
Growth-Promoting Characteristics of Fungal and Bacterial Endophytes Isolated from a Drought-Tolerant Mint Species Endostemon obtusifolius (E. Mey. ex Benth.) N. E. Br
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Sterilization, Isolation, and Purification of Endophytes
2.2. Total Phenolic and Flavonoid Content Estimation
2.3. DPPH Radical Scavenging Abilities
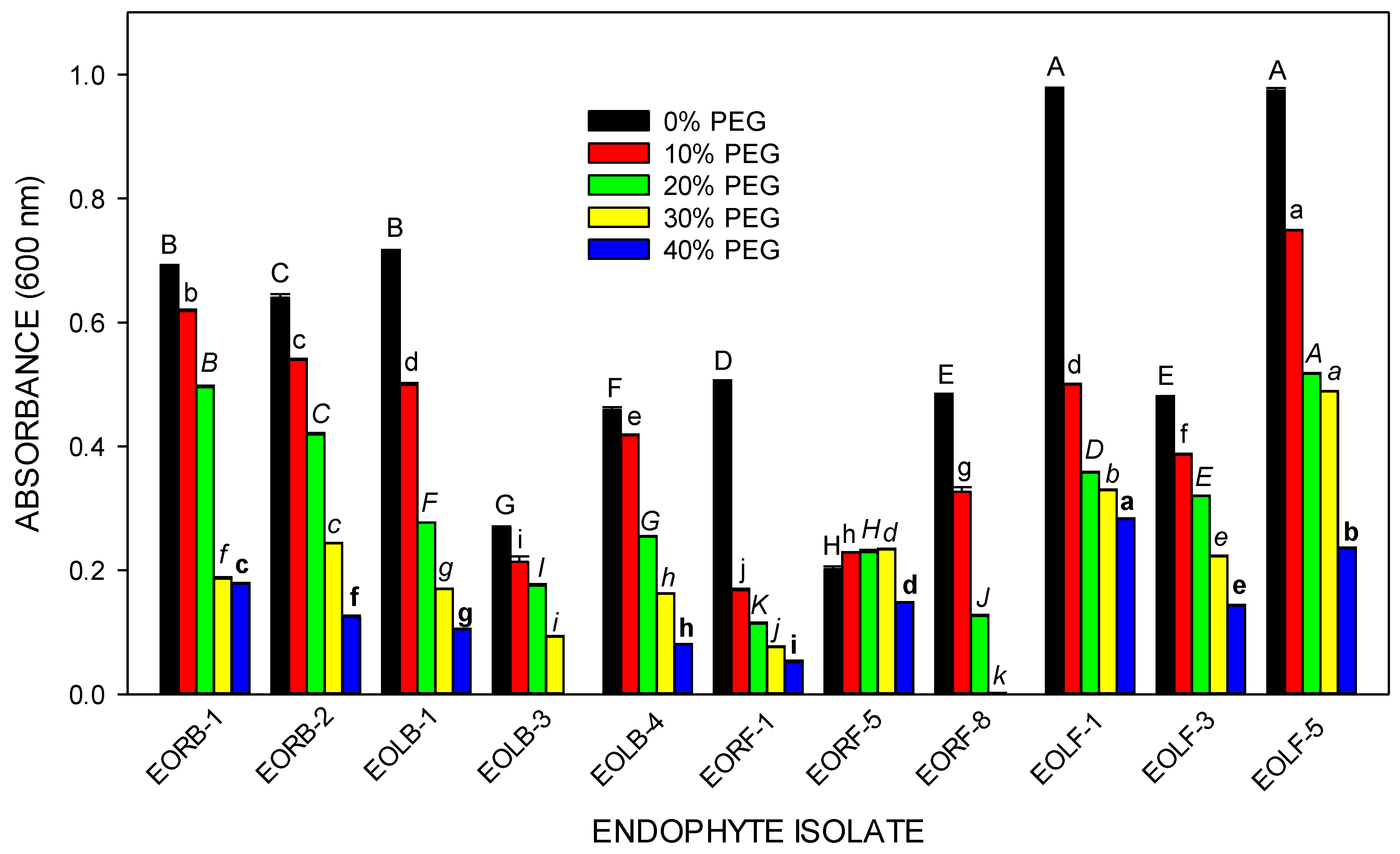
2.4. The Water-Deficit Resistance Potential of Selected Endophytes
2.5. Plant Growth-Promoting Characteristics of Selected Endophytic Isolates
2.6. Molecular Identification of Endophytes
2.7. Endophytes Antagonistic Check Using Dual Culture Method
3. Discussion
4. Materials and Methods
4.1. Plant Sample Collection
4.2. Surface Sterilization and Isolation of Endophytes
4.3. Extraction and Quantification of Secondary Metabolites from Endophytes
4.4. Antioxidant Activity of Crude Extracts from Endophytes
4.5. Screening of Endophytic Isolates for Drought Stress Tolerance
4.6. Evaluation of Plant Growth-Promoting Characteristics of Selected Endophytic Isolates
4.6.1. Ammonia Production
4.6.2. Siderophore Production
4.6.3. HCN Production
4.6.4. Phosphate Solubilization Activity
4.6.5. IAA Production
4.6.6. ACCD Activity
4.7. Molecular Identification of Endophytes
4.8. Endophyte Antagonistic Check Using Dual Culture Method
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Imadi, S.R.; Kazi, A.G.; Hashem, A.; Abd-Allah, E.F.; Alqarawi, A.A.; Ahmad, P. Medicinal plants under abiotic stress. In Plant-Environment Interaction: Responses and Approaches to Mitigate Stress; Azooz, M.M., Ahmad, P., Eds.; John Wiley and Sons: Hoboken, NJ, USA, 2015; pp. 300–310. [Google Scholar]
- Ramakrishna, A.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [PubMed]
- Joetzjer, E.; Delire, C.; Douville, H.; Ciais, P.; Decharme, B.; Fisher, R.; Christoffersen, B.; Calvet, J.C.; Da Costa, A.C.L.; Ferreira, L.V.; et al. Predicting the response of the Amazon rainforest to persistent drought conditions under current and future climates: A major challenge for global land surface models. Geosci. Model. Dev. 2014, 7, 2933–2950. [Google Scholar] [CrossRef]
- Dikilitas, M.; Karakas, S.; Hashem, A.; Abd Allah, E.F.; Ahmad, P. Oxidative stress and plant responses to pathogens under drought conditions. In Water Stress and Crop Plants: A Sustainable Approach, 1st ed.; Ahmed, P., Ed.; John Wiley and Sons: Hoboken, NJ, USA, 2016; pp. 102–123. [Google Scholar]
- Jaleel, C.A.; Manivannan, P.; Lakshmanan, G.M.A.; Gomathinayagam, M.; Panneerselvam, R. Alterations in morphological parameters and photosynthetic pigment responses of Catharanthus roseus under soil water deficits. Colloids Surf. B Biointerfaces 2008, 61, 298–303. [Google Scholar] [CrossRef]
- Ogbe, A.A.; Finnie, J.F.; Van Staden, J. The role of endophytes in secondary metabolites accumulation in medicinal plants under abiotic stress. S. Afr. J. Bot. 2020, 134, 126–134. [Google Scholar] [CrossRef]
- Silva, C.F.; Vitorino, L.C.; Mendonça, M.A.C.; Araújo, W.L.; Dourado, M.N.; Albuquerque, L.C.; Soares, M.A.; Souchie, E.L. Screening of plant growth-promoting endophytic bacteria from the roots of the medicinal plant Aloe vera. S. Afr. J. Bot. 2020, 134, 3–16. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.J.; Shurigin, V.V.; Hashem, A.; Abd Allah, E.F. Endophytic bacteria improve plant growth, symbiotic performance of chickpea (Cicer arietinum L.) and induce suppression of root rot caused by Fusarium solani under salt stress. Front. Microbiol. 2017, 8, 1887. [Google Scholar] [CrossRef]
- Nair, D.N.; Padmavathy, S. Impact of endophytic microorganisms on plants, environment, and humans. Sci. World J. 2014, 2014, 250693. [Google Scholar] [CrossRef]
- Lee, K.; Missaoui, A.; Mahmud, K.; Presley, H.; Lonnee, M. Interaction between grasses and Epichloë endophytes and its significance to biotic and abiotic stress tolerance and the rhizosphere. Microorganisms 2021, 9, 2186. [Google Scholar] [CrossRef]
- Naveed, M.; Hussain, M.B.; Zahir, Z.A.; Mitter, B.; Sessitsch, A. Drought stress amelioration in wheat through inoculation with Burkholderia phytofirmans strain PsJN. Plant Growth Regul. 2014, 73, 121–131. [Google Scholar] [CrossRef]
- Wicaksono, W.A.; Eirian Jones, E.; Monk, J.; Ridgway, H.J. Using bacterial endophytes from a New Zealand native medicinal plant for control of grapevine trunk diseases. Biol. Control 2017, 114, 65–72. [Google Scholar] [CrossRef]
- Nimaichand, S.; Devi, A.M.; Li, W.J. Direct plant growth-promoting ability of actinobacteria in grain legumes. In Plant Growth Promoting Actinobacteria: A New Avenue for Enhancing the Productivity and Soil Fertility of Grain Legumes; Subramaniam, G., Arumugam, S., Rajendran, V., Eds.; Springer: Singapore, 2016; pp. 1–298. [Google Scholar]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiol. Res. 2016, 184, 13–24. [Google Scholar] [CrossRef]
- Vurukonda, S.S.K.P.; Giovanardi, D.; Stefani, E. Plant growth promoting and biocontrol activity of Streptomyces spp. As endophytes. Int. J. Mol. Sci. 2018, 19, 952. [Google Scholar] [CrossRef] [PubMed]
- Yaish, M.W.; Antony, I.; Glick, B.R.; van Leeuwenhoek, A. Isolation and characterization of endophytic plant growth-promoting bacteria from a date palm tree (Phoenix dactylifera L.) and their potential role in salinity tolerance. Int. J. Mol. Microbiol. 2015, 107, 1519–1532. [Google Scholar] [CrossRef]
- Lin, L.; Xu, X. Indole-3-acetic acid production by endophytic Streptomyces sp. EN-1 isolated from medicinal plants. Curr. Microbiol. 2013, 67, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Kundan, R.; Pant, G. Plant growth promoting rhizobacteria: Mechanism and Current Prospective. J. Biofert. Biopest. 2015, 6, 9. [Google Scholar] [CrossRef]
- Figueiredo, J.E.F.; Gomes, E.A.; Guimarães, C.T.; Lana, U.d.P.; Teixeira, M.A.; Lima, G.V.C.; Bressan, W. Molecular analysis of endophytic bacteria from the genus Bacillus isolated from tropical maize (Zea mays L.). Braz. J. Microbiol. 2009, 40, 522–534. [Google Scholar] [CrossRef]
- Marwat, S.K.; Fazal-Ur-Rehman Khan, M.S.; Ghulam, S.; Anwar, N.; Mustafa, G.; Usman, K. Phytochemical constituents and pharmacological activities of sweet Basil—Ocimum basilicum L. (Lamiaceae). Asian J. Chem. 2011, 23, 3773–3782. [Google Scholar]
- Orhan, I.; Kartal, M.; Naz, Q.; Ejaz, A.; Yilmaz, G.; Kan, Y.; Konuklugil, B.; Şener, B.; Iqbal Choudhary, M. Antioxidant and anticholinesterase evaluation of selected Turkish Salvia species. Food Chem. 2007, 103, 1247–1254. [Google Scholar] [CrossRef]
- Sadashiva, C.T.; Naidoo, Y.; Naidoo, J.R.; Kalicharan, B.; Naidoo, G. Antioxidant and acetylcholinesterase activities of three species of the family Lamiaceae. Bangladesh J. Bot. 2014, 43, 331–335. [Google Scholar] [CrossRef]
- Hassan, S.E.D. Plant growth-promoting activities for bacterial and fungal endophytes isolated from medicinal plant of Teucrium polium L. J. Adv. Res. 2017, 8, 687–695. [Google Scholar] [CrossRef]
- You, C.; Qin, D.; Wang, Y.; Lan, W.; Li, Y.; Yu, B.; Peng, Y.; Xu, J.; Dong, J. Plant triterpenoids regulate endophyte community to promote medicinal plant Schisandra sphenanthera Growth and Metabolites Accumulation. J. Fungi 2021, 7, 788. [Google Scholar] [CrossRef]
- Tan, R.X.; Zou, W.X. Endophytes: A rich source of functional metabolites. Nat. Prod. Rep. 2001, 18, 448–459. [Google Scholar] [CrossRef] [PubMed]
- Kaur, N.; Arora, D.S.; Kalia, N.; Kaur, M. Bioactive potential of endophytic fungus Chaetomium globosum and GC–MS analysis of its responsible components. Sci. Rep. 2020, 10, 18792. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.C.; Goveas, S.; D’Souza, L.; D’Cunha, F.; D’Souza, V. Antibacterial and antioxidant potential of organic solvents extracts of Mangrove endophytic fungus Eupenicillium senticosum Scott. J. Alternat. Med. Res. 2017, 9, 65–73. [Google Scholar]
- Yadav, M.; Yadav, A.; Yadav, J.P. In vitro antioxidant activity and total phenolic content of endophytic fungi isolated from Eugenia jambolana Lam. Asian Pac. J. Trop. Med. 2014, 7, S256–S261. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cheng, C.; An, D. Characterization of endophytic bacteria from a desert plant Lepidium perfoliatum L. Plant Prot. Sci. 2017, 53, 32–43. [Google Scholar] [CrossRef]
- Ripa, F.A.; Cao, W.D.; Tong, S.; Sun, J.G. Assessment of plant growth promoting and abiotic stress tolerance properties of wheat endophytic fungi. Biomed. Res. Int. 2019, 2019, 6105865. [Google Scholar] [CrossRef]
- Malakar, P.; Singh, V.K.; Karmakar, R.; Venkatesh, K.V. Effect on β-galactosidase synthesis and burden on growth of osmotic stress in Escherichia coli. SpringerPlus 2014, 3, 748. [Google Scholar] [CrossRef]
- Liu, Z.; Fu, B.; Zheng, X.; Liu, G. Plant biomass, soil water content and soil N:P ratio regulating soil microbial functional diversity in a temperate steppe: A regional scale study. Soil Biol. Biochem. 2010, 42, 445–450. [Google Scholar] [CrossRef]
- Zak, D.R.; Holmes, W.E.; White, D.C.; Peacock, A.D.; Tilman, D.; Zak, D.R.; Holmes, W.E.; White, D.C.; Peacock, A.D.; Tilman, D. Plant diversity, soil microbial communities, and ecosystem function: Are there any links ? Ecology 2016, 84, 2042–2050. [Google Scholar] [CrossRef]
- Rolli, E.; Marasco, R.; Vigani, G.; Ettoumi, B.; Mapelli, F.; Deangelis, M.L.; Gandolfi, C.; Casati, E.; Previtali, F.; Gerbino, R.; et al. Improved plant resistance to drought is promoted by the root-associated microbiome as a water stress-dependent trait. Environ. Microbiol. 2015, 17, 316–331. [Google Scholar] [CrossRef] [PubMed]
- Vandana, U.K.; Rajkumari, J.; Singha, L.P.; Satish, L.; Alavilli, H.; Sudheer, P.D.V.N.; Chauhan, S.; Ratnala, R.; Satturu, V.; Mazumder, P.B.; et al. The endophytic microbiome as a hotspot of synergistic interactions, with prospects of plant growth promotion. Biology 2021, 10, 101. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, E.K.; Jeon, J.; Rim, S.O.; Park, Y.H.; Kyu Lee, S.; Bae, H. Composition, diversity, and bioactivity of culturable bacterial endophytes in mountain-cultivated ginseng in Korea. Sci. Rep. 2017, 7, 10098. [Google Scholar] [CrossRef] [PubMed]
- Oteino, N.; Lally, R.D.; Kiwanuka, S.; Lloyd, A.; Ryan, D.; Germaine, K.J.; Dowling, D.N. Plant growth promotion induced by phosphate solubilizing endophytic Pseudomonas isolates. Front. Microbiol. 2015, 6, 745. [Google Scholar] [CrossRef] [PubMed]
- Scagliola, M.; Pii, Y.; Mimmo, T.; Cesco, S.; Ricciuti, P.; Crecchio, C. Characterization of plant growth promoting traits of bacterial isolates from the rhizosphere of barley (Hordeum vulgare L.) and tomato (Solanum lycopersicon L.) grown under Fe sufficiency and deficiency. Plant Physiol. Biochem. 2016, 107, 187–196. [Google Scholar] [CrossRef]
- Ahmad, F.; Ahmad, I.; Khan, M.S. Indole acetic acid production by the indigenous isolates of Azotobacter and fluorescent Pseudomonas in the presence and absence of tryptophan. Turk. J. Biol. 2005, 29, 29–34. [Google Scholar]
- Maheshwari, R.; Bhutani, N.; Suneja, P. Isolation and characterization of ACC deaminase producing endophytic Bacillus mojavensis PRN2 from Pisum sativum. Iran. J. Biotechnol. 2020, 18, 11–20. [Google Scholar]
- Rauf, M.; Awais, M.; Ud-Din, A.; Ali, K.; Gul, H.; Rahman, M.M.; Hamayun, M.; Arif, M. Molecular mechanisms of the 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase producing Trichoderma asperellum MAP1 in enhancing wheat tolerance to waterlogging stress. Front. Plant Sci. 2021, 11, 614971. [Google Scholar] [CrossRef]
- Dubey, A.; Saiyam, D.; Kumar, A.; Hashem, A.; Abduallah, E.F.; Khan, M.L. Bacterial root endophytes: Characterization of their competence and plant growth promotion in soybean (Glycine max (L.) Merr.) under drought stress. Int. J. Environ. Res. Public Health 2021, 18, 931. [Google Scholar] [CrossRef]
- Chukwuneme, C.F.; Babalola, O.O.; Kutu, F.R.; Ojuederie, O.B. Characterization of actinomycetes isolates for plant growth promoting traits and their effects on drought tolerance in maize. J. Plant Interact. 2020, 15, 93–105. [Google Scholar] [CrossRef]
- Sapre, S.; Gontia-Mishra, I.; Tiwari, S. ACC deaminase-producing bacteria: A key player in alleviating abiotic stresses in plants. In Plant Growth Promoting Rhizobacteria for Agricultural Sustainability; Kumar, A., Meena, V., Eds.; Springer: Singapore, 2019; pp. 267–291. [Google Scholar]
- Glick, B.R. Bacterial ACC deaminase and the alleviation of plant stress. Adv. Appl. Microbiol. 2004, 56, 291–312. [Google Scholar] [PubMed]
- Katoch, M.; Pull, S. Endophytic fungi associated with Monarda citriodora, an aromatic and medicinal plant and their biocontrol potential. Pharm. Biol. 2017, 55, 1528–1535. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, D.; Manna, S.; Mahapatra, S.; Pati, B.R. Fungal endophytes in three medicinal plants of Lamiaceae. Acta Microbiol. Immunol. Hung. 2009, 56, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Xu, Y.; Lai, X.H.; Shan, C.; Deng, Z.; Ji, Y. Screening and characterization of endophytic Bacillus and Paenibacillus strains from medicinal plant Lonicera japonica for use as potential plant growth promoters. Braz. J. Microbiol. 2015, 46, 977–989. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Gao, J.; Chen, X.; Zhang, M.; Yang, F.; Du, Y.; Moe, T.S.; Munir, I.; Xue, J.; Zhang, X. Isolation and characterization of plant growth-promoting endophytic bacteria Paenibacillus polymyxa SK1 from Lilium lancifolium. Biomed. Res. Int. 2020, 2020, 8650957. [Google Scholar] [CrossRef]
- Ghiasvand, M.; Makhdoumi, A.; Matin, M.M.; Vaezi, J. Exploring the bioactive compounds from endophytic bacteria of a medicinal plant: Ephedra foliata (Ephedrales: Ephedraceae). Orient. Pharm. Exp. Med. 2019, 20, 61–70. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, Q.; Zang, P.; Li, X.; Ji, Q.; He, Z.; Zhao, Y.; Yang, H.; Zhao, X.; Zhang, L. An endophytic bacterium isolated from Panax ginseng C.A. Meyer enhances growth, reduces morbidity, and stimulates ginsenoside biosynthesis. Phytochem. Lett. 2015, 11, 132–138. [Google Scholar] [CrossRef]
- Wang, X.M.; Yang, B.; Wang, H.W.; Yang, T.; Ren, C.G.; Zheng, H.L.; Dai, C.C. Consequences of antagonistic interactions between endophytic fungus and bacterium on plant growth and defense responses in Atractylodes lancea. J. Basic Microbiol. 2015, 55, 659–670. [Google Scholar] [CrossRef]
- Kia, S.H.; Pallesch, S.; Piepenbring, M.; Maciá-Vicente, J.G. Root endophytic fungi show low levels of interspecific competition in planta. Fungal Ecol. 2019, 39, 184–191. [Google Scholar] [CrossRef]
- Khan, M.S.; Gao, J.; Zhang, M.; Chen, X.; Moe, T.S.; Du, Y.; Yang, F.; Xue, J.; Zhang, X. Isolation and characterization of plant growth-promoting endophytic bacteria Bacillus stratosphericus LW-03 from Lilium wardii. 3Biotech 2020, 10, 305. [Google Scholar] [CrossRef]
- Timmusk, S.; Copolovici, D.; Copolovici, L.; Teder, T.; Nevo, E.; Behers, L. Paenibacillus polymyxa biofilm polysaccharides antagonize Fusarium graminearum. Sci. Rep. 2019, 9, 662. [Google Scholar] [CrossRef] [PubMed]
- Jimtha, J.C.; Smitha, P.V.; Anisha, C.; Deepthi, T.; Meekha, G.; Radhakrishnana, E.K.; Gayatri, G.P.; Remakanthan, A. Isolation of endophytic bacteria from embryogenic suspension culture of banana and assessment of their plant growth promoting properties. Plant Cell Tiss. Org. Cult. 2014, 118, 57–66. [Google Scholar] [CrossRef]
- Khan, A.R.; Ullah, I.; Waqas, M.; Shahzad, R.; Hong, S.J.; Park, G.S.; Jung, B.K.; Lee, I.J.; Shin, J.H. Plant growth-promoting potential of endophytic fungi isolated from Solanum nigrum leaves. World J. Microbiol. Biotechnol. 2015, 31, 1461–1466. [Google Scholar] [CrossRef] [PubMed]
- Higginbotham, S.J.; Arnold, A.E.; Ibañez, A.; Spadafora, C.; Coley, P.D.; Kursar, T.A. Bioactivity of fungal endophytes as a function of endophyte taxonomy and the taxonomy and distribution of their host plants. PLoS ONE 2013, 9, e73192. [Google Scholar] [CrossRef]
- Deljou, A.; Goudarzi, S. Green extracellular synthesis of the silver nanoparticles using thermophilic Bacillus sp. AZ1 and its antimicrobial activity against several human pathogenetic bacteria. Iran. J. Biotechnol. 2016, 14, 25. [Google Scholar] [CrossRef]
- Dragović-Uzelac, V.; Levaj, B.; Bursać, D.; Pedisić, S.; Radojčić, I.; Biško, A. Total phenolics and antioxidant capacity assays of selected fruits. Agric. Conspec. Sci. 2007, 72, 279–284. [Google Scholar]
- Sridhar, K.; Charles, A.L. In vitro antioxidant activity of Kyoho grape extracts in DPPH (rad) and ABTS (rad) assays: Estimation methods for EC50 using advanced statistical programs. Food Chem. 2019, 275, 41–49. [Google Scholar] [CrossRef]
- Eke, P.; Kumar, A.; Sahu, K.P.; Wakam, L.N.; Sheoran, N.; Ashajyothi, M.; Patel, A.; Fekam, F.B. Endophytic bacteria of desert cactus (Euphorbia trigonas Mill) confer drought tolerance and induce growth promotion in tomato (Solanum lycopersicum L.). Microbiol. Res. 2019, 228, 126302. [Google Scholar] [CrossRef]
- Milagres, A.M.F.; Machuca, A.; Napoleão, D. Detection of siderophore production from several fungi and bacteria by a modification of Chrome Azurol S (CAS) agar plate assay. J. Microbiol. Methods 1999, 37, 1–6. [Google Scholar] [CrossRef]
- Bakker, A.W.; Schippers, B. Microbial cyanide production in the rhizosphere in relation to potato yield reduction and Pseudomonas spp.-mediated plant growth-stimulation. Soil Biol. Biochem. 1987, 19, 451–457. [Google Scholar] [CrossRef]
- Yadav, R.; Singh, A.V.; Kumar, M.; Yadav, S. Phytochemical analysis and plant growth promoting properties of endophytic fungi isolated from tulsi and Aloe vera. Int. J. Agric. Stat. Sci. 2016, 12, 239–248. [Google Scholar]
- Edi Premono, M.; Moawad, A.M.; Vlek, P.L.G. Effect of phosphate-solubilizing Pseudomonas putida on the growth of maize and its survival in the rhizosphere. Indones. J. Agric. Sci. 1996, 11, 13–23. [Google Scholar]
- Chand, K.; Shah, S.; Sharma, J.; Paudel, M.R.; Pant, B. Isolation, characterization, and plant growth-promoting activities of endophytic fungi from a wild orchid Vanda cristata. Plant Signal. Behav. 2020, 15, 1744294. [Google Scholar] [CrossRef] [PubMed]
- Jasim, B.; John Jimtha, C.; Jyothis, M.; Radhakrishnan, E.K. Plant growth promoting potential of endophytic bacteria isolated from Piper nigrum. Plant Growth Regul. 2013, 71, 1–11. [Google Scholar] [CrossRef]
- Naik, B.S.; Shashikala, J.; Krishnamurthy, Y.L.A. Study on the diversity of endophytic communities from rice (Oryza sativa L.) and their antagonistic activities in vitro. Microbiol. Res. 2009, 164, 290–296. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Endophyte Isolates | Phenolics (mg GAE/mg DW of Extracts) | Flavonoids (mg CE/mg DW of Extracts) | % DPPH Radical Scavenging Abilities at 100 µg/mL | DPPH IC50 (µg/mL) |
|---|---|---|---|---|
| EORB-1 | 4.00 ± 0.02 c | 27.66 ± 0.72 de | 75.52 ± 1.13 de | 48.95 ± 1.09 hij |
| EORB-2 | 4.22 ± 0.05 c | 32.26 ± 1.10 cd | 81.90 ± 0.79 bc | 35.68 ± 0.87 ij |
| EORB-3 | - | 16.582 ± 0.60 ghijk | 10.00 ± 1.08 o | 707.48 ± 10.89 b |
| EORB-4 | - | 17.21 ± 1.27 ghijk | 20.86 ± 2.94 m | 256.26 ± 59.23 cd |
| EORB-5 | - | 18.04 ± 0.75 ghij | 25.86 ± 2.88 kl | 195.03 ± 18.34 def |
| EORB-6 | - | 16.08 ± 0.75 ghijk | 20.86 ± 1.58 m | 245.47 ± 24.07 cde |
| EOLB-1 | 1.55 0.08 g | 19.30 ± 0.21 ghi | 68.27 ± 1.77 fg | 70.76 ± 2.00 ghij |
| EOLB-2 | 0.28 ± 0.02 k | 14.28 ± 1.46 hijklm | 15.49 ± 0.66 n | 245.30 ± 2.79 cde |
| EOLB-3 | 0.27 ± 0.23 k | 10.73 1.00 klmno | 67.37 ± 0.66 gh | 66.22 ± 1.23 ghij |
| EOLB-4 | 2.00 ± 0.05 f | 56. 918 ± 6.90 b | 80.30 ± 0.84 cd | 32.37 ± 0.84 ij |
| EOLB-5 | - | 7.60 ± 0.72 no | 12.18 ± 1.31 no | 352.41 ± 20.74 c |
| EOLB-6 | - | 19.72 ± 0.21 ghi | 24.81 ± 1.08 lm | 215.36 ± 14.17 def |
| EOLB-7 | - | 12.611 ± 1.90 jklmn | 35.94 ± 0.94 j | 136.79 ± 7.17 j |
| EOLB-9 | - | 11.57 ± 1.30 jklmn | 41.20 ± 0.40 i | 118.19 ± 2.65 i |
| EORF-1 | 6.74 ± 0.30 b | 69.04 ± 2.83 a | 90.38 ± 0.40 a | 23.43 ± 0.18 ij |
| EORF-3 | 3.70 ± 0.05 d | 38.00 ± 6.88 c | 23.76 ± 4.06 lm | 222.08 ± 42.44 def |
| EORF-4 | - | 13.45 ± 1.50 ijklmn | 23.16 ± 1.59 lm | 221.44 ± 22.07 def |
| EORF-5 | 2.80 ± 0.02 e | 16.60 ± 0.42 ghijk | 66.62 ± 2.77 gh | 62.57 ± 1.65 hij |
| EORF-6 | - | 16.60 1.80 ghijk | 21.05 ± 1.30 lm | 248.67 ± 19.55 cd |
| EORF-8 | 2.12 ± 0.02 f | 26.82 ± 2.54 def | 64.36 ± 1.45 gh | 64.63 ± 2.067 ghij |
| EORF-9 | 0.62 ± 0.02 j | 17.63 ± 0.63 ghij | 41.05 ± 3.26 i | 147.44 ± 10.74 defgh |
| EOLF-1 | 1.35 ± 0.07 h | 21.60 ± 4.00 efg | 63.10 ± 1.24 h | 57.31 ± 1.25 ij |
| EOLF-2 | - | 8.43 ± 0.60 mno | 29.83 ± 1.21 k | 172.12 ± 6.08 defg |
| EOLF-3 | 1.00 ± 0.04 h | 20.76 ± 0.00 fgh | 72.24 ± 1.64 ef | 47.77 ± 0.68 hij |
| EOLF-4 | - | 9.69 ± 0.21 lmno | 22.93 ± 1.08 lm | 224.78 ± 12.34 def |
| EOLF-5 | 9.80 ± 0.03 a | 15.33 ± 0.80 ghijkl | 86.44 ± 1.88 ab | 26. 67 ± 0.889 j |
| BHT | - | - | 86.21 ± 1.53 ab | 42.98 ± 3.86 hij |
| Traits | Endophyte Isolates | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| EORB-1 | EORB-2 | EOLB-1 | EOLB-3 | EOLB-4 | EORF-1 | EORF-5 | EORF-8 | EOLF-1 | EOLF-3 | EOLF-5 | |
| Ammonia production | + | +++ | +++ | + | +++ | +++ | ++ | + | ++ | ++ | +++ |
| Siderophore production | ++ | +++ | +++ | + | + | ++ | ++ | + | +++ | ++ | +++ |
| Hydrogen cyanide production | + | +++ | + | - | + | + | + | + | ++ | ++ | ++ |
| Phosphate solubilization | + | + | + | - | + | + | + | + | + | + | + |
| IAA production | + | + | + | - | + | + | - | - | - | + | + |
| ACC deaminase activity | + | + | + | + | + | - | - | - | - | + | + |
| Isolate Code | Most Closely Related Homologue Sequence (Accession Number) | Sequence Identity (%) | GeneBank Accession Number |
|---|---|---|---|
| EORB-2 | Paenibacillus polymyxa (MT163461.1) | 100.00 | OL619995 |
| EOLF-5 | Fusarium oxysporum (MT560381.1) | 99.60 | MZ598577 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogbe, A.A.; Gupta, S.; Stirk, W.A.; Finnie, J.F.; Van Staden, J. Growth-Promoting Characteristics of Fungal and Bacterial Endophytes Isolated from a Drought-Tolerant Mint Species Endostemon obtusifolius (E. Mey. ex Benth.) N. E. Br. Plants 2023, 12, 638. https://doi.org/10.3390/plants12030638
Ogbe AA, Gupta S, Stirk WA, Finnie JF, Van Staden J. Growth-Promoting Characteristics of Fungal and Bacterial Endophytes Isolated from a Drought-Tolerant Mint Species Endostemon obtusifolius (E. Mey. ex Benth.) N. E. Br. Plants. 2023; 12(3):638. https://doi.org/10.3390/plants12030638
Chicago/Turabian StyleOgbe, Abdulazeez A., Shubhpriya Gupta, Wendy A. Stirk, Jeffrey F. Finnie, and Johannes Van Staden. 2023. "Growth-Promoting Characteristics of Fungal and Bacterial Endophytes Isolated from a Drought-Tolerant Mint Species Endostemon obtusifolius (E. Mey. ex Benth.) N. E. Br" Plants 12, no. 3: 638. https://doi.org/10.3390/plants12030638