
Natural Bioactive Substances in Fruits of Aronia melanocarpa (Michx.) Elliott Exposed to Combined Light-Type, Chitosan Oligosaccharide, and Spent Mushroom Residue Treatments

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site and Experiment Layout
2.2. LED Light Treatment and Layout
2.3. SMR Amendment Design and Layout
2.4. CO Addition Treamtent
2.5. Sampling and Analysis
2.6. Statistics and Data Analysis
3. Results and Discussion
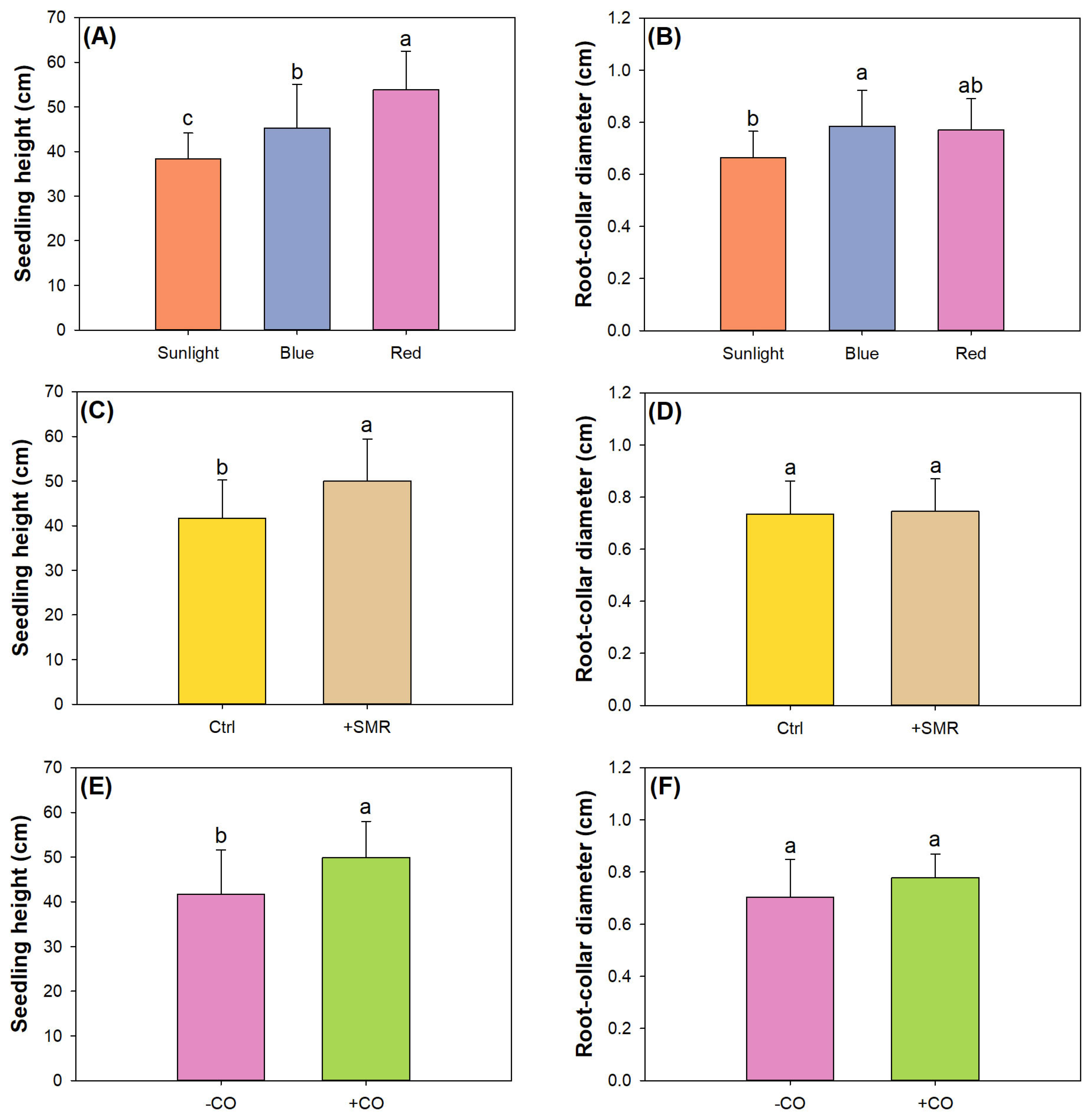
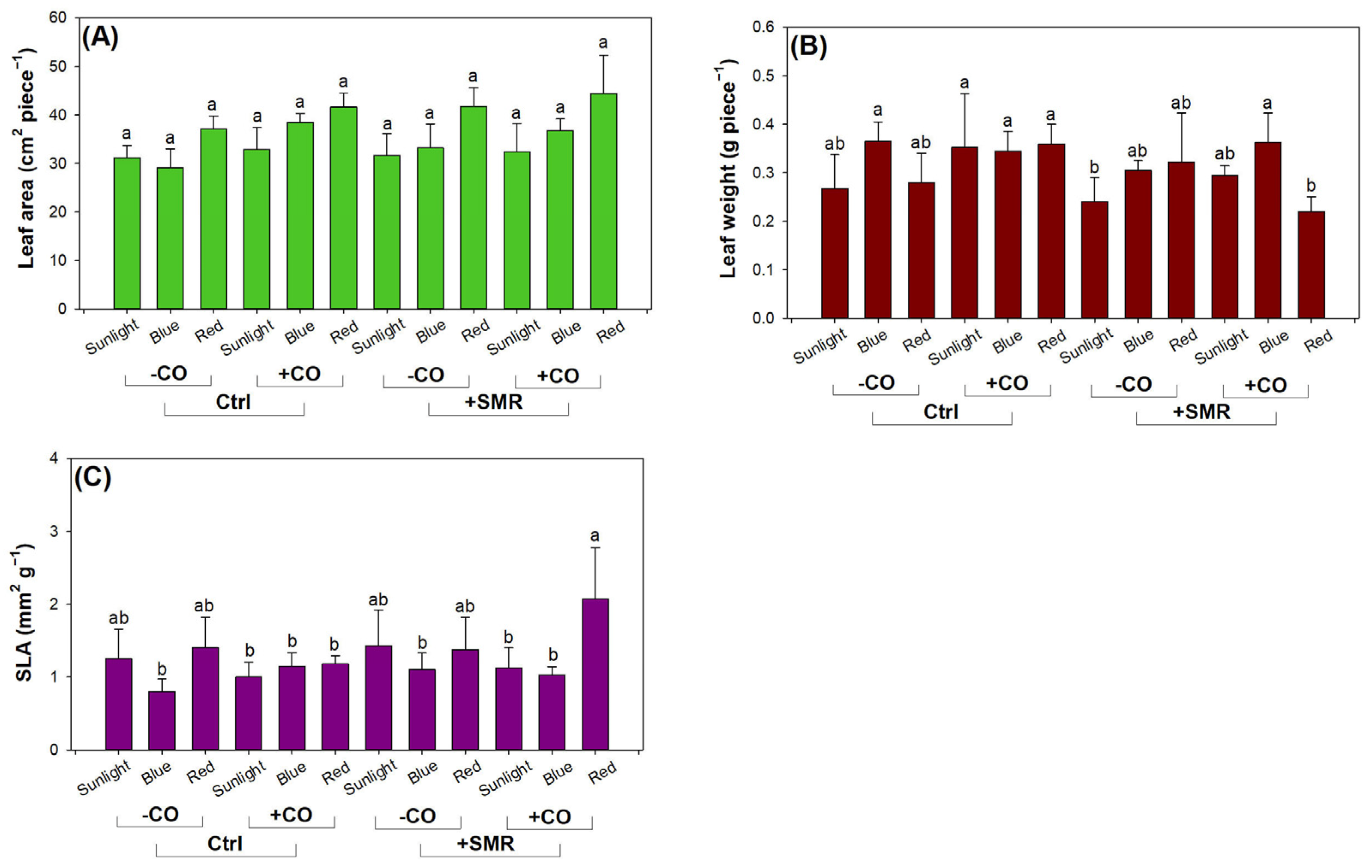
3.1. Seedling Growth and Leaf Parameter
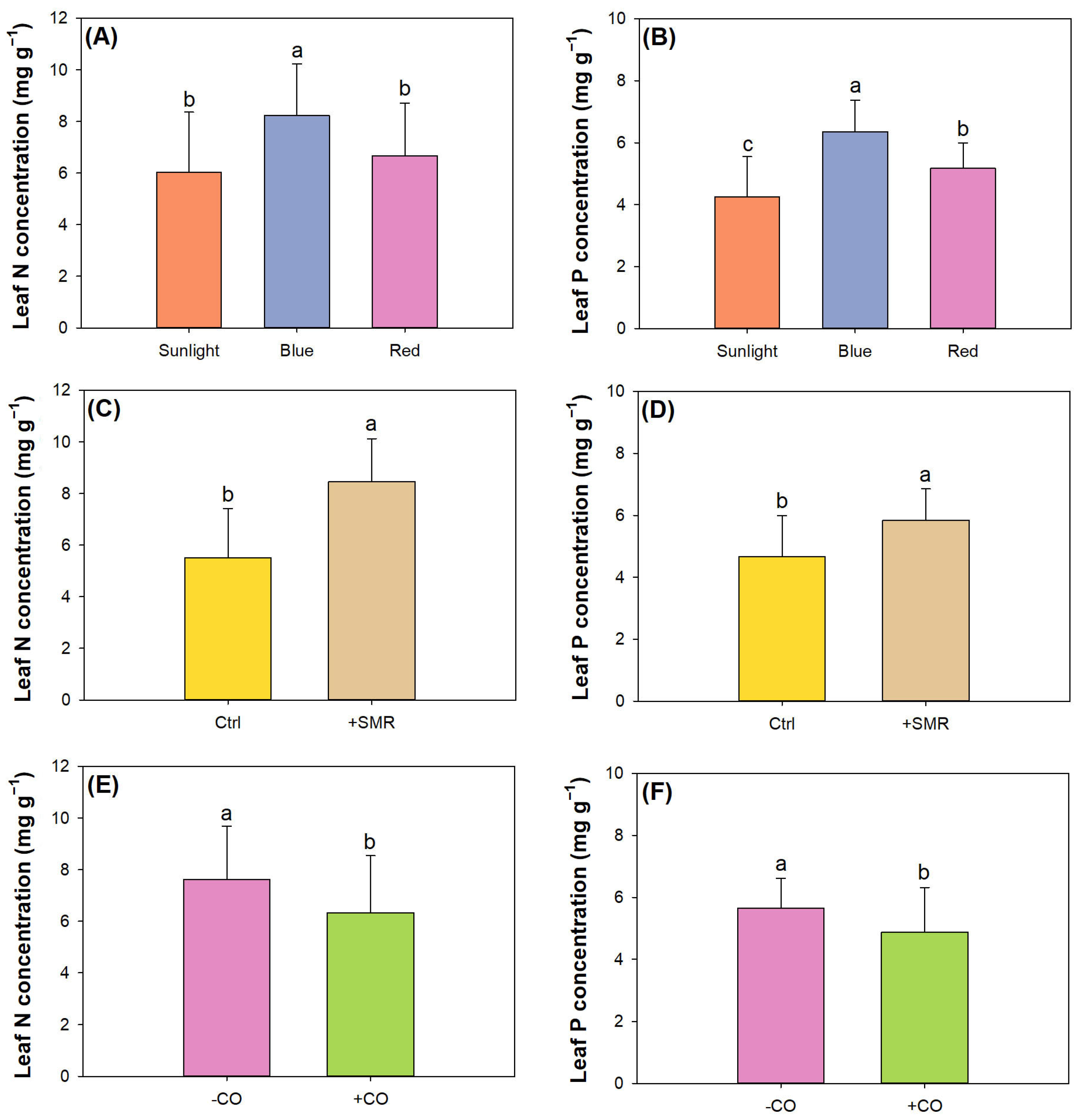
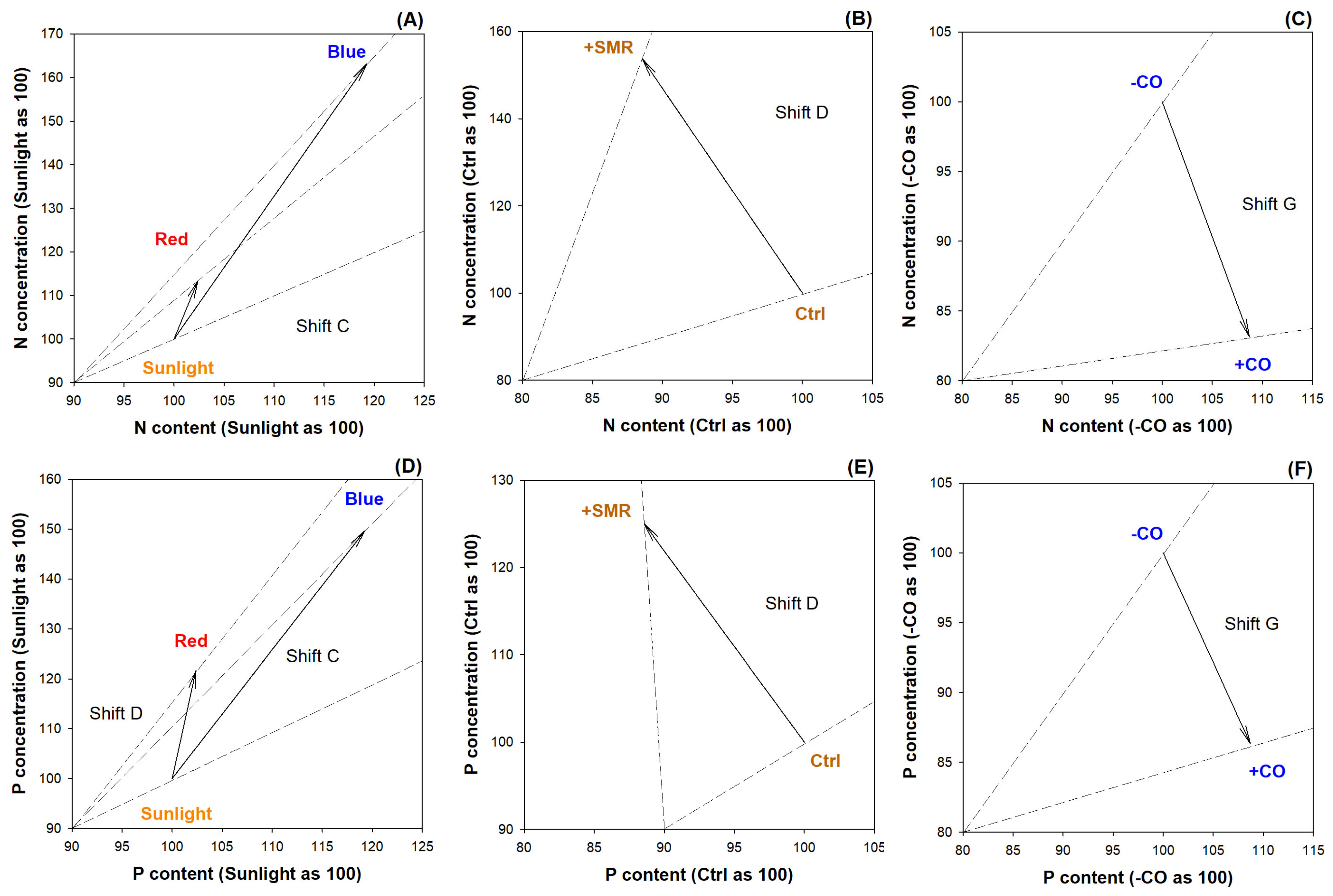
3.2. Nutrient Uptake and Status Diagnosis
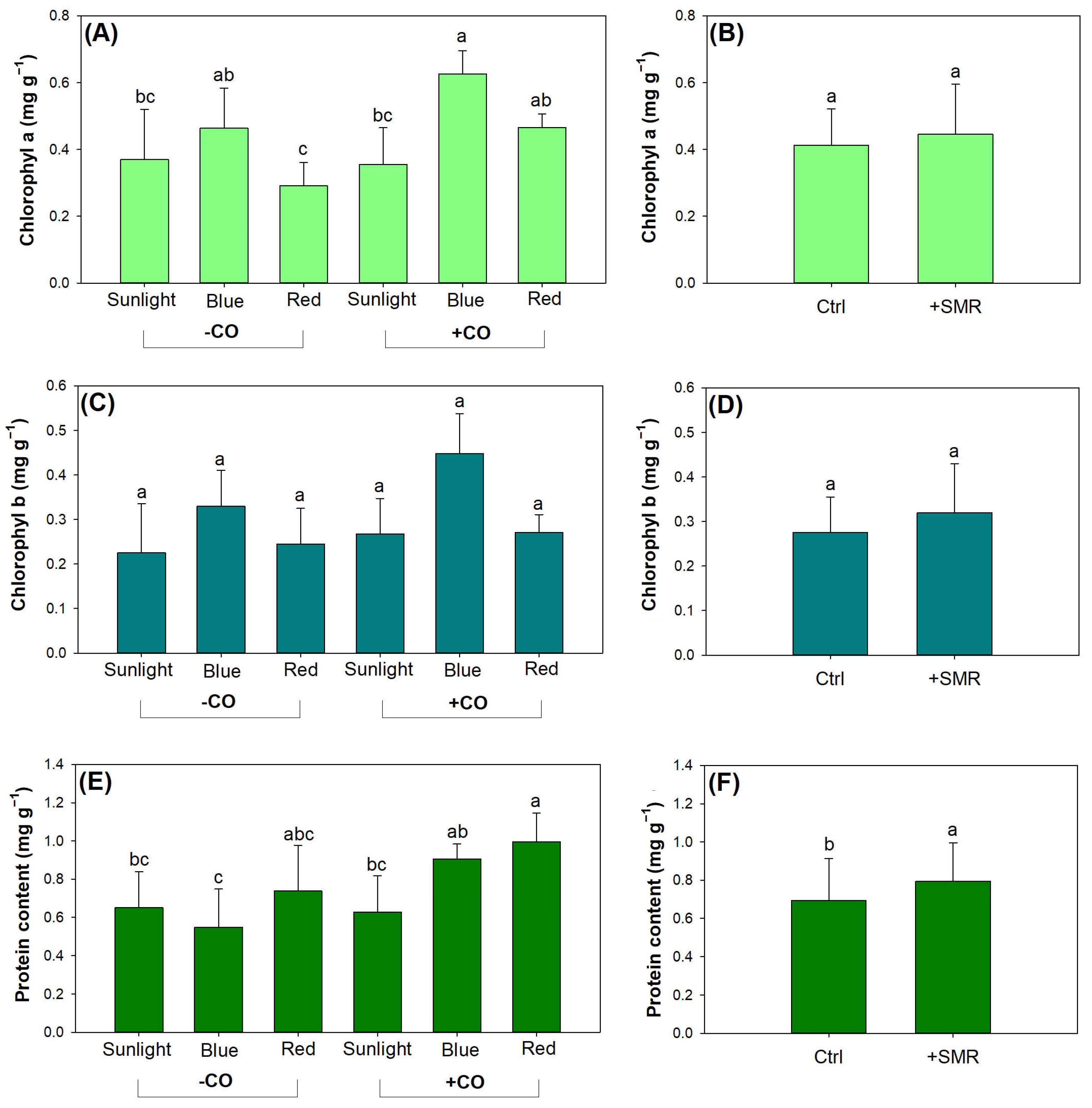
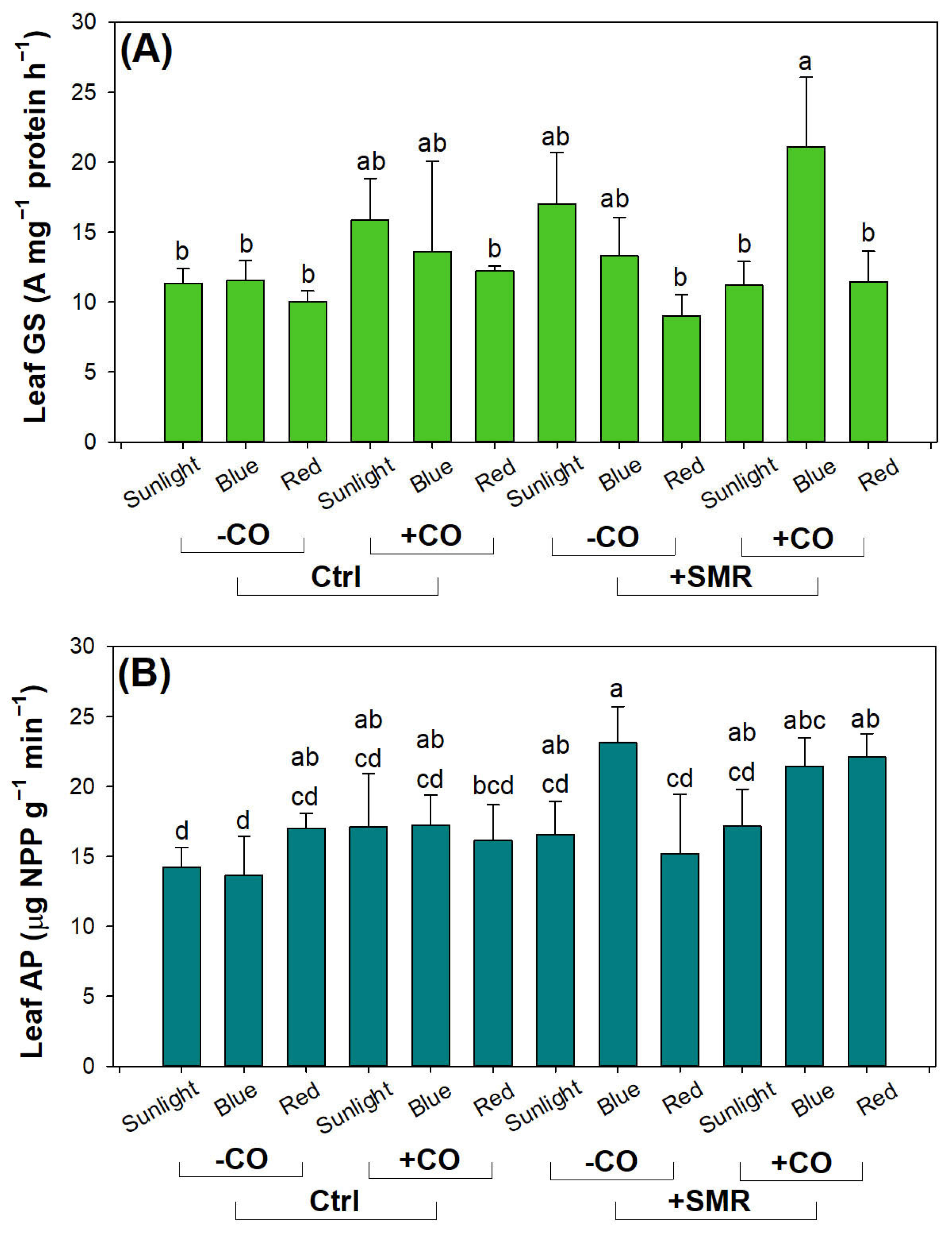
3.3. Photosynthetic Pigment and Nutrient Assimilation
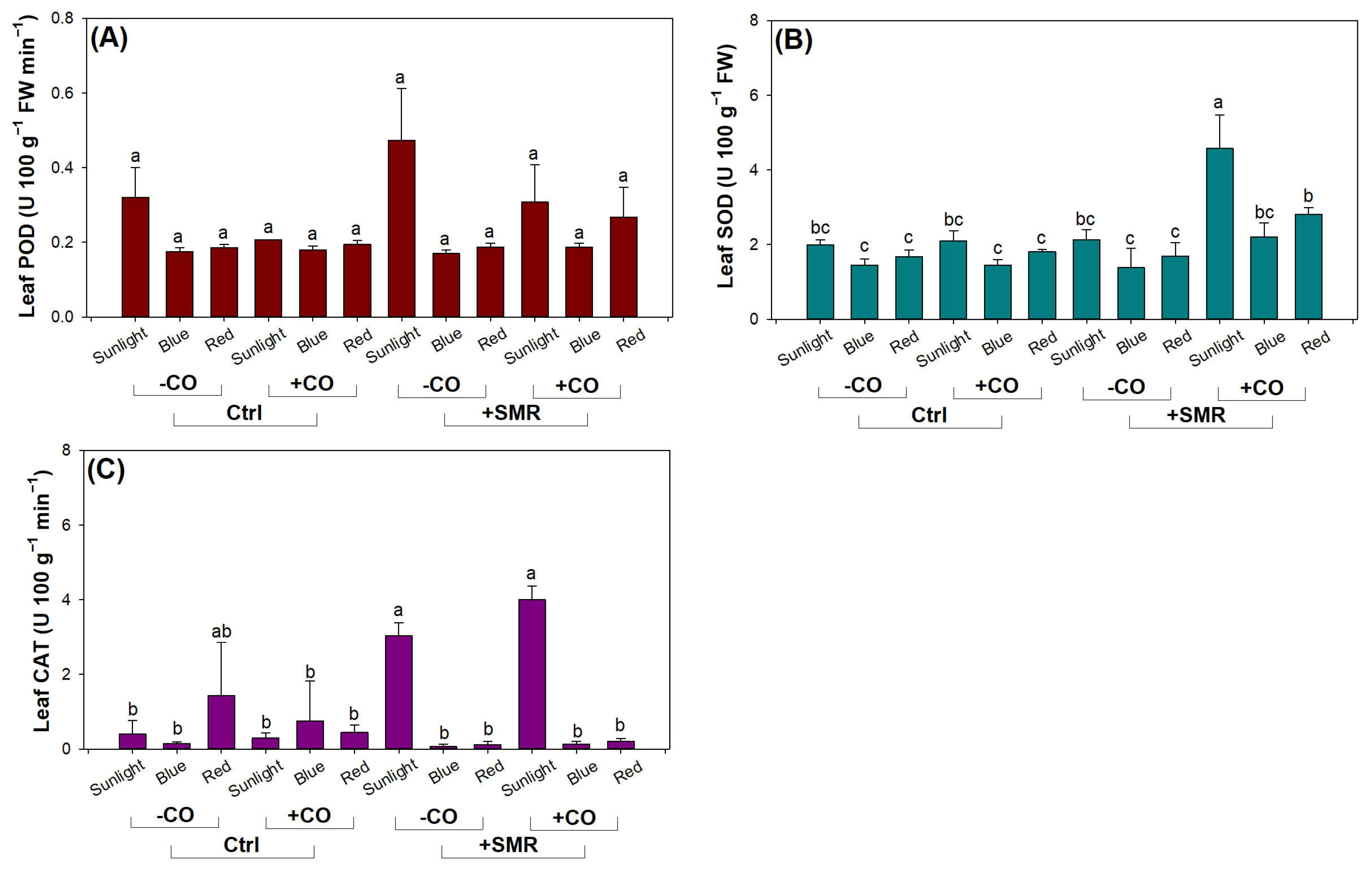
3.4. Antioxidant Activity
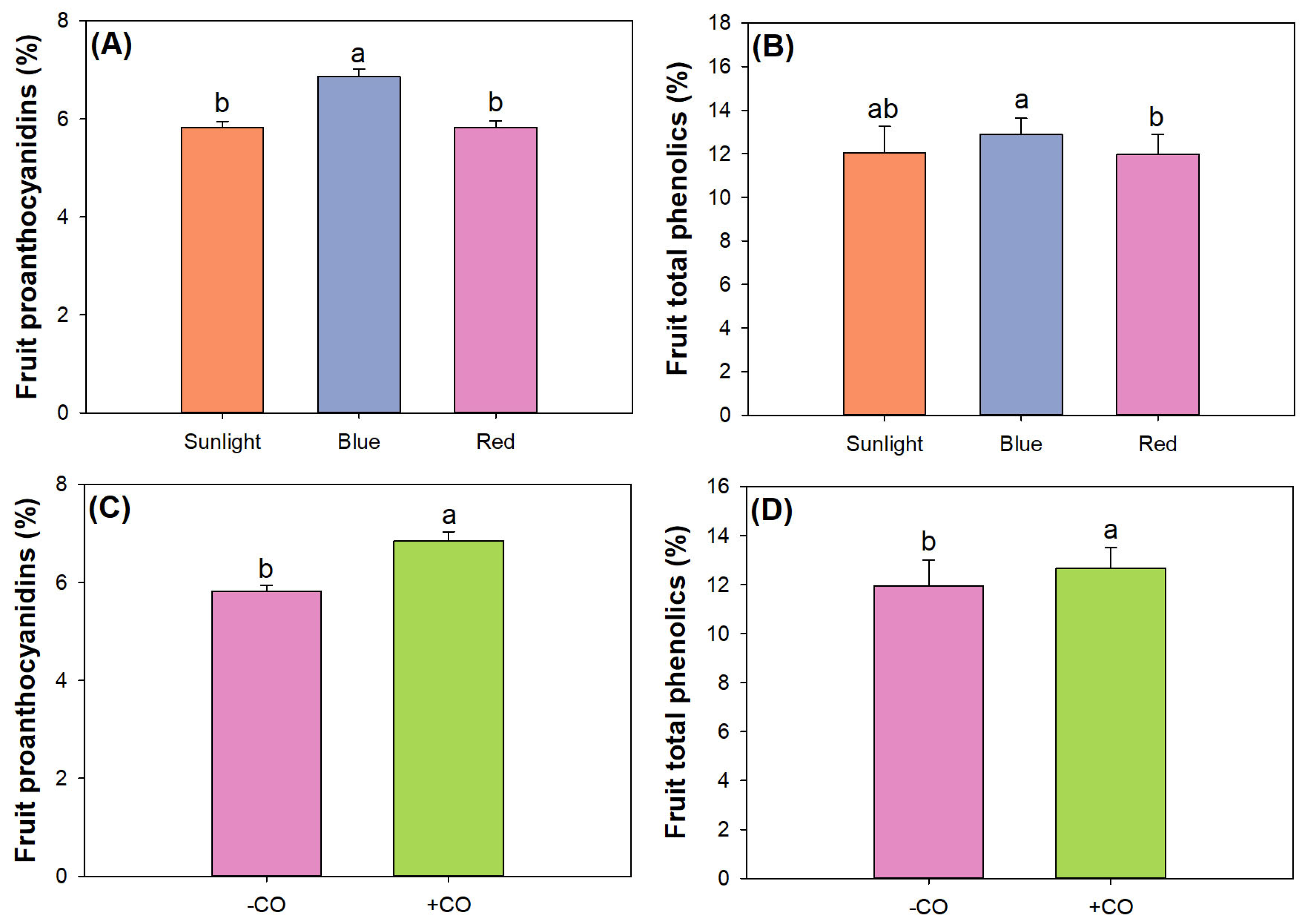
3.5. Fruit Quality Assessment
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- United Nations. Sustainable Development Goals 2015. Available online: https://www.un.org/sustainabledevelopment/sustainable-development-goals/ (accessed on 12 December 2022).
- Wei, H.X.; Zhang, J.; Xu, Z.H.; Hui, T.F.; Guo, P.; Sun, Y.X. The association between plant diversity and perceived emotions for visitors in urban forests: A pilot study across 49 parks in China. Urban For. Urban Green. 2022, 73, 127613. [Google Scholar] [CrossRef]
- Baciu, G.E.; Dobrota, C.E.; Apostol, E.N. Valuing Forest Ecosystem Services. Why Is an Integrative Approach Needed? Forests 2021, 12, 677. [Google Scholar] [CrossRef]
- Reyers, B.; Selig, E.R. Global targets that reveal the social-ecological interdependencies of sustainable development. Nat. Ecol. Evol. 2020, 4, 1011–1019. [Google Scholar] [CrossRef]
- Diaz, S.; Settele, J.; Brondizio, E.S.; Ngo, H.T.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.A.; Butchart, S.H.M.; Chan, K.M.A.; et al. Pervasive human-driven decline of life on Earth points to the need for transformative change. Science 2019, 366, eaax3100. [Google Scholar] [CrossRef] [Green Version]
- Muir, G.F.; Sorrenti, S.; Vantomme, P.; Vidale, E.; Masiero, M. Into the wild: Disentangling non-wood terms and definitions for improved forest statistics. Int. For. Rev. 2020, 22, 101–119. [Google Scholar] [CrossRef]
- Guo, S.; Zhang, D.; Wei, H.; Zhao, Y.; Cao, Y.; Yu, T.; Wang, Y.; Yan, X. Climatic Factors Shape the Spatial Distribution of Concentrations of Triterpenoids in Barks of White Birch (Betula Platyphylla Suk.) Trees in Northeast China. Forests 2017, 8, 334. [Google Scholar] [CrossRef] [Green Version]
- Li, H.G.; Wang, R.Z.; Tian, Z.W.; Xu, J.H.; Sun, W.S.; Duan, R.M.; Fu, H.; Li, Y.M.; Zhang, Y.L.; Dong, L.M. Phenotypic Variation and Diversity in Fruit, Leaf, Fatty Acid, and Their Relationships to Geoclimatic Factors in Seven Natural Populations of Malania oleifera Chun et SK Lee. Forests 2022, 13, 1733. [Google Scholar] [CrossRef]
- Wen, Y.; Su, S.C.; Zhang, H.C. Effects of Canopy Microclimate on Chinese Chestnut (Castanea mollissima Blume) Nut Yield and Quality. Forests 2020, 11, 97. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Meng, L.; Wei, H. Biomass, carbohydrate, and leakage conductance in buds of six ornamental tree species subjected to a “false spring” in Northeast China. Ann. For. Res. 2022, 65, 15–30. [Google Scholar]
- Duan, Y.; Guo, B.; Zhang, L.; Li, J.; Li, S.; Zhao, W.; Yang, G.; Zhou, S.; Zhou, C.; Song, P.; et al. Interactive climate-soil forces shape the spatial distribution of foliar N:P stoichiometry in Vaccinium uliginosum planted in agroforests of Northeast China. Front. Ecol. Evol. 2022, 10, 1065680. [Google Scholar] [CrossRef]
- Wei, H.X.; Chen, X.; Chen, G.S.; Zhao, H.T. Foliar nutrient and carbohydrate in Aralia elata can be modified by understory light quality in forests with different structures at Northeast China. Ann. For. Res. 2019, 62, 125–137. [Google Scholar] [CrossRef]
- Schaubroeck, T. Towards a general sustainability assessment of human/industrial and nature-based solutions. Sustain. Sci. 2018, 13, 1185–1191. [Google Scholar] [CrossRef]
- Duan, Y.; Fu, X.; Zhou, X.; Gao, D.; Zhang, L.; Wu, F. Removal of Dominant Species Impairs Nitrogen Utilization in Co-Existing Ledum palustre and Vaccinium uliginosum Communities Subjected to Five-Year Continuous Interruptions. Agronomy 2022, 12, 932. [Google Scholar] [CrossRef]
- Wei, H.X.; Zhao, H.T.; Chen, X. Foliar N:P Stoichiometry in Aralia elata Distributed on Different Slope Degrees. Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 887–895. [Google Scholar] [CrossRef] [Green Version]
- Chiang, C.; Aas, O.T.; Jetmundsen, M.R.; Lee, Y.; Torre, S.; Floistad, I.S.; Olsen, J.E. Day Extension with Far-Red light Enhances Growth of Subalpine Fir (Abies lasiocarpa (Hooker) Nuttall) Seedlings. Forests 2018, 9, 175. [Google Scholar] [CrossRef] [Green Version]
- Li, X.W.; Xia, H.T.; Wang, J.W.; Chen, Q.X. Nutrient uptake and assimilation in fragrant rosewood (Dalbergia odorifera TC Chen) seedlings in growing media with un-composted spent mushroom residue. PLoS ONE 2021, 16, e0249534. [Google Scholar]
- He, C.X.; Zhao, Y.; Zhang, J.S.; Gao, J. Chitosan Oligosaccharide Addition to Buddhist Pine (Podocarpus macrophyllus (Thunb) Sweet) under Drought: Reponses in Ecophysiology and delta C-13 Abundance. Forests 2020, 11, 526. [Google Scholar] [CrossRef]
- Gao, J.; Zhang, J.S.; He, C.X.; Wang, Q.R. Effects of light spectra and N-15 pulses on growth, leaf morphology, physiology, and internal nitrogen cycling in Quercus variabilis Blume seedlings. PLoS ONE 2021, 16, e0243954. [Google Scholar] [CrossRef]
- Valerio, M.; Ibanez, R.; Gazol, A. The Role of Canopy Cover Dynamics over a Decade of Changes in the Understory of an Atlantic Beech-Oak Forest. Forests 2021, 12, 938. [Google Scholar] [CrossRef]
- Tanioka, Y.; Ida, H.; Hirota, M. Relationship between Canopy Structure and Community Structure of the Understory Trees in a Beech Forest in Japan. Forests 2022, 13, 494. [Google Scholar] [CrossRef]
- Chen, C.L.; Luo, X.H.; Jin, G.R.; Cheng, Z.; Pan, X.Y.; Zhu, G.L.; Li, S.; Zhu, Y.G.; Tang, N.N. Shading effect on survival, growth, and contents of secondary metabolites in micropropagated Anoectochilus plantlets. Braz. J. Bot. 2017, 40, 599–607. [Google Scholar] [CrossRef]
- Zhou, C.; Cui, W.; Yuan, T.; Cheng, H.; Su, Q.; Guo, P. Water content, carbohydrate accumulation, and secondary metabolites in Allium victorialis sprouts exposed to shoot cutting in varied irradiations. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12524. [Google Scholar] [CrossRef]
- Zhou, C.; Shang, C.; Chen, F.; Bao, J.; Yu, L.; Guo, P. Light-emitting diode spectra modify nutritional status, physiological response, and secondary metabolites in Ficus hirta and Alpinia oxyphylla. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12314. [Google Scholar] [CrossRef]
- Guo, S.; Wang, H.; Sui, Y.; Liu, X.; Tan, L. Bioactive extracts and association with C and N in Eleutherococcus senticosus subjected to chitosan nanoparticles in contrasting light spectra. PLoS ONE 2022, 17, e0277233. [Google Scholar] [CrossRef]
- Wei, H.; Ma, B.; Hauer, R.J.; Liu, C.; Chen, X.; He, X. Relationship between environmental factors and facial expressions of visitors during the urban forest experience. Urban For. Urban Green. 2020, 53, 126699. [Google Scholar] [CrossRef]
- Velasco, M.H.; Mattsson, A. Light Shock Stress after Outdoor Sunlight Exposure in Seedlings of Picea abies (L.) Karst. and Pinus sylvestris L. Pre-Cultivated under LEDs-Possible Mitigation Treatments and Their Energy Consumption. Forests 2020, 11, 354. [Google Scholar] [CrossRef] [Green Version]
- Wei, H.X.; Hauer, R.J.; Chen, G.S.; Chen, X.; He, X.Y. Growth, Nutrient Assimilation, and Carbohydrate Metabolism in Korean Pine (Pinus koraiensis) Seedlings in Response to Light Spectra. Forests 2020, 11, 44. [Google Scholar] [CrossRef] [Green Version]
- Song, Q.Y.; Xu, L.; Long, W.; Guo, J.; Zhang, X. Quality assessment and nutrient uptake and utilization in Luohan pine (Podocarpus macrophyllus) seedlings raised by chitosan spraying in varied LED spectra. PLoS ONE 2022, 17, e0267632. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, C.; Wu, Y. Dry mass input into fruits can be predicted by fine root morphology of pepper cultivars exposed to varied lighting spectra. Not. Bot. Horti Agrobot. Cluj-Napoca 2022, 50, 12930. [Google Scholar] [CrossRef]
- Wei, H.X.; Zhao, H.T.; Chen, X.; He, X.Y. Secondary metabolites, carbohydrate accumulation, and nutrient uptake in Aralia elata (Miq.) Seem seedlings exposed to shoot cutting and different LED spectra. Acta Physiol. Plant. 2020, 42, 162. [Google Scholar] [CrossRef]
- Choi, J.; Kim, J.; Yoon, H.I.; Son, J.E. Effect of far-red and UV-B light on the growth and ginsenoside content of ginseng (Panax ginseng C. A. Meyer) sprouts aeroponically grown in plant factories. Hortic. Environ. Biotechnol. 2022, 63, 77–87. [Google Scholar] [CrossRef]
- Zhou, C.W.; Cui, W.J.; Yuan, T.; Cheng, H.Y.; Su, Q.; Wei, H.X.; Guo, P. Root Foraging Behavior of Two Agronomical Herbs Subjected to Heterogeneous P Pattern and High Ca Stress. Agronomy 2022, 12, 624. [Google Scholar] [CrossRef]
- He, C.X.; Gao, J.; Zhao, Y.; Liu, J. Root Foraging Precision of Pinus pumila (Pall.) Regel Subjected to Contrasting Light Spectra. Plants 2021, 10, 1482. [Google Scholar] [CrossRef] [PubMed]
- Ramsey, C.L.; Jose, S.; Brecke, B.J.; Merritt, S. Growth response of longleaf pine (Pinus palustris Mill.) seedlings to fertilization and herbaceous weed control in an old field in southern USA. For. Ecol. Manag. 2003, 172, 281–289. [Google Scholar] [CrossRef]
- Qasem, J.R. Response of onion (Allium cepa L.) plants to fertilizers, weed competition duration, and planting times in the central Jordan Valley. Weed Biol. Manag. 2006, 6, 212–220. [Google Scholar] [CrossRef]
- Gallina, G.C.; Cregg, B.M.; Patterson, E.L.; Saha, D. A Review of Chemical Weed Control Practices in Christmas Tree Production in the United States. Forests 2022, 13, 250. [Google Scholar] [CrossRef]
- Madrid-Aispuro, R.E.; Prieto-Ruiz, J.A.; Aldrete, A.; Hernandez-Diaz, J.C.; Wehenkel, C.; Chavez-Simental, J.A.; Mexal, J.G. Alternative Substrates and Fertilization Doses in the Production of Pinus cembroides Zucc. in Nursery. Forests 2020, 11, 71. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.; Sun, X.K.; Xu, C.B.; Ma, X.P.; Huang, Y.; Fan, Z.P.; Cao, X.Y. Effects of Sewage Sludge Application on Plant Growth and Soil Characteristics at a Pinus sylvestris var. mongolica Plantation in Horqin Sandy Land. Forests 2022, 13, 984. [Google Scholar] [CrossRef]
- Wei, H.X.; Guo, P. Carbohydrate metabolism during new root growth in transplanted Larix olgensis seedlings: Post-transplant response to nursery-applied inorganic fertilizer and organic amendment. iFor. Biogeosci. For. 2017, 10, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Zhao, S.; Jin, A.; Tang, J.; Luo, Y. The use of un-composted spent mushroom residue as a replacement of peat in substrates for Gossypium herbaceum and Talinum paniculatum. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12193. [Google Scholar] [CrossRef]
- Zhu, H.; Zhao, S.; Yang, J.; Meng, L.; Luo, Y.; Hong, B.; Cui, W.; Wang, M.; Liu, W. Growth, Nutrient Uptake, and Foliar Gas Exchange in Pepper Cultured with Un-composted Fresh Spent Mushroom Residue. Not. Bot. Horti Agrobot. Cluj-Napoca 2019, 47, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Chen, X.; Wei, H.X.; Lv, J.; Chen, C.; Liu, X.Y.; Wen, Q.; Jia, L.M. Nutrient uptake and utilization in Prince Rupprecht’s larch (Larix principis-rupprechtii Mayr.) seedlings exposed to a combination of light-emitting diode spectra and exponential fertilization. Soil Sci. Plant Nutr. 2019, 65, 358–368. [Google Scholar] [CrossRef]
- Chu, X.L.; Luo, X.Y.; Zhou, Z.C. Exponential fertilization on red-seed tree (Ormosia hosiei) seedlings subjected to contrasting light conditions: Do we really need intensive nutrient loading? Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12244. [Google Scholar] [CrossRef]
- Tan, L.; Fan, R.; Sun, H.; Guo, S. Root foraging of birch and larch in heterogeneous soil nutrient patches under water deficit. PLoS ONE 2021, 16, e0255848. [Google Scholar] [CrossRef]
- Aung, T.; Lee, W.H.; Eun, J.B. Metabolite profiling and pathway prediction of laver (Porphyra dentata) kombucha during fermentation at different temperatures. Food Chem. 2022, 397, 133636. [Google Scholar] [CrossRef]
- Sharma, D.; Shree, B.; Kumar, S.; Kumar, V.; Sharma, S.; Sharma, S. Stress induced production of plant secondary metabolites in vegetables: Functional approach for designing next generation super foods. Plant Physiol. Biochem. 2022, 192, 252–272. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Ding, J.R.; Huang, W.J.; Yu, H.; Wu, S.W.; Li, W.Y.; Mao, X.X.; Chen, W.F.; Xing, J.L.; Li, C.; et al. OsPP65 Negatively Regulates Osmotic and Salt Stress Responses Through Regulating Phytohormone and Raffinose Family Oligosaccharide Metabolic Pathways in Rice. Rice 2022, 15, 34. [Google Scholar] [CrossRef]
- Lyu, R.H.; Wang, R.; Wu, C.Y.; Bao, Y.J.; Guo, P. Comparative transcriptome analysis of leaves of sour jujube seedlings under salt stress. Acta Physiol. Plant. 2022, 44, 119. [Google Scholar] [CrossRef]
- Liu, G.D.; Jin, T.; Xu, Y.M.; Yao, F.X.; Guan, G.; Zhou, G.F. Exogenous citrate restores the leaf metabolic profiles of navel orange plants under boron deficiency. Plant Physiol. Biochem. 2022, 192, 101–109. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, Y.; Wei, H.X. Chitosan oligosaccharide addition affects current-year shoot of post-transplant Buddhist pine (Podocarpus macrophyllus) seedlings under contrasting photoperiods. iFor. Biogeosci. For. 2017, 10, 715–721. [Google Scholar] [CrossRef] [Green Version]
- He, C.; Zhao, Y.; Wang, Y.; Cai, J.; Gao, J.; Zhang, J. Forage quality and physiological performance of mowed alfalfa (Medicago sativa L.) subjected to combined light quality and drought. Front. Plant Sci. 2022, 13, 1047294. [Google Scholar] [CrossRef] [PubMed]
- Li, X.W.; Chen, Q.X.; Lei, H.Q.; Wang, J.W.; Yang, S.; Wei, H.X. Nutrient Uptake and Utilization by Fragrant Rosewood (Dalbergia odorifera) Seedlings Cultured with Oligosaccharide Addition under Different Lighting Spectra. Forests 2018, 9, 29. [Google Scholar] [CrossRef]
- Guo, S.L.; Zhang, S.; Jia, L.W.; Xu, M.Y.; Wang, Z.Y. Root growth of Eleuthero (Eleutherococcus senticosus Rupr. & Maxim. Maxim.) seedlings cultured with chitosan oligosaccharide addition under different light spectra. Not. Bot. Horti Agrobot. Cluj-Napoca 2020, 48, 626–635. [Google Scholar]
- Pankhurst, R.J. Aronia melanocarpa. In Flora of North America North of Mexico (FNA); Flora of North America Editorial Committee, Ed.; Oxford University Press: Oxford, UK; New York, NY, USA, 2014; Volume 9. [Google Scholar]
- Ochmian, I.; Grajkowski, J.; Smolik, M. Comparison of Some Morphological Features, Quality and Chemical Content of Four Cultivars of Chokeberry Fruits (Aronia melanocarpa). Not. Bot. Horti Agrobot. Cluj-Napoca 2012, 40, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Persson, H.A.; Jeppsson, N.; Nybom, H. Genetic variation in wild and cultivated material of black chokeberry. In Proceedings of the International Symposium on Molecular Markers for Characterizing Genotypes and Identifying Cultivars in Horticulture, Montpellier, France, 6–8 March 2000; pp. 253–255. [Google Scholar]
- Zelazinski, T. Properties of Biocomposites from Rapeseed Meal, Fruit Pomace and Microcrystalline Cellulose Made by Press Pressing: Mechanical and Physicochemical Characteristics. Materials 2021, 14, 890. [Google Scholar] [CrossRef] [PubMed]
- Sosnowska, D.; Kajszczak, D.; Podsedek, A. The Effect of Different Growth Stages of Black Chokeberry Fruits on Phytonutrients, Anti-Lipase Activity, and Antioxidant Capacity. Molecules 2022, 27, 8031. [Google Scholar] [CrossRef]
- Kobus, Z.; Nadulski, R.; Wilczyński, K.; Kozak, M.; Guz, T.; Rydzak, L. Effect of the black chokeberry (Aronia melanocarpa (Michx.) Elliott) juice acquisition method on the content of polyphenols and antioxidant activity. PLoS ONE 2019, 14, e0219585. [Google Scholar] [CrossRef] [Green Version]
- Bushmeleva, K.; Vyshtakalyuk, A.; Terenzhev, D.; Belov, T.; Nikitin, E.; Zobov, V. Antioxidative and Immunomodulating Properties of Aronia melanocarpa Extract Rich in Anthocyanins. Plants 2022, 11, 3333. [Google Scholar] [CrossRef]
- Velmurugan, P.; Vedhanayakisri, K.A.; Park, Y.J.; Jin, J.S.; Oh, B.T. Use of Aronia melanocarpa Fruit Dye Combined with Silver Nanoparticles to Dye Fabrics and Leather and Assessment of Its Antibacterial Potential Against Skin Bacteria. Fibers Polym. 2019, 20, 302–311. [Google Scholar] [CrossRef]
- Monteleone, A.; Skousen, J.; Shuler, J.; McDonald, L.; Williams, R.; Holaskova, I. Survival and growth of 20 species of trees and shrubs on Appalachian surface mines. Land Degrad. Dev. 2018, 29, 1683–1693. [Google Scholar] [CrossRef] [Green Version]
- Bussieres, J.; Boudreau, S.; Clement-Mathieu, G.; Dansereau, B.; Rochefort, L. Growing black chokeberry (Aronia melanocarpa) in cut-over peatlands. Hortscience 2008, 43, 494–499. [Google Scholar] [CrossRef] [Green Version]
- An, B.Y.; Wei, H.X.; Li, L.L.; Guo, P. Nutrient Uptake and Utilization and Antioxidants of Fruits in Red Raspberry (Rubus idaeus L.) Cultivar ‘Autumn Bliss’ in response to Fertilization under Extended Photoperiod. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 46, 440–448. [Google Scholar] [CrossRef] [Green Version]
- Carew, J.G.; Mahmood, K.; Darby, J.; Hadley, P.; Battey, N.H. The effects of low temperatures on the vegetative growth and flowering of the primocane fruiting raspberry ‘Autumn Bliss’. J. Hortic. Sci. Biotechnol. 2001, 76, 264–270. [Google Scholar] [CrossRef]
- Sonsteby, A.; Heide, O.M. Effects of photoperiod and temperature on growth and flowering in the annual (primocane) fruiting raspberry (Rubus idaeus L.) cultivar ‘Polka’. J. Hortic. Sci. Biotechnol. 2009, 84, 439–446. [Google Scholar] [CrossRef]
- Roberts, J.; Hopkins, R.; Morecroft, M. Towards a predictive description of forest canopies from litter properties. Funct. Ecol. 1999, 13, 265–272. [Google Scholar] [CrossRef]
- Nosheen, A.; Yasmin, H.; Naz, R.; Bano, A.; Keyani, R.; Hussain, I. Pseudomonas putida improved soil enzyme activity and growth of kasumbha under low input of mineral fertilizers. Soil Sci. Plant Nutr. 2018, 64, 520–525. [Google Scholar] [CrossRef]
- Serapiglia, M.J.; Minocha, R.; Minocha, S.C. Changes in polyamines, inorganic ions and glutamine synthetase activity in response to nitrogen availability and form in red spruce (Picea rubens). Tree Physiol. 2008, 28, 1793–1803. [Google Scholar] [CrossRef] [Green Version]
- Kolari, K.K.; Sarjala, T. Acid-phosphatase-activity and phosphorus-nutrition in Scots pine needles. Tree Physiol. 1995, 15, 747–752. [Google Scholar] [CrossRef]
- Hussain, S.; Khalid, M.F.; Saqib, M.; Ahmad, S.; Zafar, W.; Rao, M.J.; Morillon, R.; Anjum, M.A. Drought tolerance in citrus rootstocks is associated with better antioxidant defense mechanism. Acta Physiol. Plant. 2018, 40, 135. [Google Scholar] [CrossRef]
- Podsędek, A.; Majewska, I.; Redzynia, M.; Sosnowska, D.; Koziołkiewicz, M. In Vitro Inhibitory Effect on Digestive Enzymes and Antioxidant Potential of Commonly Consumed Fruits. J. Agric. Food Chem. 2014, 62, 4610–4617. [Google Scholar] [CrossRef]
- Rösch, D.; Bergmann, M.; Knorr, D.; Kroh, L.W. Structure−Antioxidant Efficiency Relationships of Phenolic Compounds and Their Contribution to the Antioxidant Activity of Sea Buckthorn Juice. J. Agric. Food Chem. 2003, 51, 4233–4239. [Google Scholar] [CrossRef] [PubMed]
- Salifu, K.F.; Timmer, V.R. Optimizing nitrogen loading of Picea mariana seedlings during nursery culture. Can. J. For. Res. 2003, 33, 1287–1294. [Google Scholar] [CrossRef]
- Wei, H.X.; Ren, J.; Zhou, J.H. Effect of exponential fertilization on growth and nutritional status in Buddhist pine (Podocarpus macrophyllus Thunb. D. Don) seedlings cultured in natural and prolonged photoperiods. Soil Sci. Plant Nutr. 2013, 59, 933–941. [Google Scholar] [CrossRef]
- Zhang, Y.T.; Dong, H.; Song, S.W.; Su, W.; Liu, H.C. Morphological and Physiological Responses of Cucumber Seedlings to Supplemental LED Light under Extremely Low Irradiance. Agronomy 2020, 10, 1698. [Google Scholar] [CrossRef]
- Nguyen, A.D.; Vo, T.P.K.; Tran, T.D. Research on impact of chitosan oligomers on biophysical characteristics, growth, development and drought resistance of coffee. Carbohydr. Polym. 2011, 84, 751–755. [Google Scholar]
- Elmlinger, M.W.; Bolle, C.; Batschauer, A.; Oelmüller, R.; Mohr, H. Coaction of blue light and light absorbed by phytochrome in control of glutamine synthetase gene expression in Scots pine (Pinus sylvestris L.) seedlings. Planta 1994, 192, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Ru, L.; Jiang, L.F.; Wills, R.B.H.; Golding, J.B.; Huo, Y.R.; Yang, H.Q.; Li, Y.X. Chitosan oligosaccharides induced chilling resistance in cucumber fruit and associated stimulation of antioxidant and HSP gene expression. Sci. Hortic. 2020, 264, 109187. [Google Scholar] [CrossRef]
- Zhang, Y.T.; Hu, W.J.; Peng, X.R.; Sun, B.; Wang, X.R.; Tang, H.R. Characterization of anthocyanin and proanthocyanidin biosynthesis in two strawberry genotypes during fruit development in response to different light qualities. J. Photochem. Photobiol. B Biol. 2018, 186, 225–231. [Google Scholar] [CrossRef]
- Acemi, A.; Gun Polat, E.; Cakir, M.; Demiryurek, E.; Yavuz, B.; Ozen, F. Molecular weight and concentration of chitosan affect plant development and phenolic substance pattern in arugula. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 12296. [Google Scholar] [CrossRef]
- Jing, Y.J.; Huang, J.H.; Yu, X.Q. Preparation, characterization, and functional evaluation of proanthocyanidin-chitosan conjugate. Carbohydr. Polym. 2018, 194, 139–145. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light Type | Current Composition 1 | PPFD 2 (µ mol−1 m−2 s−1) | Spectrum Composition 3 |
|---|---|---|---|
| Red-tinted | 70% R and 10% B + G | 72.39 ± 1.99 | 69.4%:30.2%:0.4% 4 |
| Blue-tinted | 30% R and 100% B + G | 70.81 ± 2.22 | 15.3%:64.9%:19.8% |
| Source | NH4+-N 1 (mg kg−1) | NO3−-N 2 (mg kg−1) | PO43−-P 3 (mg kg−1) | pH | EC 4 (dS m−1) |
|---|---|---|---|---|---|
| SMR | 11.59 ± 1.21 a 5 | 12.47 ± 2.15 b | 773.57 ± 159.50 a | 5.36 ± 0.29 b | 2.30 ± 0.34 a |
| Soil | 3.97 ± 3.05 b | 93.34 ± 9.39 a | 1.85 ± 0.29 b | 6.19 ± 0.56 a | 0.13 ± 0.04 b |
| F value | 18.73 | 245.26 | 81.52 | 6.14 | 137.41 |
| p value | 0.0049 | <0.0001 | 0.0001 | 0.0479 | <0.0001 |
| Source | Height | RCD 1 | LA 2 | LW 3 | SLA 4 |
|---|---|---|---|---|---|
| Light | 16.18 *** 5 | 3.88 * | 17.07 *** | 3.55 * | 6.96 ** |
| Soil | 13.96 *** | 0.07 | 1.49 | 4.07 | 4.88 * |
| CO | 13.57 *** | 3.74 | 8.01 ** | 1.93 | 0.03 |
| Light × Soil | 0.03 | 0.15 | 0.67 | 0.20 | 1.16 |
| Light × CO | 1.34 | 0.07 | 1.29 | 1.63 | 2.03 |
| Soil × CO | 1.12 | 4.12 | 1.18 | 1.46 | 0.58 |
| Light × Soil × CO | 0.14 | 1.81 | 0.30 | 4.12 * | 3.28 * |
| Source | TN 1 | TP 2 | Chla 3 | Chlb 4 | Protein 5 | GS 6 | AP 7 |
|---|---|---|---|---|---|---|---|
| Light | 6.40 ** 8 | 20.10 *** | 12.21 *** | 12.78 *** | 8.87 *** | 7.28 ** | 3.47 * |
| Soil | 32.32 *** | 18.40 *** | 1.00 | 3.05 | 5.28 * | 2.25 | 17.56 *** |
| CO | 6.12 * | 8.01 ** | 10.37 ** | 5.91 * | 19.43 *** | 5.51 * | 5.53 * |
| Light × Soil | 0.23 | 1.03 | 0.79 | 0.96 | 6.26 ** | 3.14 | 4.75 * |
| Light × CO | 1.80 | 5.07 * | 3.38 * | 1.21 | 6.51 ** | 2.93 | 0.55 |
| Soil × CO | 0.07 | 0.34 | 0.36 | 1.20 | 16.43 *** | 0.59 | 0.01 |
| Light × Soil × CO | 2.20 | 0.48 | 0.22 | 0.67 | 0.62 | 6.28 ** | 6.04 ** |
| Source | POD 1 | SOD 2 | CAT 3 | Proa 4 | Total Phenolics |
|---|---|---|---|---|---|
| Light | 23.36 *** 5 | 30.22 *** | 14.30 *** | 13.45 *** | 3.65 * |
| Soil | 8.59 ** | 39.29 *** | 65.12 *** | 0.39 | 2.07 |
| CO | 2.15 | 45.64 *** | 1.83 | 25.36 *** | 5.53 * |
| Light × Soil | 3.91 * | 6.68 *** | 9.42 *** | 0.23 | 1.35 |
| Light × CO | 9.05 *** | 5.02 * | 23.90 *** | 0.33 | 0.13 |
| Soil × CO | 0.07 | 36.87 *** | 2.40 | 0.72 | 0.50 |
| Light × Soil × CO | 0.89 | 4.47 * | 21.49 *** | 0.36 | 0.59 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, Y.; Wei, X.; Zhao, W.; Li, J.; Yang, G.; Zhou, S.; Zhou, C.; Zhang, L.; Li, P.; Hou, S.; et al. Natural Bioactive Substances in Fruits of Aronia melanocarpa (Michx.) Elliott Exposed to Combined Light-Type, Chitosan Oligosaccharide, and Spent Mushroom Residue Treatments. Plants 2023, 12, 604. https://doi.org/10.3390/plants12030604
Duan Y, Wei X, Zhao W, Li J, Yang G, Zhou S, Zhou C, Zhang L, Li P, Hou S, et al. Natural Bioactive Substances in Fruits of Aronia melanocarpa (Michx.) Elliott Exposed to Combined Light-Type, Chitosan Oligosaccharide, and Spent Mushroom Residue Treatments. Plants. 2023; 12(3):604. https://doi.org/10.3390/plants12030604
Chicago/Turabian StyleDuan, Yadong, Xin Wei, Wenbo Zhao, Jinxia Li, Guang Yang, Shuang Zhou, Chunwei Zhou, Lei Zhang, Pengju Li, Shuai Hou, and et al. 2023. "Natural Bioactive Substances in Fruits of Aronia melanocarpa (Michx.) Elliott Exposed to Combined Light-Type, Chitosan Oligosaccharide, and Spent Mushroom Residue Treatments" Plants 12, no. 3: 604. https://doi.org/10.3390/plants12030604