
Effects of Hydrogen Sulfide on Sugar, Organic Acid, Carotenoid, and Polyphenol Level in Tomato Fruit


Abstract
:1. Introduction
2. Results
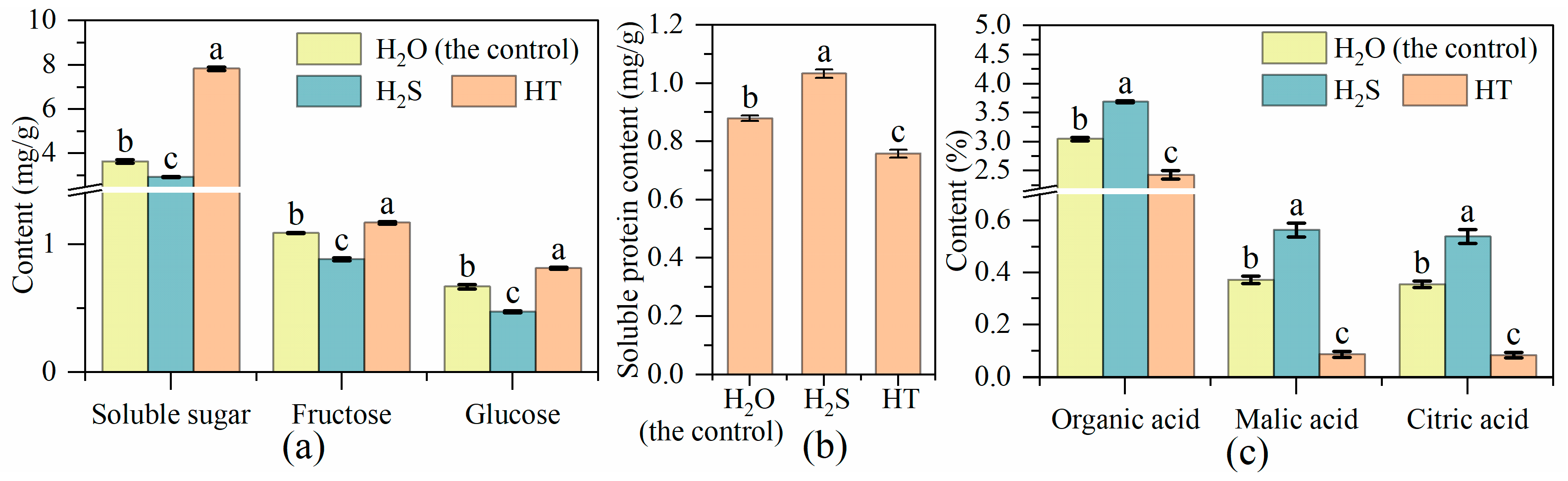
2.1. Effects of H2S on Soluble Sugar, Soluble Protein and Organic Acids Content in Tomato Fruit
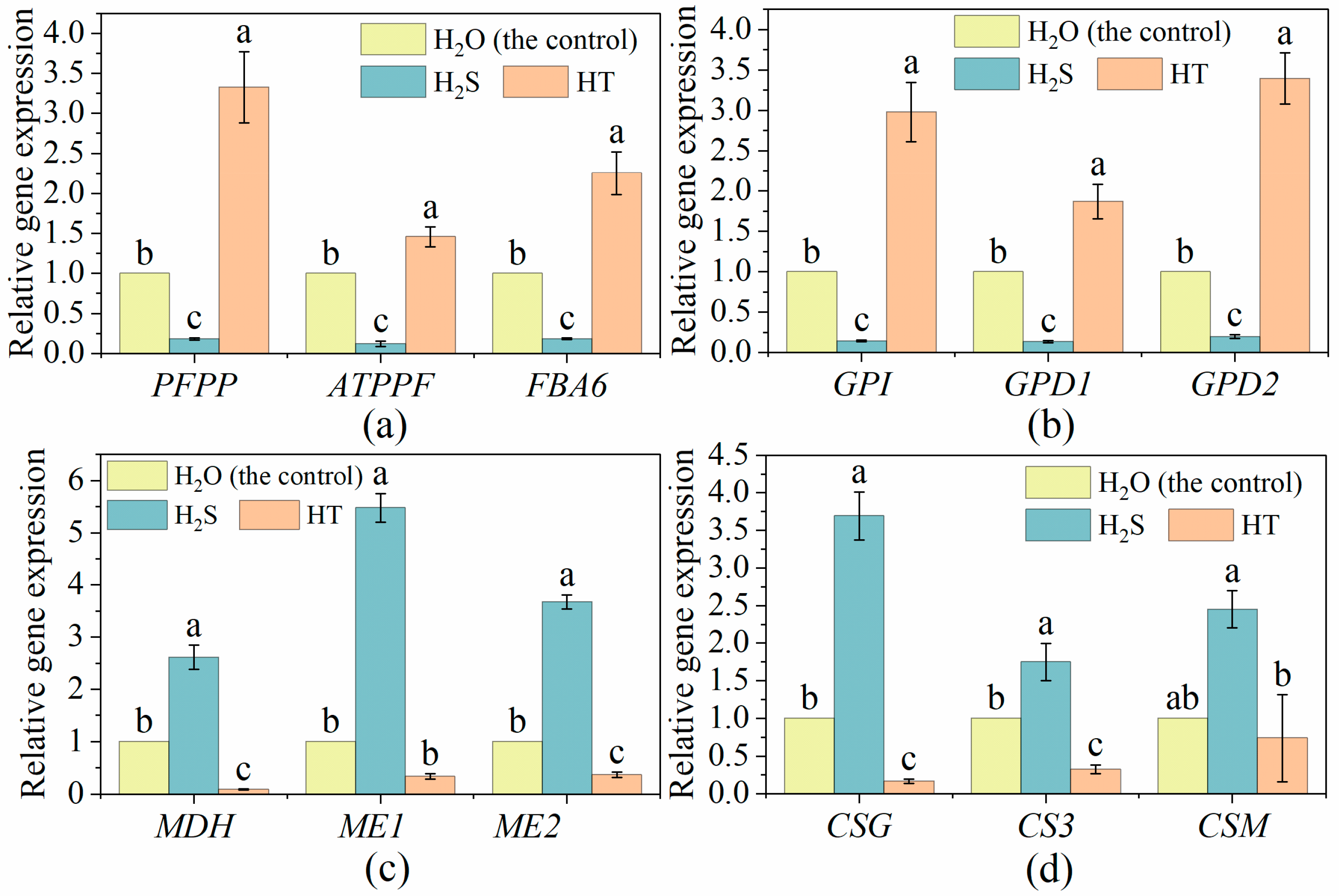
2.2. Effects of H2S on the Expression of Genes Related to Soluble Sugar and Organic Acids Metabolism in Tomato Fruit
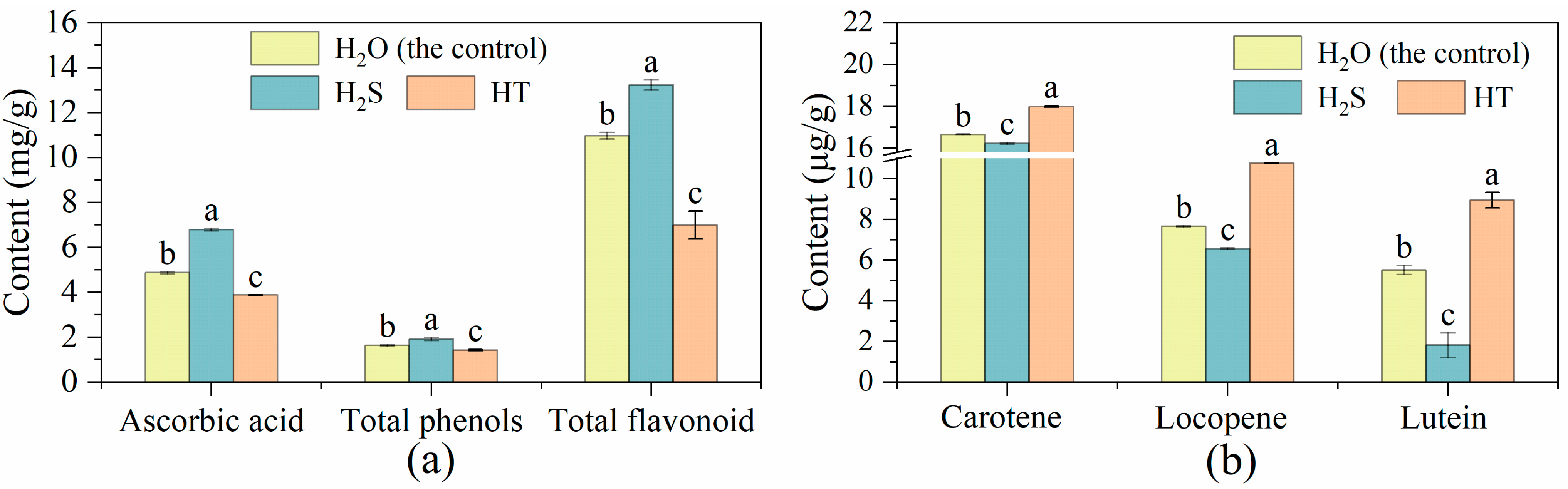
2.3. Effects of H2S on Ascorbic Acid, Total Phenols, Total Flavonoid and Carotenes Content in Tomato Fruit
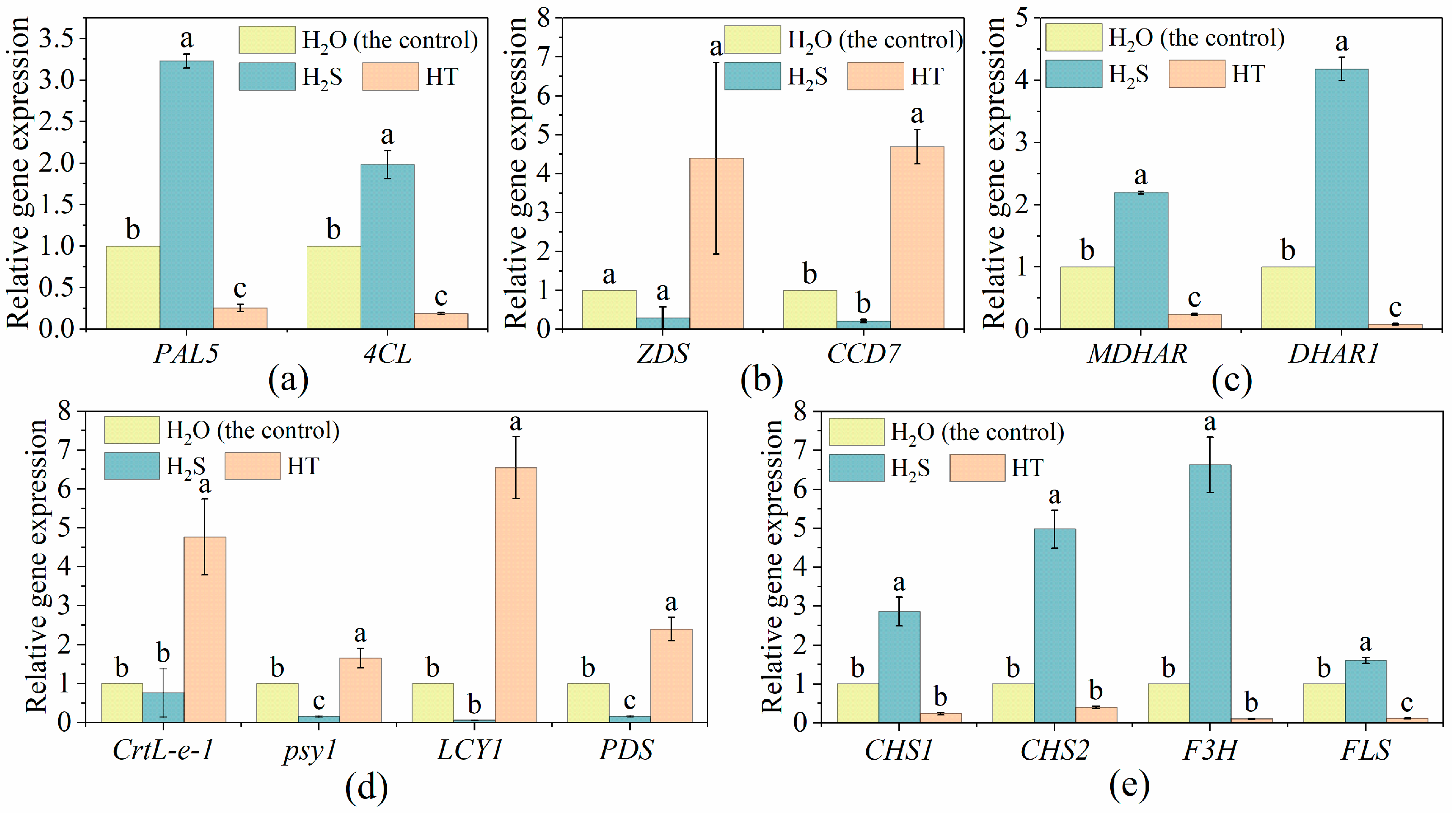
2.4. Effects of H2S on the Expression of Genes Related to Polyphenols, Flavonoid, Ascorbic Acid and Carotene Metabolism in Tomato Fruit
2.5. Effects of H2S on Polyphenol Content in Tomato Fruit
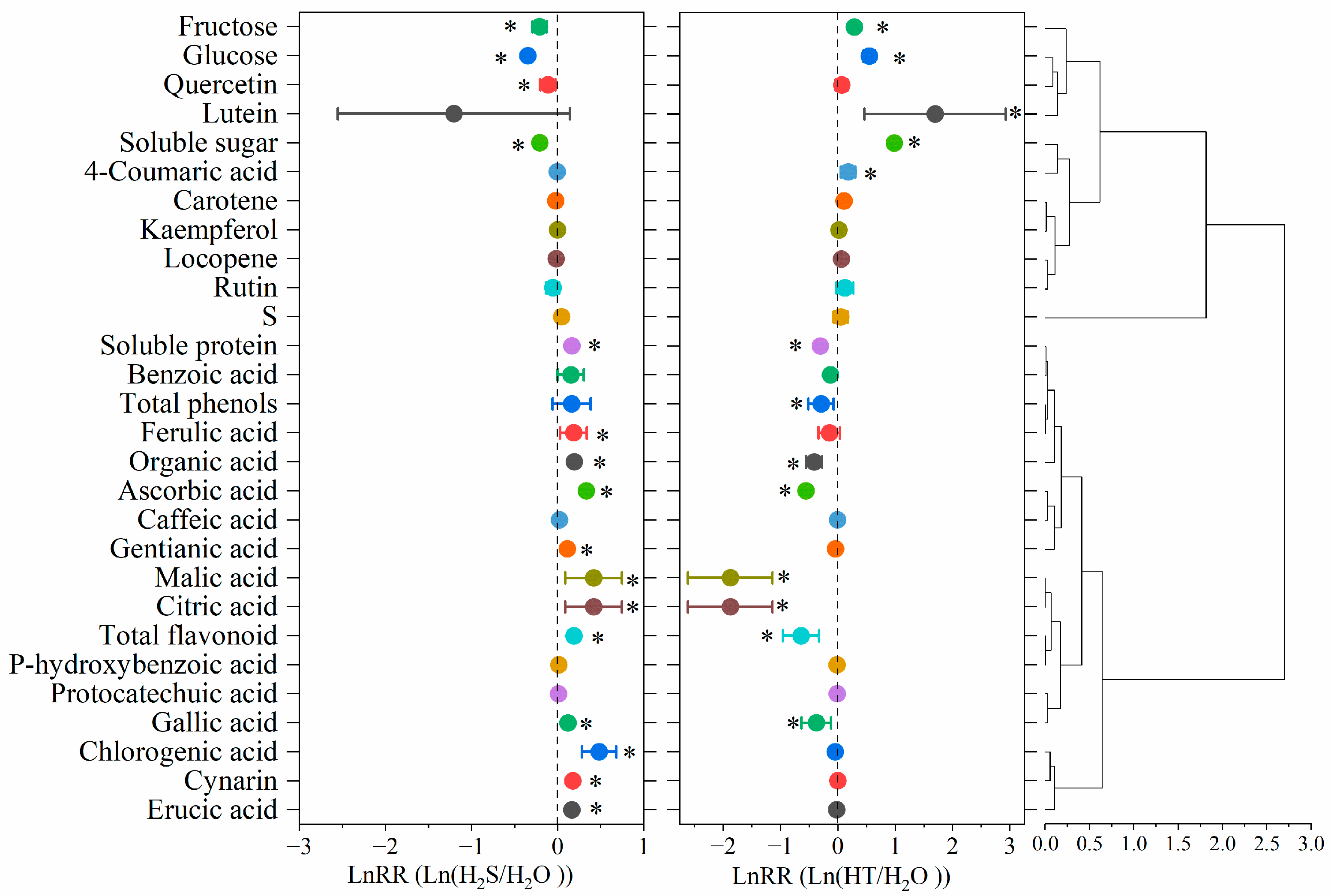
2.6. Effect Sizes of H2S on Flavor Quality in Tomato Fruit
3. Discussion
4. Materials and Methods
4.1. Plant Material and Treatment Conditions
4.2. Determination of Soluble Sugar and Soluble Protein Content
4.3. Determination of Sugar and Acid Fraction
4.4. Determination of Lycopene, Carotene and Lutein Content
4.5. Determination of Total Phenol, Total Flavonoid and Ascorbic Acid Content
4.6. Determination of the Content of Polyphenols
4.7. Total RNA Isolation and Gene Expression Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, R. The gasotransmitter role of hydrogen sulfide. Antioxid. Redox Signal. 2003, 5, 493–501. [Google Scholar] [CrossRef]
- Baskar, R.; Bian, J. Hydrogen sulfide gas has cell growth regulatory role. Eur. J. Pharmacol. 2011, 656, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-G.; Min, X.; Zhou, Z.-H. Hydrogen sulfide: A signal molecule in plant cross-adaptation. Front. Plant Sci. 2016, 7, 1621. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Huo, J.; Liao, W. Hydrogen sulfide: Roles in plant abiotic stress response and crosstalk with other signals. Plant Sci. 2021, 302, 110733. [Google Scholar] [CrossRef]
- Mei, Y.; Zhao, Y.; Jin, X.; Wang, R.; Xu, N.; Hu, J.; Huang, L.; Guan, R.; Shen, W. L-Cysteine desulfhydrase-dependent hydrogen sulfide is required for methane-induced lateral root formation. Plant Mol. Biol. 2019, 99, 283–298. [Google Scholar] [CrossRef]
- Li, Z.-G. Analysis of some enzymes activities of hydrogen sulfide metabolism in plants. Meth. Enzymol. 2015, 555, 253–269. [Google Scholar]
- Huo, J.; Huang, D.; Zhang, J.; Fang, H.; Wang, B.; Wang, C.; Liao, W. Hydrogen sulfide: A gaseous molecule in postharvest freshness. Front. Plant Sci. 2018, 9, 1172. [Google Scholar] [CrossRef]
- Song, J.; Trueba, S.; Yin, X.; Cao, K.; Brodribb, T.; Hao, G. Hydraulic vulnerability segmentation in compound-leaved trees: Evidence from an embolism visualization technique. Mediat. Inflamm. 2022, 189, 204–214. [Google Scholar] [CrossRef]
- Dooley, F.D.; Nair, S.P.; Ward, P.D. Increased growth and germination success in plants following hydrogen sulfide administration. PLoS ONE 2013, 8, e62048. [Google Scholar] [CrossRef]
- Hu, H.; Shen, W.; Li, P. Effects of hydrogen sulphide on quality and antioxidant capacity of mulberry fruit. Int. J. Food Sci. Technol. 2014, 49, 399–409. [Google Scholar] [CrossRef]
- Wang, W.; Ni, Z.-J.; Thakur, K.; Cao, S.-Q.; Wei, Z.-J. Recent update on the mechanism of hydrogen sulfide improving the preservation of postharvest fruits and vegetables. Curr. Opin. Food Sci. 2022, 47, 100906. [Google Scholar] [CrossRef]
- Tayal, R.; Kumar, V.; Irfan, M. Harnessing the power of hydrogen sulphide (H2S) for improving fruit quality traits. Plant Biol. 2022, 24, 594–601. [Google Scholar] [CrossRef]
- Hu, L.-Y.; Hu, S.-L.; Wu, J.; Li, Y.-H.; Zheng, J.-L.; Wei, Z.-J.; Liu, J.; Wang, H.-L.; Liu, Y.-S.; Zhang, H. Hydrogen sulfide prolongs postharvest shelf life of strawberry and plays an antioxidative role in fruits. J. Agric. Food Chem. 2012, 60, 8684–8693. [Google Scholar] [CrossRef] [PubMed]
- Al Ubeed, H.; Wills, R.; Bowyer, M.; Golding, J. Interaction of the hydrogen sulphide inhibitor, propargylglycine (PAG), with hydrogen sulphide on postharvest changes of the green leafy vegetable, pak choy. Postharvest Biol. Technol. 2019, 147, 54–58. [Google Scholar] [CrossRef]
- Li, D.; Limwachiranon, J.; Li, L.; Du, R.; Luo, Z. Involvement of energy metabolism to chilling tolerance induced by hydrogen sulfide in cold-stored banana fruit. Food Chem. 2016, 208, 272–278. [Google Scholar] [CrossRef]
- Zhang, H.; Hu, S.-L.; Zhang, Z.-J.; Hu, L.-Y.; Jiang, C.-X.; Wei, Z.-J.; Liu, J.; Wang, H.-L.; Jiang, S.-T. Hydrogen sulfide acts as a regulator of flower senescence in plants. Postharvest Biol. Technol. 2011, 60, 251–257. [Google Scholar] [CrossRef]
- Li, T.-T.; Li, Z.-R.; Hu, K.-D.; Hu, L.-Y.; Chen, X.-Y.; Li, Y.-H.; Yang, Y.; Yang, F.; Zhang, H. Hydrogen sulfide alleviates kiwifruit ripening and senescence by antagonizing effect of ethylene. HortScience 2017, 52, 1556–1562. [Google Scholar] [CrossRef]
- Ge, Y.; Hu, K.-D.; Wang, S.-S.; Hu, L.-Y.; Chen, X.-Y.; Li, Y.-H.; Yang, Y.; Yang, F.; Zhang, H. Hydrogen sulfide alleviates postharvest ripening and senescence of banana by antagonizing the effect of ethylene. PLoS ONE 2017, 12, e0180113. [Google Scholar] [CrossRef]
- Ali, S.; Nawaz, A.; Ejaz, S.; Haider, S.T.-A.; Alam, M.W.; Javed, H.U. Effects of hydrogen sulfide on postharvest physiology of fruits and vegetables: An overview. Sci. Hortic. 2019, 243, 290–299. [Google Scholar] [CrossRef]
- Chang, Z.; Jingying, S.; Liqin, Z.; Changle, L.; Qingguo, W. Cooperative effects of hydrogen sulfide and nitric oxide on delaying softening and decay of strawberry. Int. J. Agric. Biol. Eng. 2014, 7, 114–122. [Google Scholar]
- FAOSTAT. 2015. Available online: http//:www.fao.org/faostat (accessed on 12 December 2022).
- Ellinger, S.; Ellinger, J.; Stehle, P. Tomatoes, tomato products and lycopene in the prevention and treatment of prostate cancer: Do we have the evidence from intervention studies? Curr. Opin. Clin. Nutr. 2006, 9, 722–727. [Google Scholar] [CrossRef]
- Weisburger, J.H. Lycopene and tomato products in health promotion. Exp. Biol. Med. 2002, 227, 924–927. [Google Scholar] [CrossRef] [PubMed]
- Abushita, A.; Daood, H.; Biacs, P. Change in carotenoids and antioxidant vitamins in tomato as a function of varietal and technological factors. J. Agric. Food Chem. 2000, 48, 2075–2081. [Google Scholar] [CrossRef] [PubMed]
- Giovanelli, G.; Lavelli, V.; Peri, C.; Nobili, S. Variation in antioxidant components of tomato during vine and post-harvest ripening. J. Agric. Food Chem. 1999, 79, 1583–1588. [Google Scholar] [CrossRef]
- Raiola, A.; Rigano, M.M.; Calafiore, R.; Frusciante, L.; Barone, A. Enhancing the health-promoting effects of tomato fruit for biofortified food. Mediat. Inflamm. 2014, 2014, 139873. [Google Scholar] [CrossRef]
- Liu, Z.; Alseekh, S.; Brotman, Y.; Zheng, Y.; Fei, Z.; Tieman, D.M.; Giovannoni, J.J.; Fernie, A.R.; Klee, H.J. Identification of a Solanum pennellii chromosome 4 fruit flavor and nutritional quality-associated metabolite QTL. Front. Plant Sci. 2016, 7, 1671. [Google Scholar] [CrossRef]
- Martí, R.; Roselló, S.; Cebolla-Cornejo, J. Tomato as a source of carotenoids and polyphenols targeted to cancer prevention. Cancers 2016, 8, 58. [Google Scholar] [CrossRef]
- Riso, P.; Visioli, F.; Erba, D.; Testolin, G.; Porrini, M. Lycopene and vitamin C concentrations increase in plasma and lymphocytes after tomato intake. Effects on cellular antioxidant protection. Eur. J. Clin. Nutr. 2004, 58, 1350–1358. [Google Scholar] [CrossRef]
- Viuda-Martos, M.; Sanchez-Zapata, E.; Sayas-Barberá, E.; Sendra, E.; Pérez-Álvarez, J.; Fernández-López, J. Tomato and tomato byproducts. Human health benefits of lycopene and its application to meat products: A review. Crit. Rev. Food. Sci. 2014, 54, 1032–1049. [Google Scholar] [CrossRef]
- Tohge, T.; Fernie, A.R. Metabolomics-inspired insight into developmental, environmental and genetic aspects of tomato fruit chemical composition and quality. Plant Cell Physiol. 2015, 56, 1681–1696. [Google Scholar] [CrossRef]
- Gadzhieva, A.M.; Kasyanov, G.I.; Mungieva, N.A.; Musaeva, N.M.; Alivova, N.M. Effective complex processing of raw tomatoes. Sci. Study Res. Chem. Chem. Eng. Biotechnol. Food Ind. 2018, 19, 83–89. [Google Scholar]
- FAOSTAT. 2022. Available online: https://faostat3.fao.org/download/Q/QC/E (accessed on 12 December 2022).
- Rehman, M.; Khan, N.; Jan, I. Post harvest losses in tomato crop (a case study of Peshawar Valley). Sarhad J. Agric. 2007, 23, 1279–1284. [Google Scholar]
- Peralta-Ruiz, Y.; Tovar, C.D.G.; Sinning-Mangonez, A.; Coronell, E.A.; Marino, M.F.; Chaves-Lopez, C. Reduction of Postharvest Quality Loss and Microbiological Decay of Tomato “Chonto” (Solanum lycopersicum L.) Using Chitosan-E Essential Oil-Based Edible Coatings under Low-Temperature Storage. Polymers 2020, 12, 1822. [Google Scholar] [CrossRef]
- Arah, I.K.; Amaglo, H.; Kumah, E.K.; Ofori, H. Preharvest and postharvest factors affecting the quality and shelf life of harvested tomatoes: A mini review. Int. J. Agron. 2015, 2015, 478041. [Google Scholar] [CrossRef] [Green Version]
- Paul, V.; Pandey, R. Role of internal atmosphere on fruit ripening and storability—A review. J. Food Sci. Technol. 2014, 51, 1223–1250. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-P.; Hu, K.-D.; Hu, L.-Y.; Li, Y.-H.; Jiang, A.-M.; Xiao, F.; Han, Y.; Liu, Y.-S.; Zhang, H. Hydrogen sulfide alleviates postharvest senescence of broccoli by modulating antioxidant defense and senescence-related gene expression. J. Agric. Food Chem. 2014, 62, 1119–1129. [Google Scholar] [CrossRef]
- Zhu, L.; Wang, W.; Shi, J.; Zhang, W.; Shen, Y.; Du, H.; Wu, S. Hydrogen sulfide extends the postharvest life and enhances antioxidant activity of kiwifruit during storage. J. Sci. Food Agric. 2014, 94, 2699–2704. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.-L.; Hu, L.-Y.; Hu, K.-D.; Wu, J.; Yang, F.; Zhang, H. Hydrogen sulfide alleviates senescence of fresh-cut apple by regulating antioxidant defense system and senescence-related gene expression. HortScience 2016, 51, 152–158. [Google Scholar] [CrossRef]
- Yao, G.-F.; Li, C.; Sun, K.-K.; Tang, J.; Huang, Z.-Q.; Yang, F.; Huang, G.-G.; Hu, L.-Y.; Jin, P.; Hu, K.-D. Hydrogen sulfide maintained the good appearance and nutrition in post-harvest tomato fruit by antagonizing the effect of ethylene. Front. Plant Sci. 2020, 11, 584. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, H.; Zhang, Y.; Martin, C. Can the world’s favorite fruit, tomato, provide an effective biosynthetic chassis for high-value metabolites? Plant Cell Rep. 2018, 37, 1443–1450. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Zhang, X.; Gong, B.; Yan, Y.; Shi, Q. Proteomics and metabolomics analysis of tomato fruit at different maturity stages and under salt treatment. Food Chem. 2020, 311, 126009. [Google Scholar] [CrossRef]
- Thole, V.; Vain, P.; Martin, C. Effect of Elevated Temperature on Tomato Post-Harvest Properties. Plants 2021, 10, 2359. [Google Scholar] [CrossRef] [PubMed]
- Changwal, C.; Shukla, T.; Hussain, Z.; Singh, N.; Kar, A.; Singh, V.P.; Abdin, M.Z.; Arora, A. Regulation of Postharvest Tomato Fruit Ripening by Endogenous Salicylic Acid. Front. Plant Sci. 2021, 12, 768. [Google Scholar] [CrossRef]
- Tao, X.; Wu, Q.; Li, J.; Huang, S.; Cai, L.; Mao, L.; Luo, Z.; Li, L.; Ying, T. Exogenous methyl jasmonate regulates phenolic compounds biosynthesis during postharvest tomato ripening. Postharvest Biol. Technol. 2022, 184, 111760. [Google Scholar] [CrossRef]
- Torun, H.; Uluisik, S. Postharvest application of hydrogen peroxide affects physicochemical characteristics of tomato fruit during storage. Hortic. Environ. Biotechnol. 2022, 63, 391–401. [Google Scholar] [CrossRef]
- Zhang, B.; Tieman, D.M.; Jiao, C.; Xu, Y.; Chen, K.; Fei, Z.; Giovannoni, J.J.; Klee, H.J. Chilling-induced tomato flavor loss is associated with altered volatile synthesis and transient changes in DNA methylation. Proc. Natl. Acad. Sci. USA 2016, 113, 12580–12585. [Google Scholar] [CrossRef]
- Zuccarelli, R.; Rodríguez-Ruiz, M.; Lopes-Oliveira, P.J.; Pascoal, G.B.; Andrade, S.C.; Furlan, C.M.; Purgatto, E.; Palma, J.M.; Corpas, F.J.; Rossi, M. Multifaceted roles of nitric oxide in tomato fruit ripening: NO-induced metabolic rewiring and consequences for fruit quality traits. J. Exp. Bot. 2021, 72, 941–958. [Google Scholar] [CrossRef]
- Xu, F.; Wang, H.; Tang, Y.; Dong, S.; Qiao, X.; Chen, X.; Zheng, Y. Effect of 1-methylcyclopropene on senescence and sugar metabolism in harvested broccoli florets. Postharvest Biol. Technol. 2016, 116, 45–49. [Google Scholar] [CrossRef]
- Yao, S.; Cao, Q.; Xie, J.; Deng, L.; Zeng, K. Alteration of sugar and organic acid metabolism in postharvest granulation of Ponkan fruit revealed by transcriptome profiling. Postharvest Biol. Technol. 2018, 139, 2–11. [Google Scholar] [CrossRef]
- Hu, K.-D.; Wang, Q.; Hu, L.-Y.; Gao, S.-P.; Wu, J.; Li, Y.-H.; Zheng, J.-L.; Han, Y.; Liu, Y.-S.; Zhang, H. Hydrogen sulfide prolongs postharvest storage of fresh-cut pears (Pyrus pyrifolia) by alleviation of oxidative damage and inhibition of fungal growth. PLoS ONE 2014, 9, e85524. [Google Scholar] [CrossRef] [PubMed]
- Osorio, S.; Ruan, Y.-L.; Fernie, A.R. An update on source-to-sink carbon partitioning in tomato. Front. Plant Sci. 2014, 5, 516. [Google Scholar] [CrossRef]
- Wang, L.; Chen, Y.; Wang, S.; Xue, H.; Su, Y.; Yang, J.; Li, X. Identification of candidate genes involved in the sugar metabolism and accumulation during pear fruit post-harvest ripening of ‘Red Clapp’s Favorite’(Pyrus communis L.) by transcriptome analysis. Hereditas 2018, 155, 1–10. [Google Scholar] [CrossRef]
- Vimolmangkang, S.; Zheng, H.; Peng, Q.; Jiang, Q.; Wang, H.; Fang, T.; Liao, L.; Wang, L.; He, H.; Han, Y. Assessment of sugar components and genes involved in the regulation of sucrose accumulation in peach fruit. J. Agric. Food Chem. 2016, 64, 6723–6729. [Google Scholar] [CrossRef] [PubMed]
- Vinha, A.; Alves, R.; Barreira, S.; Castro, A.; Costa, A.; Oliveira, M. Effect of peel and seed removal on the nutritional value and antioxidant activity of tomato (Lycopersicon esculentum L.) fruits. LWT–Food Sci. Technol. 2014, 55, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Zhong, T.-Y.; Yao, G.-F.; Wang, S.-S.; Li, T.-T.; Sun, K.-K.; Tang, J.; Huang, Z.-Q.; Yang, F.; Li, Y.-H.; Chen, X.-Y. Hydrogen sulfide maintains good nutrition and delays postharvest senescence in postharvest tomato fruit by regulating antioxidative metabolism. J. Plant Growth Regul. 2021, 40, 2548–2559. [Google Scholar] [CrossRef]
- Petro-Turza, M. Flavor of tomato and tomato products. Food Rev. Int. 1986, 2, 309–351. [Google Scholar] [CrossRef]
- Tieman, D.; Bliss, P.; McIntyre, L.M.; Blandon-Ubeda, A.; Bies, D.; Odabasi, A.Z.; Rodríguez, G.R.; van der Knaap, E.; Taylor, M.G.; Goulet, C. The chemical interactions underlying tomato flavor preferences. Curr. Biol. 2012, 22, 1035–1039. [Google Scholar] [CrossRef]
- Vogel, J.T.; Tieman, D.M.; Sims, C.A.; Odabasi, A.Z.; Clark, D.G.; Klee, H.J. Carotenoid content impacts flavor acceptability in tomato (Solanum lycopersicum). J. Sci. Food Agric. 2010, 90, 2233–2240. [Google Scholar] [CrossRef] [PubMed]
- Ni, Z.; Hu, K.; Song, C.; Ma, R.; Li, Z.; Zheng, J.; Fu, L.; Wei, Z.; Zhang, H. Hydrogen sulfide alleviates postharvest senescence of grape by modulating the antioxidant defenses. Oxid. Med. Cell. Longev. 2016, 2016, 4715651. [Google Scholar] [CrossRef]
- Gao, S.; Hu, K.; Hu, L.; Li, Y.; Han, Y.; Wang, H.; Lv, K.; Liu, Y.; Zhang, H. Hydrogen sulfide delays postharvest senescence and plays an antioxidative role in fresh-cut kiwifruit. HortScience 2013, 48, 1385–1392. [Google Scholar] [CrossRef]
- Cianciosi, D.; Forbes-Hernández, T.Y.; Regolo, L.; Alvarez-Suarez, J.M.; Navarro-Hortal, M.D.; Xiao, J.; Quiles, J.L.; Battino, M.; Giampieri, F. The reciprocal interaction between polyphenols and other dietary compounds: Impact on bioavailability, antioxidant capacity and other physico-chemical and nutritional parameters. Food Chem. 2022, 375, 131904. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhu, H. The regulation of nutrient and flavor metabolism in tomato fruit. Veg. Res. 2022, 2, 1–14. [Google Scholar] [CrossRef]
- Choi, S.-H.; Lee, S.-H.; Kim, H.-J.; Lee, I.-S.; Kozukue, N.; Levin, C.E.; Friedman, M. Changes in free amino acid, phenolic, chlorophyll, carotenoid, and glycoalkaloid contents in tomatoes during 11 stages of growth and inhibition of cervical and lung human cancer cells by green tomato extracts. J. Agric. Food Chem. 2010, 58, 7547–7556. [Google Scholar] [CrossRef]
- Olsen, K.M.; Lea, U.S.; Slimestad, R.; Verheul, M.; Lillo, C. Differential expression of four Arabidopsis PAL genes; PAL1 and PAL2 have functional specialization in abiotic environmental-triggered flavonoid synthesis. J. Plant Physiol. 2008, 165, 1491–1499. [Google Scholar] [CrossRef]
- Han, C.; Li, J.; Jin, P.; Li, X.; Wang, L.; Zheng, Y. The effect of temperature on phenolic content in wounded carrots. Food Chem. 2017, 215, 116–123. [Google Scholar] [CrossRef]
- Aghdam, M.S.; Mahmoudi, R.; Razavi, F.; Rabiei, V.; Soleimani, A. Hydrogen sulfide treatment confers chilling tolerance in hawthorn fruit during cold storage by triggering endogenous H2S accumulation, enhancing antioxidant enzymes activity and promoting phenols accumulation. Sci. Hortic. 2018, 238, 264–271. [Google Scholar] [CrossRef]
- Liu, C.; Zheng, H.; Sheng, K.; Liu, W.; Zheng, L. Effects of postharvest UV-C irradiation on phenolic acids, flavonoids, and key phenylpropanoid pathway genes in tomato fruit. Sci. Hortic. 2018, 241, 107–114. [Google Scholar] [CrossRef]
- Barros, L.; Dueñas, M.; Pinela, J.; Carvalho, A.M.; Buelga, C.S.; Ferreira, I.C. Characterization and quantification of phenolic compounds in four tomato (Lycopersicon esculentum L.) farmers’ varieties in northeastern Portugal homegardens. Plant Foods Hum. Nutr. 2012, 67, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, J. Determination of the content of soluble sugar in sweet corn with optimized anthrone colorimetric method. Storage Process 2013, 13, 24–27. [Google Scholar]
- Gomez, L.; Rubio, E.; Auge, M. A new procedure for extraction and measurement of soluble sugars in ligneous plants. J. Sci. Food Agric. 2002, 82, 360–369. [Google Scholar] [CrossRef]
- Gao, Q.H.; Wu, C.S.; Yu, J.G.; Wang, M.; Ma, Y.J.; Li, C.L. Textural characteristic, antioxidant activity, sugar, organic acid, and phenolic profiles of 10 promising jujube (Ziziphus jujuba Mill.) selections. J. Food Sci. 2012, 77, C1218–C1225. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, C.; Sun, J.; Jackson, A. Dynamic changes of enzymes involved in sugar and organic acid level modification during blueberry fruit maturation. Food Chem. 2020, 309, 125617. [Google Scholar] [CrossRef]
- Toor, R.K.; Savage, G.P. Antioxidant activity in different fractions of tomatoes. Food Res. Int. 2005, 38, 487–494. [Google Scholar] [CrossRef]
- Jia, Z.; Tang, M.; Wu, J. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar]
- Castillo, J.M.; Mancilla-Leytón, J.M.; Martins-Noguerol, R.; Moreira, X.; Moreno-Pérez, A.J.; Muñoz-Vallés, S.; Pedroche, J.J.; Figueroa, M.E.; García-González, A.; Salas, J.J. Interactive effects between salinity and nutrient deficiency on biomass production and bio-active compounds accumulation in the halophyte Crithmum maritimum. Sci. Hortic. 2022, 301, 111136. [Google Scholar] [CrossRef]
- Huang, D.; Li, W.; Dawuda, M.M.; Huo, J.; Li, C.; Wang, C.; Liao, W. Hydrogen sulfide reduced colour change in lanzhou lily-bulb scales. Postharvest Biol. Technol. 2021, 176, 111520. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolic Compounds (μg/mL) | H2O | H2S | HT |
|---|---|---|---|
| Rutin | 153.656 ± 0.643 b | 145.073 ± 2.412 b | 173.269 ± 6.199 a |
| Gentianic acid | 34.519 ± 0.182 b | 38.484 ± 0.215 a | 32.963 ± 0.285 c |
| Quercetin | 16.978 ± 0.17 b | 15.142 ± 0.278 c | 18.056 ± 0.260 a |
| Benzoic acid | 8.683 ± 0.138 b | 10.113 ± 0.198 a | 7.594 ± 0.108 c |
| Gallic acid | 8.484 ± 0.018 b | 9.540 ± 0.114 a | 5.829 ± 0.364 c |
| Kaempferol | 7.887 ± 0.004 b | 7.849 ± 0.002 c | 8.003 ± 0.012 a |
| Chlorogenic acid | 5.358 ± 0.002 b | 8.668 ± 0.400 a | 5.09 ± 0.025 b |
| Ferulic acid | 5.332 ± 0.197 b | 6.393 ± 0.003 a | 4.574 ± 0.042 c |
| Caffeic acid | 5.179 ± 0.0147 b | 5.277 ± 0.019 a | 5.125 ± 0.019 b |
| Cinnamic acid | 5.144 ± 0.084 a | 5.370 ± 0.072 a | 5.378 ± 0.075 a |
| Erucic acid | 5.031 ± 0.018 b | 5.923 ± 0.011 a | 4.900 ± 0.005 b |
| Cynarin | 4.993 ± 0.025 b | 5.937 ± 0.095 a | 4.960 ± 0.034 b |
| Protocatechuic acid | 4.605 ± 0.004 b | 4.636 ± 0.004 a | 4.522 ± 0.004 c |
| P-hydroxybenzoic acid | 4.552 ± 0.012 a | 4.605 ± 0.031 a | 4.458 ± 0.027 b |
| 4-Coumaric acid | 4.302 ± 0.001 b | 4.270 ± 0.014 b | 5.141 ± 0.148 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Yun, F.; Man, X.; Huang, D.; Liao, W. Effects of Hydrogen Sulfide on Sugar, Organic Acid, Carotenoid, and Polyphenol Level in Tomato Fruit. Plants 2023, 12, 719. https://doi.org/10.3390/plants12040719
Zhang Y, Yun F, Man X, Huang D, Liao W. Effects of Hydrogen Sulfide on Sugar, Organic Acid, Carotenoid, and Polyphenol Level in Tomato Fruit. Plants. 2023; 12(4):719. https://doi.org/10.3390/plants12040719
Chicago/Turabian StyleZhang, Yanqin, Fahong Yun, Xiaoling Man, Dengjing Huang, and Weibiao Liao. 2023. "Effects of Hydrogen Sulfide on Sugar, Organic Acid, Carotenoid, and Polyphenol Level in Tomato Fruit" Plants 12, no. 4: 719. https://doi.org/10.3390/plants12040719