

(1′S)-1′-Acetoxyeugenol Acetate Enhances Glucose-Stimulated Insulin Secretion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
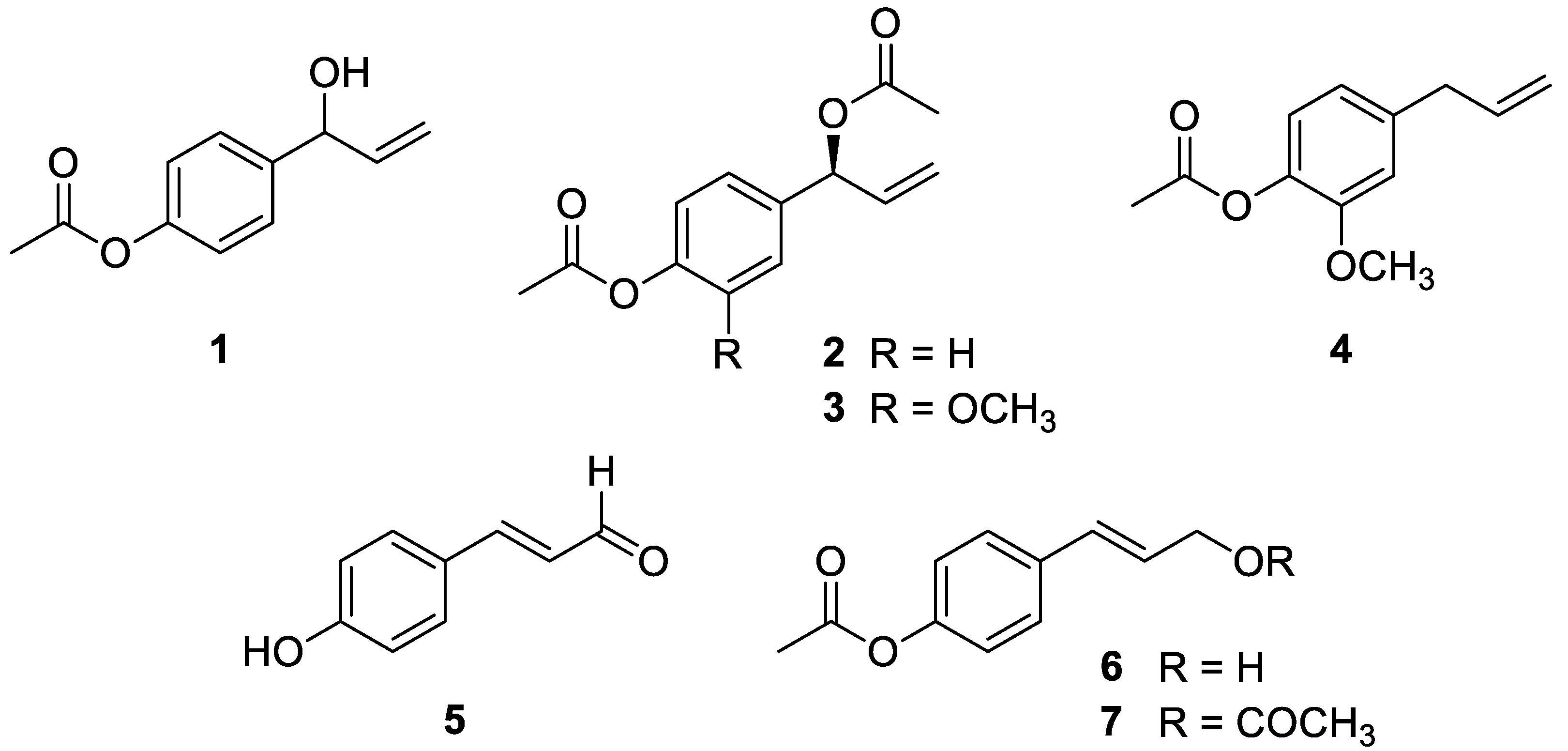
2.1. Isolation and Identification of Phenylpropanoids 1–7 from the Rhizomes of A. galanga
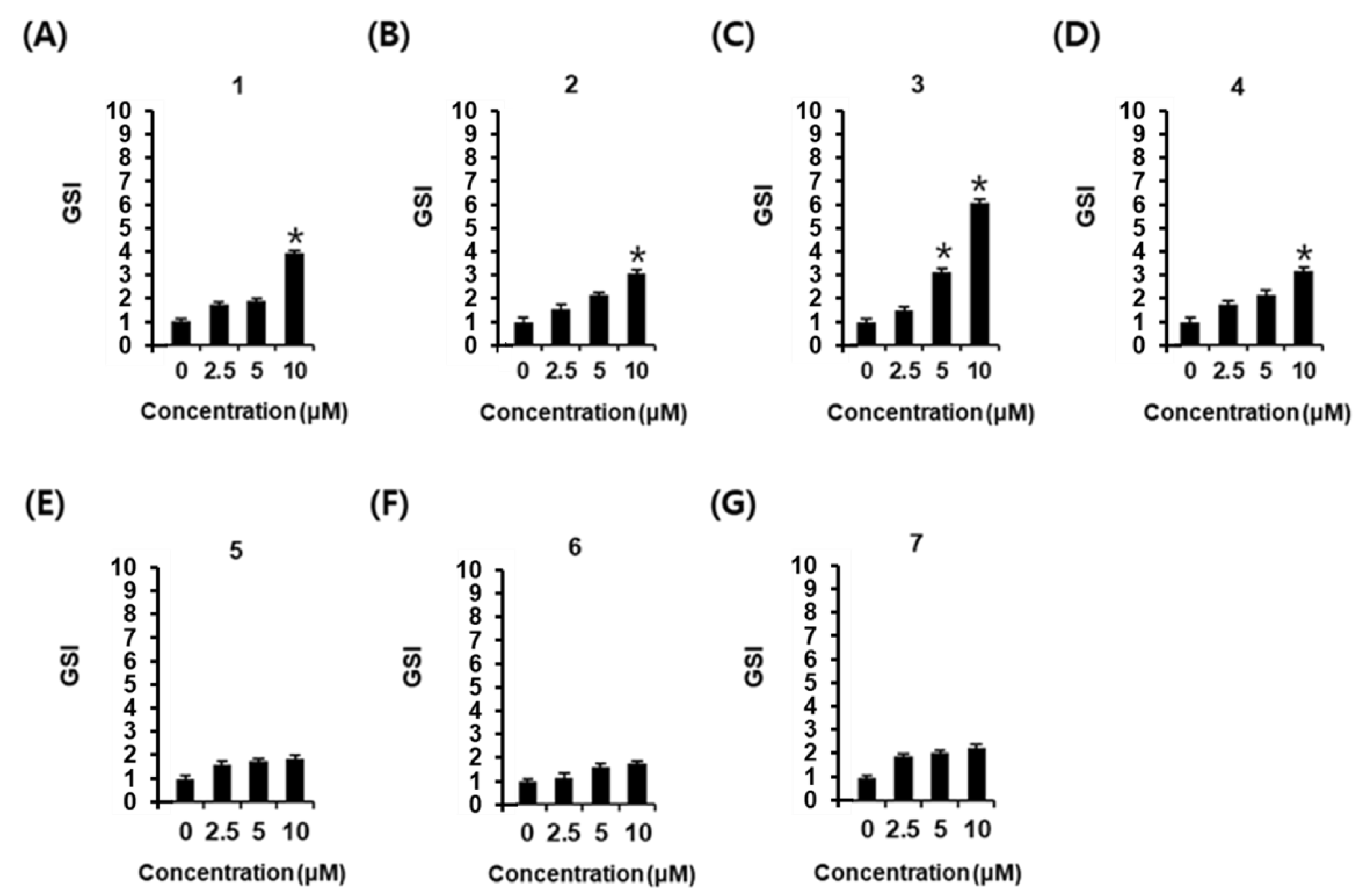
2.2. Effects of Phenylpropanoids 1–7 on Glucose-Stimulated Insulin Secretion
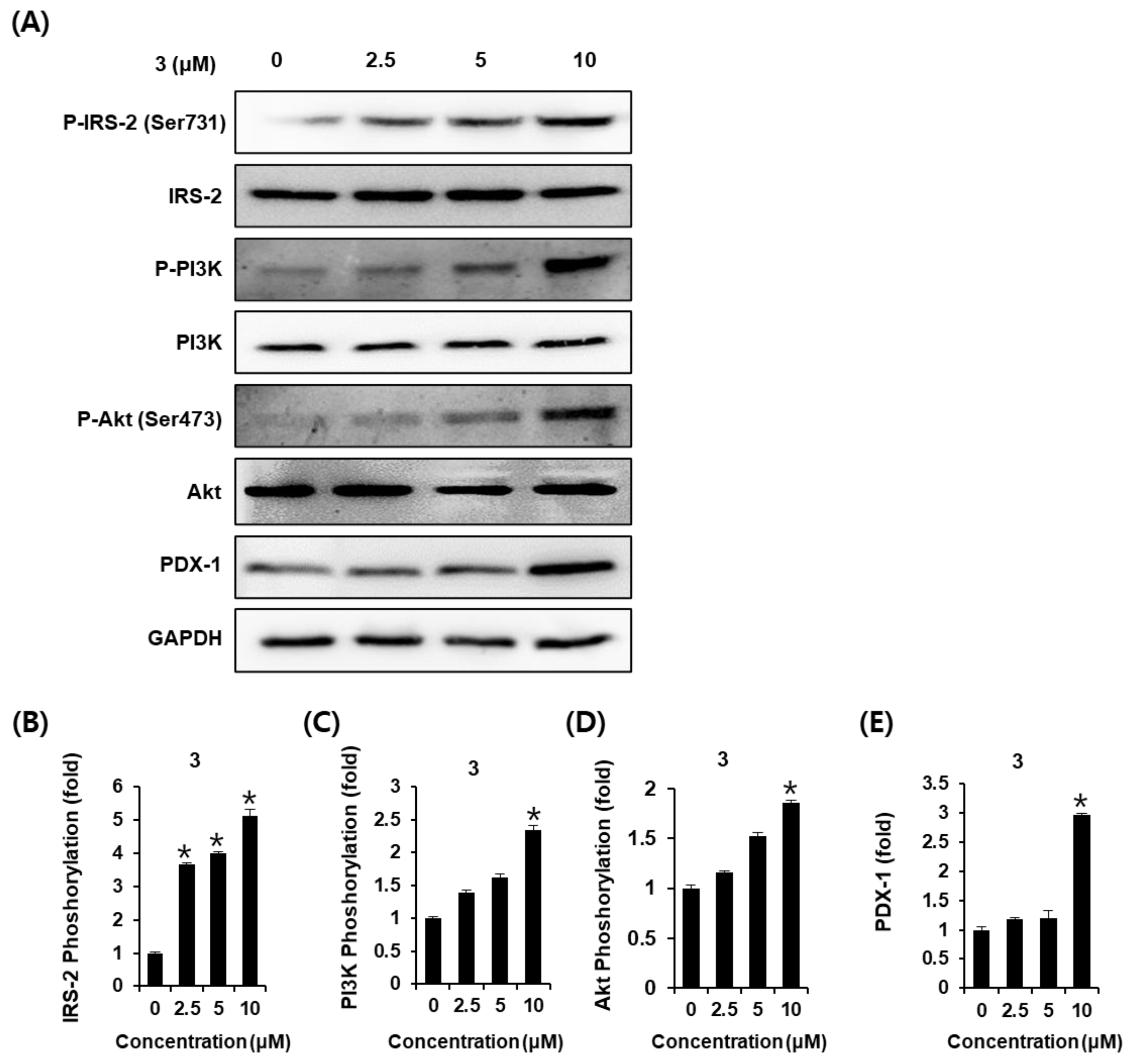
2.3. Effects of AEA (3) on the Protein Expression of P-IRS-2, IRS-2, P-PI3K, PI3K, P-Akt (Ser473), Akt, and PDX-1
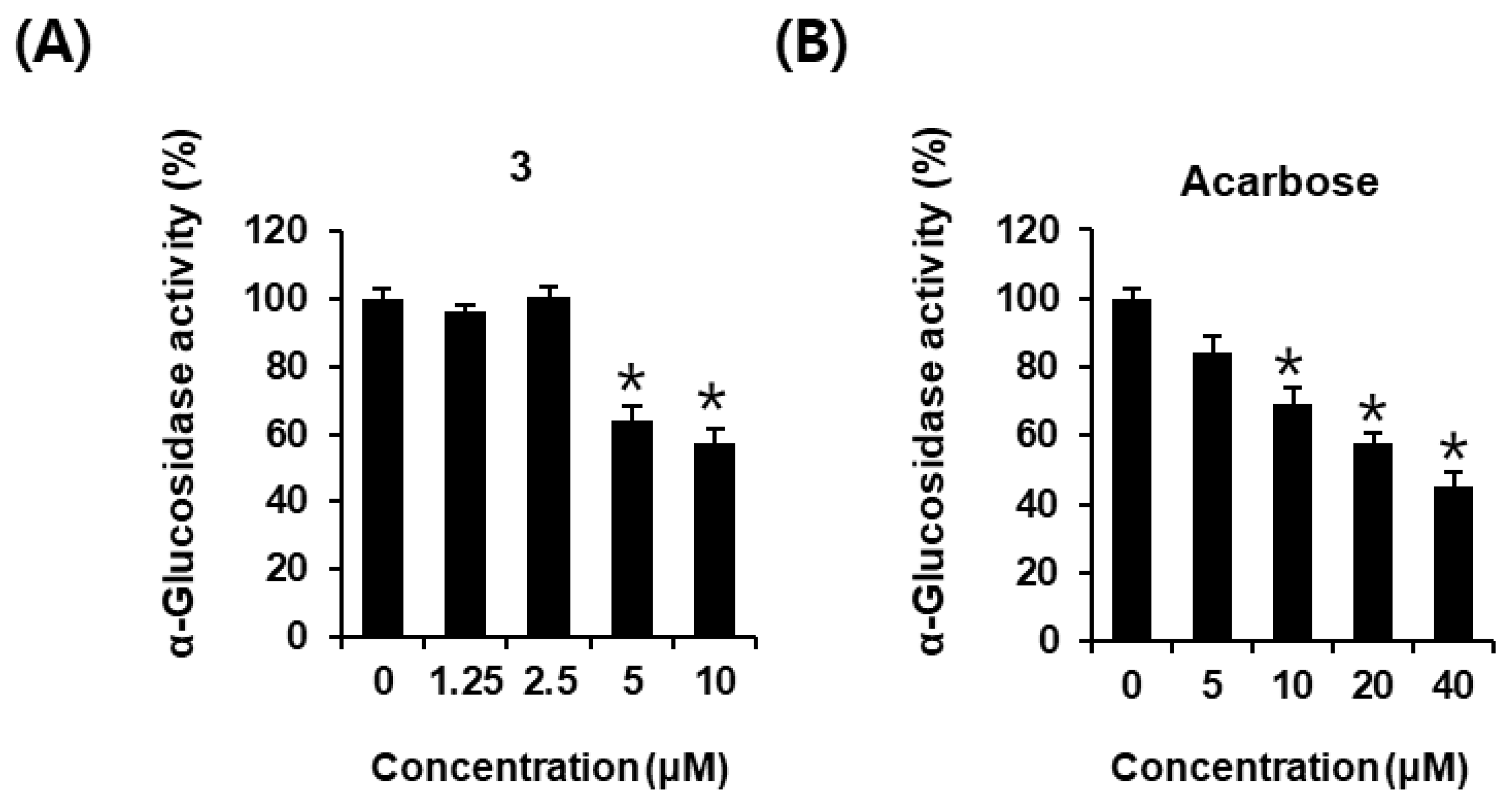
2.4. Effects of AEA (3) on α-Glucosidase Inhibitory Activity
3. Discussion
4. Materials and Methods
4.1. Isolation of Phenylpropanoids 1–7 from A. galanga
4.2. Cell Culture
4.3. Cell Viability Assay
4.4. GSIS Assay
4.5. Western Blot Analysis
4.6. Assay of α-Glucosidase Activity
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cantley, J.; Ashcroft, F.M. Q&A: Insulin secretion and type 2 diabetes: Why do β-cells fail? BMC Biol. 2015, 13, 33. [Google Scholar]
- Asif, M. The prevention and control the type-2 diabetes by changing lifestyle and dietary pattern. J. Educ. Health Promot. 2014, 3, 1. [Google Scholar] [CrossRef]
- Chaudhury, A.; Duvoor, C.; Dendi, V.S.R.; Kraleti, S.; Chada, A.; Ravilla, R.; Marco, A.; Shekhawat, N.S.; Montales, M.T.; Kuriakose, K.; et al. Clinical Review of Antidiabetic Drugs: Implications for Type 2 Diabetes Mellitus Management. Front. Endocrinol. 2017, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Seino, S.; Sugawara, K.; Yokoi, N.; Takahashi, H. β-Cell signalling and insulin secretagogues: A path for improved diabetes therapy. Diabetes Obes. Metab. 2017, 19 (Suppl. S1), 22–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soccio, R.E.; Chen, E.R.; Lazar, M.A. Thiazolidinediones and the promise of insulin sensitization in type 2 diabetes. Cell Metab. 2014, 20, 573–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Narwal, S.; Kumar, V.; Prakash, O. α-glucosidase inhibitors from plants: A natural approach to treat diabetes. Pharmacogn. Rev. 2011, 5, 19. [Google Scholar] [CrossRef] [Green Version]
- Raptis, S.; Dimitriadis, G. Oral hypoglycemic agents: Insulin secretagogues, α-glucosidase inhibitors and insulin sensitizers. Exp. Clin. Endocrinol. Diabetes 2001, 109 (Suppl. S2), S265–S287. [Google Scholar] [CrossRef]
- Shapiro, K.; Gong, W.C. Natural products used for diabetes. J. Am. Pharm. Assoc. 2002, 42, 217–226. [Google Scholar] [CrossRef]
- Coxon, G.D.; Furman, B.L.; Harvey, A.L.; McTavish, J.; Mooney, M.H.; Arastoo, M.; Kennedy, A.R.; Tettey, J.M.; Waigh, R.D. Benzylguanidines and other galegine analogues inducing weight loss in mice. J. Med. Chem. 2009, 52, 3457–3463. [Google Scholar] [CrossRef]
- Ahlina, F.N.; Nugraheni, N.; Salsabila, I.A.; Haryanti, S.; Da’i, M.; Meiyanto, E. Revealing the reversal effect of galangal (Alpinia galanga L.) extract against oxidative stress in metastatic breast cancer cells and normal fibroblast cells intended as a co-chemotherapeutic and anti-ageing agent. Asian Pac. J. Cancer Prev. 2020, 21, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Alagawadi, K. Influence of Alpinia galanga rhizomes on cafeteria diet induced obesity in rats. J. Nat. Remedies 2011, 11, 158–166. [Google Scholar]
- Singh, J.H.; Alagarsamy, V.; Diwan, P.V.; Kumar, S.S.; Nisha, J.; Reddy, Y.N. Neuroprotective effect of Alpinia galanga (L.) fractions on Aβ (25–35) induced amnesia in mice. J. Ethnopharmacol. 2011, 138, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, H.; Morikawa, T.; Managi, H.; Yoshikawa, M. Antiallergic principles from Alpinia galanga: Structural requirements of phenylpropanoids for inhibition of degranulation and release of TNF-α and IL-4 in RBL-2H3 cells. Bioorganic Med. Chem. Lett. 2003, 13, 3197–3202. [Google Scholar] [CrossRef] [PubMed]
- Haraguchi, H.; Kuwata, Y.; Inada, K.; Shingu, K.; Miyahara, K.; Nagao, M.; Yagi, A. Antifungal activity from Alpinia galanga and the competition for incorporation of unsaturated fatty acids in cell growth. Planta Med. 1996, 62, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Banerjee, M.; Bhattacharyya, N. Anti-inflammatory activity of root of Alpinia galanga willd. Chron. Young Sci. 2011, 2, 139. [Google Scholar]
- Chouni, A.; Paul, S. A review on phytochemical and pharmacological potential of Alpinia galanga. Pharmacogn. J. 2018, 10, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Kojima-Yuasa, A.; Matsui-Yuasa, I. Pharmacological Effects of 1′-Acetoxychavicol Acetate, a Major Constituent in the Rhizomes of Alpinia galanga and Alpinia conchigera. J. Med. Food 2020, 23, 465–475. [Google Scholar] [CrossRef]
- Verma, R.K.; Mishra, G.; Singh, P.; Jha, K.K.; Khosa, R.L. Anti-diabetic activity of methanolic extract of Alpinia galanga Linn. aerial parts in streptozotocin induced diabetic rats. Ayu 2015, 36, 91. [Google Scholar]
- Sumathy, S.; Anuradha, G.; Revathi, V. An In vitro Evaluation of Antidiabetic Activity of Alpinia galangal and Alpinia calcarata. J. Pharm. Res. Int. 2021, 33, 28–35. [Google Scholar]
- Fujimoto, K.; Polonsky, K.S. Pdx1 and other factors that regulate pancreatic β-cell survival. Diabetes Obes. Metab. 2009, 11, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Niessen, M. On the role of IRS2 in the regulation of functional β-cell mass. Arch. Physiol. Biochem. 2006, 112, 65–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Shangguan, Z.; Liu, Y.; Wang, J.; Li, X.; Yang, S.; Liu, S. Puerarin protects pancreatic b-cell survival via PI3K/Akt signaling pathway. J. Mol. Endocrinol. 2014, 53, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.C.; Vo, V.; Gorjala, P.; Fiscus, R.R. Pancreatic-β-cell survival and proliferation are promoted by protein kinase G type Iα and downstream regulation of AKT/FOXO1. Diabetes Vasc. Dis. Res. 2017, 14, 434–449. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-Y.; Lee, N.-K.; Wang, Y.-Y.; Hong, J.-P.; Son, S.R.; Gu, D.-H.; Jang, D.S.; Choi, J.-H. 1′-Acetoxyeugenol Acetate Isolated from Thai Ginger Induces Apoptosis in Human Ovarian Cancer Cells by ROS Production via NADPH Oxidase. Antioxidants 2022, 11, 293. [Google Scholar] [CrossRef]
- Janssen, A.; Scheffer, J. Acetoxychavicol acetate, an antifungal component of Alpinia galanga. Planta Med. 1985, 51, 507–511. [Google Scholar] [CrossRef]
- Lee, S.-J.; Ando, T. Optically active 1-acetoxychavicol acetate and its positional isomers: Synthesis and repellent effect against adzuki bean weevil. J. Pestic. Sci.-Pestic. Sci. Soc. Jpn. 2001, 26, 76–81. [Google Scholar] [CrossRef] [Green Version]
- Chua, B.; Abdullah, Z.; Pin, K.Y.; Abdullah, L.C.; Choong, T.S.Y.; Yusof, U.K. Isolation, structure elucidation, identification and quantitative analysis of 1′-acetoxychavicol (ACA) from the roots of chlorophytum boriviliuanum (SAFED MUSLI). J. Eng. Sci. Technol. 2017, 12, 198–213. [Google Scholar]
- Noro, T.; Sekiya, T.; Katoh, M.; Oda, Y.; Miyase, T.; Kuroyanagi, M.; Ueno, A.; Fukushima, S. Inhibitors of xanthine oxidase from Alpinia galanga. Chem. Pharm. Bull. 1988, 36, 244–248. [Google Scholar] [CrossRef] [Green Version]
- dos Santos, A.L.; Chierice, G.O.; Alexander, K.; Riga, A. Crystal structure determination for eugenyl acetate. J. Chem. Crystallogr. 2009, 39, 655–661. [Google Scholar] [CrossRef]
- Jiang, H.; Sun, S.-L.; Zhang, C.; Yuan, E.; Wei, Q.-Y.; Zeng, Z. Antioxidative activities of natural hydroxy-bearing cinnamaldehydes and cinnamic acids: A comparative study. Trop. J. Pharm. Res. 2013, 12, 1017–1022. [Google Scholar] [CrossRef] [Green Version]
- Sukhirun, N.; Pluempanupat, W.; Bullangpoti, V.; Koul, O. Bioefficacy of Alpinia galanga (Zingiberaceae) rhizome extracts,(E)-p-acetoxycinnamyl alcohol, and (E)-p-coumaryl alcohol ethyl ether against Bactrocera dorsalis (Diptera: Tephritidae) and the impact on detoxification enzyme activities. J. Econ. Entomol. 2011, 104, 1534–1540. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, A.; Rashidi, R.; Shafiee-Nick, R. Flavonoids for preserving pancreatic beta cell survival and function: A mechanistic review. Biomed. Pharmacother. 2019, 111, 947–957. [Google Scholar] [CrossRef]
- Banjerdpongchai, R.; Punyati, P.; Nakrob, A.; Pompimon, W.; Kongtawelert, P. 4’-Hydroxycinnamaldehyde from Alpinia galanga (Linn.) induces human leukemic cell apoptosis via mitochondrial and endoplasmic reticulum stress pathways. Asian Pac. J Cancer Prev. 2011, 12, 593–598. [Google Scholar] [PubMed]
- Taib, M.; Anuar, N.; Hanafiah, K.M.; Al-Shammary, A.A.K.; Saaid, M.; Awang, K. Chemicals Constituents Isolated from Cultivate Alpinia conchigera Griff. and Antimicrobial Activity. Trop Life Sci. Res. 2020, 31, 159–178. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Zou, L.; Wu, D.-T.; Zhuang, Q.-G.; Li, H.-B.; Mavumengwana, V.; Corke, H.; Gan, R.-Y. Discovery of 1’-acetoxychavicol acetate (ACA) as a promising antibacterial compound from galangal (Alpinia galanga (Linn.) Willd). Ind. Crops Prod. 2021, 171, 113883. [Google Scholar] [CrossRef]
- Ong, G.H.; Ori, D.; Kawasaki, T.; Kawai, T. Inhibition of lipopolysaccharide-induced inflammatory responses by 1′-acetoxychavicol acetate. Genes Cells 2022, 27, 482–492. [Google Scholar] [CrossRef]
- Williams, M.; Tietzel, I.; Quick, Q.A. 1′-Acetoxychavicol acetate promotes caspase 3-activated glioblastoma cell death by overcoming enhanced cytokine expression. Oncol. Lett. 2013, 5, 1968–1972. [Google Scholar] [CrossRef] [Green Version]
- Leibiger, B.; Wåhlander, K.; Berggren, P.-O.; Leibiger, I.B. Glucose-stimulated insulin biosynthesis depends on insulin-stimulated insulin gene transcription. J. Biol. Chem. 2000, 275, 30153–30156. [Google Scholar] [CrossRef] [Green Version]
- Goodison, S.; Ashcroft, S.J. Trans-acting factor (s) confer glucose-responsive transcriptional regulation in the insulin gene. In Physiology and Pathophysiology of the Islets of Langerhans; Springer: Cham, Switzerland, 1997; pp. 97–100. [Google Scholar]
- Gupta, D.; Kono, T.; Evans-Molina, C. The role of peroxisome proliferator-activated receptor γ in pancreatic β cell function and survival: Therapeutic implications for the treatment of type 2 diabetes mellitus. Diabetes Obes. Metab. 2010, 12, 1036–1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, D.; Jetton, T.L.; Mortensen, R.M.; Duan, S.Z.; Peshavaria, M.; Leahy, J.L. In vivo and in vitro studies of a functional peroxisome proliferator-activated receptor γ response element in the mouse pdx-1 promoter. J. Biol. Chem. 2008, 283, 32462–32470. [Google Scholar] [CrossRef] [Green Version]
- Brissova, M.; Shiota, M.; Nicholson, W.E.; Gannon, M.; Knobel, S.M.; Piston, D.W.; Wright, C.V.; Powers, A.C. Reduction in pancreatic transcription factor PDX-1 impairs glucose-stimulated insulin secretion. J. Biol. Chem. 2002, 277, 11225–11232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima-Nagata, N.; Sugai, M.; Sakurai, T.; Miyazaki, J.-i.; Tabata, Y.; Shimizu, A. Pdx-1 enables insulin secretion by regulating synaptotagmin 1 gene expression. Biochem. Biophys. Res. Commun. 2004, 318, 631–635. [Google Scholar] [CrossRef] [PubMed]
- Matveyenko, A.V.; Butler, P. Relationship between β-cell mass and diabetes onset. Diabetes Obes. Metab. 2008, 10, 23–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sesti, G.; Federici, M.; Hribal, M.L.; Lauro, D.; Sbraccia, P.; Lauro, R. Defects of the insulin receptor substrate (IRS) system in human metabolic disorders. FASEB J. 2001, 15, 2099–2111. [Google Scholar] [CrossRef] [Green Version]
- Feng, Z.-C.; Donnelly, L.; Li, J.; Krishnamurthy, M.; Riopel, M.; Wang, R. Inhibition of Gsk3 β activity improves β-cell function in c-Kit Wv/+ male mice. Lab. Investig. 2012, 92, 543–555. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.W.; Li, X.; Sun, W.L.; Xing, Y.; Xiu, Z.L.; Zhuang, C.L.; Dong, Y.S. Dietary Flavonoids and Acarbose Synergistically Inhibit α-Glucosidase and Lower Postprandial Blood Glucose. J. Agric. Food Chem. 2017, 65, 8319–8330. [Google Scholar] [CrossRef]
- Dong, Y.; Sui, L.; Yang, F.; Ren, X.; Xing, Y.; Xiu, Z. Reducing the intestinal side effects of acarbose by baicalein through the regulation of gut microbiota: An in vitro study. Food Chem. 2022, 394, 133561. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, D.; Son, S.-R.; Qi, Y.; Kang, K.S.; Jang, D.S. (1′S)-1′-Acetoxyeugenol Acetate Enhances Glucose-Stimulated Insulin Secretion. Plants 2023, 12, 579. https://doi.org/10.3390/plants12030579
Lee D, Son S-R, Qi Y, Kang KS, Jang DS. (1′S)-1′-Acetoxyeugenol Acetate Enhances Glucose-Stimulated Insulin Secretion. Plants. 2023; 12(3):579. https://doi.org/10.3390/plants12030579
Chicago/Turabian StyleLee, Dahae, So-Ri Son, Yutong Qi, Ki Sung Kang, and Dae Sik Jang. 2023. "(1′S)-1′-Acetoxyeugenol Acetate Enhances Glucose-Stimulated Insulin Secretion" Plants 12, no. 3: 579. https://doi.org/10.3390/plants12030579