition receptors are required for NLR-mediated plant immunity. Nature 2021, 592, 105–109. [Google Scholar] [CrossRef]
- Ngou, B.P.H.; Ahn, H.K.; Ding, P.; Jones, J.D.G. Mutual potentiation of plant immunity by cell-surface and intracellular receptors. Nature 2021, 592, 110–115. [Google Scholar]
- Zhang, Y.; Zhang, K.; Fang, A.; Han, Y.; Yang, J.; Xue, M.; Bao, J.; Hu, D.; Zhou, B.; Sun, X.; et al. Specific adaptation of Ustilaginoidea virens in occupying host florets revealed by comparative and functional genomics. Nat. Commun. 2014, 5, 3849. [Google Scholar] [CrossRef]
- NCBI (Natl. Cent. Biotechnol. Inf.). Ustilaginoidea virens; Genome Assembly Annot. Rep.; NCBI: Bethesda, MD, USA, 2019. Available online: https://www.ncbi.nlm.nih.gov/genome/genomes/31935 (accessed on 1 December 2022).
- Zhang, K.; Li, Y.J.; Li, T.J.; Li, Z.G.; Hsiang, T.; Zhang, Z.D.; Sun, W.X. Pathogenicity genes in Ustilaginoidea virens revealed by a predicted protein–protein interaction network. J. Proteome Res. 2017, 16, 1193–1206. [Google Scholar] [CrossRef]
- Catanzariti, A.M.; Dodds, P.N.; Lawrence, G.J.; Ayliffe, M.A.; Ellis, J.G. Haustorially expressed secreted proteins from flax rust are highly enriched for avirulence elicitors. Plant Cell 2006, 18, 243–256. [Google Scholar] [CrossRef] [Green Version]
- Fiorin, G.; Sanchez-Vallet, A.; Thomazella, D.P.D.; do Prado, P.F.V.; do Nascimento, L.C.; Figueira, A.V.O.; Thomma, B.P.H.J.; Pereira, G.A.G.; Teixeira, P.J.P.L. Suppression of plant immunity by fungal chitinase-like effectors. Curr. Biol. 2018, 28, 3023–3030. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Fan, J.; Fang, A.; Li, Y.; Tariqjaveed, M.; Li, D.; Hu, D.; Wang, W.M. Ustilaginoidea virens: Insights into an Emerging Rice Pathogen. Annu. Rev. Phytopathol. 2020, 58, 363–385. [Google Scholar] [CrossRef]
- Fang, A.F.; Gao, H.; Zhang, N.; Zheng, X.H.; Qiu, S.S.; Li, Y.J.; Zhou, S.; Cui, F.H.; Sun, W.X. A novel effector gene SCRE2 contributes to full virulence of Ustilaginoidea virens to rice. Front. Microbiol. 2019, 10, 845. [Google Scholar] [CrossRef]
- Zhang, N.; Yang, J.; Fang, A.F.; Wang, J.Y.; Li, D.Y.; Li, Y.J.; Wang, S.Z.; Cui, F.H.; Yu, J.J.; Liu, Y.J. The essential effector SCRE1 in Ustilaginoidea virens suppresses rice immunity via a small peptide region. Mol. Plant Pathol. 2020, 21, 445–459. [Google Scholar] [CrossRef]
- Qiu, S.; Fang, A.; Zheng, X.; Wang, S.; Wang, J.; Fan, J.; Sun, Z.; Gao, H.; Yang, J.; Zeng, Q. Ustilaginoidea virens Nuclear effector SCRE4 suppresses rice immunity via inhibiting expression of a positive immune regulator OsARF17. Int. J. Mol. Sci. 2022, 23, 10527. [Google Scholar] [CrossRef]
- Zheng, X.; Fang, A.; Qiu, S.; Zhao, G.; Wang, J.; Wang, S.; Wei, J.; Gao, H.; Yang, J.; Mou, B.; et al. Ustilaginoidea virens secretes a family of phosphatases that stabilize the negative immune regulator OsMPK6 and suppress plant immunity. Plant Cell 2022, 34, 3088–3109. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xiang, S.; Wang, Y.; Zhou, J.; Hai, Y.; Peng, X.; Wang, Y.; Wei, S. UvHrip1, an effector secreted by Ustilaginoidea virens, suppresses basal defense and promotes disease development in Arabidopsis thaliana. Gene 2020, 751, 144776. [Google Scholar] [CrossRef] [PubMed]
- Toruno, T.Y.; Stergiopoulos, I.; Coaker, G. Plant-pathogen effectors: Cellular probes interfering with plant defenses in spatial and temporal manners. Annu. Rev. Phytopathol. 2016, 54, 419–441. [Google Scholar] [CrossRef] [PubMed]
- Win, J.; Chaparro-Garcia, A.; Belhaj, K.; Saunders, D.G.O.; Yoshida, K.; Dong, S.; Schornack, S.; Zipfel, C.; Robatzek, S.; Hogenhout, S.A.; et al. Effector biology of plantassociated organisms: Concepts and perspectives. Cold Spring Harb. Symp. Quant. Biol. 2012, 77, 235–247. [Google Scholar] [CrossRef]
- Li, Y.S.; Huang, S.D.; Yang, J.; Wang, C.-L. Analysis of Quantitative Trait Loci for Resistance to Rice False Smut. Acta Agron. Sin. 2011, 37, 778–783. [Google Scholar] [CrossRef]
- Xu, J.L.; Xue, Q.Z.; Luo, L.J.; Li, Z.K. Preliminary report on quantitative trait loci mapping of false smut resistance using near-isogenic introgression lines in rice. Acta Agric. Zhejiangensis 2002, 14, 14–19. [Google Scholar]
- Yuan, L.H.; Li, M.; Li, J.H. Study of genetic algorithmsbased on pleiotropy. Syst. Eng. Electron. 2007, 29, 1969–1972. [Google Scholar]
- Zhou, Y.L.; Xie, X.W.; Zhang, F.; Wang, S.; Liu, X.-Z.; Zhu, L.-H.; Xu, J.-L.; Gao, Y.-M.; Li, Z.-K. Detection of quantitative resistance loci associated with resistance to rice false smut (Ustilaginoidea virens) using introgression lines. Plant Pathol. 2014, 63, 365–372. [Google Scholar] [CrossRef]
- Qiu, J.H.; Lu, F.F.; Wang, H.; Xie, J.; Wang, C.; Liu, Z.; Meng, S.; Shi, H.; Shen, X.; Kou, Y. A candidate gene for the determination of rice resistant to rice false smut. Mol. Breed. 2020, 40, 105. [Google Scholar] [CrossRef]
- Long, W.X.; Yuan, Z.Q.; Fan, F.F.; Dan, D.; Pan, G.; Sun, H.; Zhang, Z.; Li, N.; Li, S. Genome-wide association analysis of resistance to rice false smut. Mol. Breed. 2020, 40, 46. [Google Scholar] [CrossRef]
- Neelam, K.; Kumar, K.; Kaur, A.; Kishore, A.; Kaur, P.; Babbar, A.; Kaur, G.; Kamboj, I.; Lore, J.S.; Vikal, Y.; et al. High-resolution mapping of the quantitative trait locus (QTLs) conferring resistance to false smut disease in rice. J. Appl. Genet. 2022, 63, 35–45. [Google Scholar] [CrossRef]
- Huang, Y.; Cui, K.; Zhang, Z.; Chai, R.; Xie, H.; Shou, J.; Fu, J.; Li, G.; Liu, J.; Wu, S.; et al. Identification and fine-mapping of quantitative trait loci (QTL) conferring rice false smut resistance in rice. J. Genet. Genom. 2022, 8527, 00254–00255. [Google Scholar] [CrossRef]
- Song, T.Q.; Zhang, Y.; Zhang, Q.; Zhang, X.; Shen, D.; Yu, J.; Yu, M.; Pan, X.; Cao, H.; Yong, M.; et al. The N-terminus of an Ustilaginoidea virens Ser-Thr-rich glycosylphosphatidylinositol- anchored protein elicits plant immunity as a MAMP. Nat. Commun. 2021, 12, 2451. [Google Scholar] [CrossRef]
- Pei, D.; Hua, D.; Deng, J.; Wang, Z.; Song, C.; Wang, Y.; Wang, Y.; Qi, J.; Kollist, H.; Yang, S.; et al. Phosphorylation of the plasma membrane H+-ATPase AHA2 by BAK1 is required for ABA-induced stomatal closure in Arabidopsis. Plant Cell 2022, 34, 2708–2729. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Y.; Duan, Y.H.; Qiao, F.G.; Liu, H.; Huang, J.; Luo, C.; Chen, X.; Li, G.; Xie, K.; Hsiang, T.; et al. A secreted fungal effector suppresses rice immunity through host histone hypoacetylation. New Phytol. 2022, 235, 1977–1994. [Google Scholar] [CrossRef] [PubMed]
- Li, G.B.; He, J.X.; Wu, J.L.; Wang, H.; Zhang, X.; Liu, J.; Hu, X.H.; Zhu, Y.; Shen, S.; Bai, Y.F.; et al. Overproduction of OsRACK1A, an effector-targeted scaffold protein promoting OsRBOHB-mediated ROS production, confers rice floral resistance to false smut disease without yield penalty. Mol. Plant 2022, 15, 1790–1806. [Google Scholar] [CrossRef]
- Cheng, Z.; Li, J.F.; Niu, Y.; Zhang, X.C.; Woody, O.Z.; Xiong, Y.; Djonovic, S.; Millet, Y.; Bush, J.; McConkey, B.J.; et al. Pathogen-secreted proteases activate a novel plant immune pathway. Nature 2015, 521, 213–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Zhang, N.; Wang, J.; Fang, A.; Fan, J.; Li, D.; Li, Y.; Wang, S.; Cui, F.; Yu, J.; et al. SnRK1A-mediated phosphorylation of a cytosolic ATPase positively regulates rice innate immunity and is inhibited by Ustilaginoidea virens effector SCRE1. New Phytol. 2022, 236, 1422–1440. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.S.; Gao, J.Z.; Cao, G.P.; Huang, S.X.; Liu, R.M.; Liu, C.D. Studies on the grading standard of rice false smut disease. Plant Prot. 2001, 27, 18–21. [Google Scholar]
- Song, J.H.; Wang, Y.F.; Yin, W.X.; Huang, J.B.; Luo, C.X. Effect of chemical seed treatment on rice false smut control in field. Plant Dis. 2021, 105, 3218–3223. [Google Scholar] [CrossRef] [PubMed]
- Khanal, S.; Zhou, X.G.; Gaire, S. Kernel Smut and False smut: The old-emerging diseases of rice—A review. Phytopathology 2022, 1–42. [Google Scholar] [CrossRef]
- Deng, R.; Tao, M.; Xing, H.; Yang, X.; Liu, C.; Liao, K.; Qi, L. Automatic diagnosis of rice diseases using deep learning. Front. Plant Sci. 2021, 12, 701038. [Google Scholar] [CrossRef]
- Li, X.; Huang, R.; Liu, J.; Xu, G.; Yuan, M. Engineering false smut resistance rice via host-induced gene silencing of two chitin synthase genes of Ustilaginoidea virens. Plant Biotechnol. J. 2021, 19, 2386–2388. [Google Scholar] [CrossRef]
- Chen, X.; Pei, Z.; Liu, H.; Huang, J.; Chen, X.; Luo, C.; Hsiang, T.; Zheng, L. Host-induced gene silencing of fungal-specific genes of Ustilaginoidea virens confers effective resistance to rice false smut. Plant Biotechnol. J. 2022, 20, 253–255. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, Y.; Yu, X.; Lin, Y.; Zhu, Y.; Chen, J.; Xie, H.; Zhang, Q.; Wang, L.; Wei, Y.; et al. SH3P2, an SH3 domain-containing protein that interacts with both Pib and AvrPib, suppresses effector-triggered, Pib-mediated immunity in rice. Mol. Plant 2022, 15, 1931–1946. [Google Scholar] [CrossRef]
- van der Biezen, E.A.; Jones, J.D. Plant disease-resistance proteins and the gene-for-gene concept. Trends Biochem. Sci. 1998, 23, 454–456. [Google Scholar] [CrossRef]
- Tasset, C.; Bernoux, M.; Jauneau, A.; Pouzet, C.; Brière, C.; Kieffer-Jacquinod, S.; Rivas, S.; Marco, Y.; Deslandes, L. Autoacetylation of the Ralstonia solanacearum effector PopP2 targets a lysine residue essential for RRS1-Rmediated immunity in Arabidopsis. PLoS Pathog. 2010, 6, e1001202. [Google Scholar] [CrossRef]
- Zhang, J.; Li, W.; Xiang, T.; Liu, Z.; Laluk, K.; Ding, X.; Zou, Y.; Gao, M.; Zhang, X.; Chen, S.; et al. Receptor-like cytoplasmic kinases integrate signaling from multiple plant immune receptors and are targeted by a Pseudomonassyringae effector. Cell Host Microbe 2010, 7, 290–301. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Zhu, L.; Song, T.; Wang, Y.; Zhang, Q.; Xia, Y.; Qiu, M.; Lin, Y.; Li, H.; Kong, L.; et al. A paralogous decoy protects Phytophthora sojae apoplastic effector PsXEG1 from a host inhibitor. Science 2017, 355, 710–714. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Chen, J.; Li, M.; Chang, M.; Xu, K.; Shang, Z.; Zhao, Y.; Palmer, I.; Zhang, Y.; McGill, J.; et al. A bacterial type III effector targets the master regulator of salicylic acid signaling NPR1 to subvert plant immunity. Cell Host Microbe 2017, 22, 777–788.e7. [Google Scholar] [CrossRef] [PubMed]
- Lopez, V.; Park, B.C.; Nowak, D. A bacterial effector mimics a host HSP90 client to undermine immunity. Cell 2019, 179, 205–218.e21. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Gong, H.; Song, T.; Yu, M.; Pan, X.; Yu, J.; Qi, Z.; Du, Y.; Liu, Y. The adaptor protein UvSte50 governs fungal pathogenicity of Ustilaginoidea virens via the MAPK signaling pathway. J. Fungi 2022, 8, 954. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Yang, J.; Wang, Y.Q.; Li, G.B.; Li, Y.; Huang, F.; Wang, W.M. Current understanding on Villosiclava virens, a unique flower-infecting fungus causing rice false smut disease. Mol. Plant Pathol. 2016, 17, 1321–1330. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, T.; Luo, C.; Jiang, D.; Li, G.; Li, Q.; Hsiang, T.; Huang, J. Prevalence and diversity of mycoviruses infecting the plant pathogen Ustilaginoidea virens. Virus Res. 2015, 195, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.J.; Chen, D.; Zhong, J.; Zhang, S.Y.; Gao, B.D. A novel mycovirus identified from the rice false smut fungus Ustilaginoidea virens. Virus Genes 2015, 51, 159–162. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene | Gene Product | Pathogenicity of Deletion Mutant | Gene Function | Reference |
|---|---|---|---|---|
| UV_1261/SCRE2 | Effector protein | Reduced virulence | Suppresses cell death and pattern-triggered immunity in plant | [31,37] |
| UV_5215 | Effector protein | Reduced virulence | Suppresses B. glumae-induced HR in N. benthamiana | [31] |
| UV_2964 | Effector protein | Reduced virulence | Suppresses B. glumae-induced HR in N. benthamiana | [31] |
| SCRE1 | Effector protein | Reduced virulence | Inhibits host immunity | [38] |
| SCRE4 | Effector protein | Reduced virulence | An essential virulence effector and suppresses the expression of OsARF17 | [39] |
| SCRE6 | Effector protein | Reduced virulence | Interacts with and dephosphorylates the negative immune regulator OsMPK6 in rice, and suppresses plant immunity. | [40] |
| UvHrip1 | Effector protein | Reduced virulence | Suppresses plant innate immunity and promotes disease development | [41] |
| Name | Chromosome | Resistant Material | Reference |
|---|---|---|---|
| qFsrl | 1 | IR28 | [44] |
| qFsr2 | 2 | IR28 | [44] |
| qFsr4 | 4 | IR28 | [44] |
| qFsr8 | 8 | IR28 | [44] |
| qFsr10 | 10 | IR28 | [44] |
| qFsr11 | 11 | IR28 | [44] |
| qFsr12 | 12 | IR28 | [44] |
| QFsr10 | 10 | Near-isogenic line | [45] |
| QFsr12 | 12 | Near-isogenic line | [45] |
| qFsr2-1 | 2 | MR183-2, 08R2394 | [46] |
| qFsr2-2 | 2 | MR183-2, 08R2394 | [46] |
| qFsr3-1 | 3 | MR183-2, 08R2394 | [46] |
| qFsr8-1 | 8 | MR183-2, 08R2394 | [46] |
| qFSR-6-7 | 6 | Near-isogenic line | [47] |
| qFSR-10-5 | 10 | Near-isogenic line | [47] |
| qFSR-10-2 | 10 | Near-isogenic line | [47] |
| qFSR-11-2 | 11 | Near-isogenic line | [47] |
| FSR1 | 11 | NanJing11 | [48] |
| LOC_Os01g15580 | 1 | 315 core rice materials | [49] |
| qRFSr5.3 | 5 | RYT2668 | [50] |
| qRFSr7.1a | 7 | RYT2668 | [50] |
| qRFsr9.1 | 9 | RYT2668 | [50] |
| qRFSr2.2 | 2 | RYT2668 | [50] |
| qRFSr4.3 | 4 | RYT2668 | [50] |
| qRFSr5.4 | 5 | RYT2668 | [50] |
| qRFSr7.1b | 7 | RYT2668 | [50] |
| qFSR10 | 10 | XS47, XS664 | [51] |
| qFSR2 | 2 | XS47, XS664 | [51] |
| qFSR9 | 9 | XS47, XS664 | [51] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, D.; He, N.; Huang, F.; Jin, Y.; Li, S. The Genetic Mechanism of the Immune Response to the Rice False Smut (RFS) Fungus Ustilaginoidea virens. Plants 2023, 12, 741. https://doi.org/10.3390/plants12040741
Yang D, He N, Huang F, Jin Y, Li S. The Genetic Mechanism of the Immune Response to the Rice False Smut (RFS) Fungus Ustilaginoidea virens. Plants. 2023; 12(4):741. https://doi.org/10.3390/plants12040741
Chicago/Turabian StyleYang, Dewei, Niqing He, Fenghuang Huang, Yidan Jin, and Shengping Li. 2023. "The Genetic Mechanism of the Immune Response to the Rice False Smut (RFS) Fungus Ustilaginoidea virens" Plants 12, no. 4: 741. https://doi.org/10.3390/plants12040741