

Metabolomic Profiling, Antibacterial, and Molluscicidal Properties of the Medicinal Plants Calotropis procera and Atriplex halimus: In Silico Molecular Docking Study

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Metabolic Profiling of the C. procera and A. halimus Methanol Extracts
2.2. Antibacterial Activity
2.3. Molluscicidal Activity
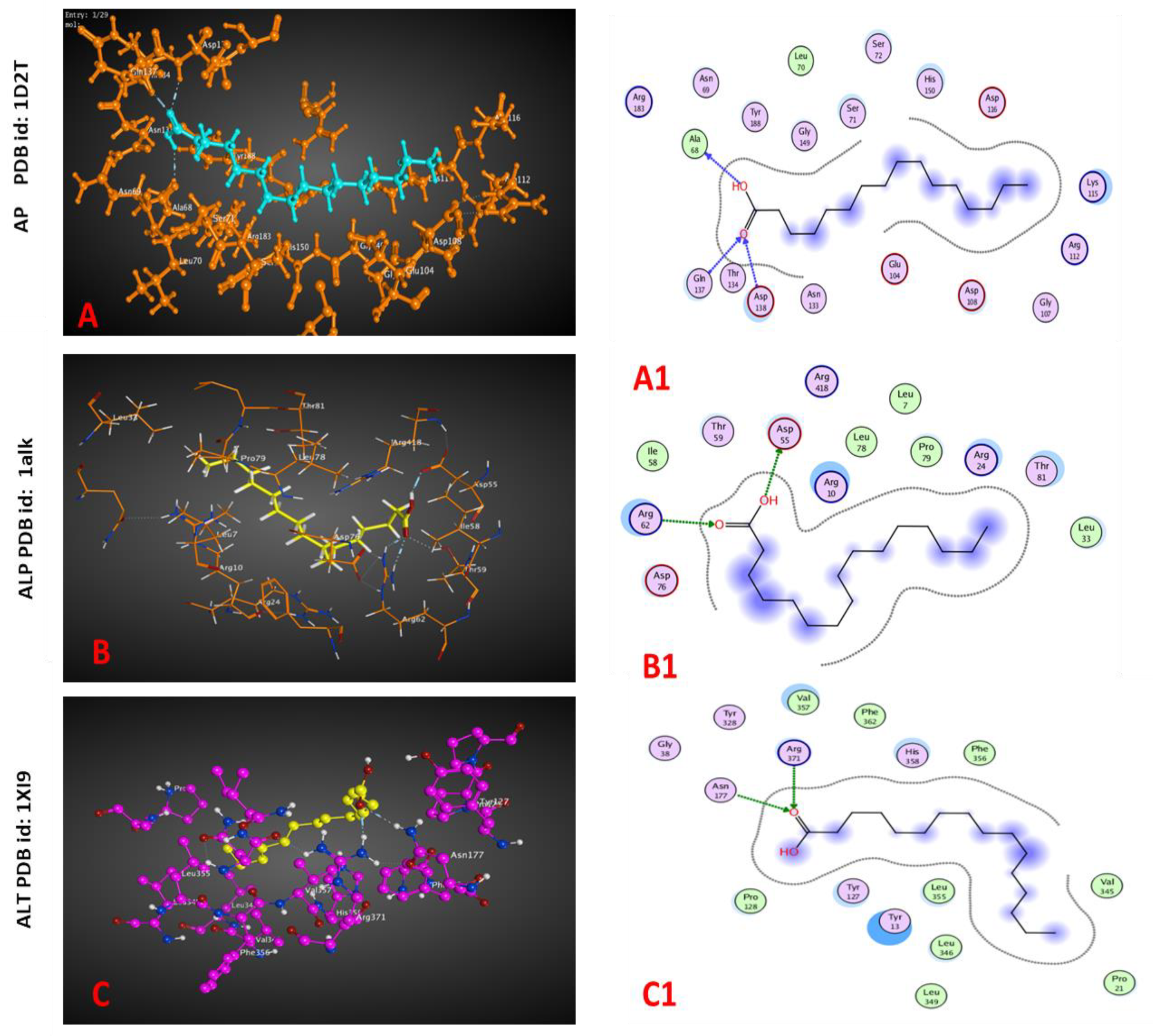
2.4. Molecular Docking
3. Discussion
4. Materials and Methods
4.1. Plants
4.1.1. Metabolites Extraction
4.1.2. Gas Chromatography-Mass Spectroscopy (GC-MS) Analysis
4.2. The Antibacterial Activities
4.3. Molluscicidal Activity
4.3.1. Snails
4.3.2. Assessment of the Molluscicidal Activity of the Plant’s Methanol Extracts
4.3.3. Effect of Plants Extract on Survival Rate of Snails
4.3.4. Effect of Plants’ Extracts on Hatchability of Snails’ Eggs
4.4. Tissue Preparation
4.4.1. Biochemical Analysis
4.4.2. Comet Assay
4.4.3. Histopathological and Immunohistochemical Analysis
4.5. The Molecular Docking Study
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Schistosomiasis. 2020. Available online: https://www.Who.Int/Newsroom/Fact-Sheets/Detail/Schistosomiasis (accessed on 10 November 2022).
- Ibrahim, A.M.; Sayed, S.S.M. Assessment of the Molluscicidal Activity of the Methanolic Seed Extracts of Ziziphus Spina-Christi and Carica Papaya on Immunological and Molecular Aspects of Biomphalaria Alexandrina Snails. Aquac. Res. 2021, 52, 2014–2024. [Google Scholar] [CrossRef]
- Loker, E.S.; Mkoji, G.M. Schistosomes and Their Snail Hosts BT—Schistosomiasis; Secor, W.E., Colley, D.G., Eds.; Springer US: Boston, MA, USA, 2005; pp. 1–11. ISBN 978-0-387-23362-8. [Google Scholar]
- Elsareh, F.; Abdalla, R.; Abdalla, E. The Effect of Aqueous Leaves Extract of Solenostemma Argel (Del Hayne) on Egg Masses and Neonates of Biomphalaria Pfeifferi Snails. J. Med. Plants 2016, 4, 271–274. [Google Scholar]
- Abd El Ghaffar, M.M.; Sadek, G.S.; Harba, N.M.; Abd El Samee, M.F. Evaluation of the Effect of Some Plant Molluscicides on the Infectivity of Schistosoma Mansoni Cercariae. Menoufia Med. J. 2019, 31, 1448–1455. [Google Scholar] [CrossRef]
- Uddin, T.M.; Chakraborty, A.J.; Khusro, A.; Zidan, B.R.M.; Mitra, S.; Emran, T.B.; Dhama, K.; Ripon, M.K.H.; Gajdács, M.; Sahibzada, M.U.K.; et al. Antibiotic Resistance in Microbes: History, Mechanisms, Therapeutic Strategies and Future Prospects. J. Infect. Public Health 2021, 14, 1750–1766. [Google Scholar] [CrossRef]
- Tagboto, S.; Townson, S. Antiparasitic Properties of Medicinal Plants and Other Naturally Occurring Products. Adv. Parasitol. 2001, 50, 199–295. [Google Scholar] [CrossRef]
- Batool, H.; Hussain, M.; Hameed, M.; Ahmad, R. A review on Calotropis procera its phytochemistry and traditional uses. Big Data Agric. 2020, 2, 56–58. [Google Scholar] [CrossRef]
- Nadeem, M.; Mumtaz, M.W.; Danish, M.; Rashid, U.; Mukhtar, H.; Anwar, F.; Raza, S.A. Calotropis Procera: UHPLC-QTOF-MS/MS Based Profiling of Bioactives, Antioxidant and Anti-Diabetic Potential of Leaf Extracts and an Insight into Molecular Docking. J. Food Meas. Charact. 1234, 13, 3206–3220. [Google Scholar] [CrossRef]
- Singh, V.S.; Dhawale, S.C.; Shakeel, F.; Faiyazuddin, M.; Alshehri, S. Antiarthritic Potential of Calotropis procera Leaf Fractions in FCA-Induced Arthritic Rats: Involvement of Cellular Inflammatory Mediators and Other Biomarkers. Agriculture 2021, 11, 68. [Google Scholar] [CrossRef]
- Darra Wadhwani, B.; Mali, D.; Vyas, P.; Nair, R.; Khandelwal, P. A Review on Phytochemical Constituents and Pharmacological Potential of Calotropis Procera. RSC Adv. 2021, 11, 35854–35878. [Google Scholar] [CrossRef]
- Walker, D.J.; Lutts, S.; Sánchez-García, M.; Correal, E. Atriplex halimus L. Its Biology and Uses. J. Arid. Environ. 2014, 100–101, 111–121. [Google Scholar] [CrossRef]
- Nedjimi, B. Heavy Metal Tolerance in Two Algerian Saltbushes: A Review on Plant Responses to Cadmium and Role of Calcium in Its Mitigation. Plant Nutr. Abiotic Stress Toler. 2018, 205–220. [Google Scholar]
- Acosta, J.A.; Abbaspour, A.; Martínez, G.R.; Martínez-Martínez, S.; Zornoza, R.; Gabarrón, M.; Faz, A. Phytoremediation of Mine Tailings with Atriplex halimus and Organic/Inorganic Amendments: A Five-Year Field Case Study. Chemosphere 2018, 204, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Clauser, M.; Dall’Acqua, S.; Loi, M.; Innocenti, G. Phytochemical Investigation on Atriplex halimus L. from Sardinia. Nat. Prod. Res. 2013, 20, 1940–1944. [Google Scholar] [CrossRef] [PubMed]
- Elbouzidi, A.; Ouassou, H.; Aherkou, M.; Kharchoufa, L.; Meskali, N.; Baraich, A.; Mechchate, H.; Bouhrim, M.; Idir, A.; Hano, C.; et al. LC–MS/MS Phytochemical Profiling, Antioxidant Activity, and Cytotoxicity of the Ethanolic Extract of Atriplex halimus L. against Breast Cancer Cell Lines: Computational Studies and Experimental Validation. Pharmaceuticals 2022, 15, 1156. [Google Scholar] [CrossRef]
- Al-Rowaily, S.L.; Abd-Elgawad, A.M.; Assaeed, A.M.; Elgamal, A.M.; El-Nasser, A.; el Gendy, G.; Mohamed, T.A.; Dar, B.A.; Mohamed, T.K.; Elshamy, A.I. Molecules Essential Oil of Calotropis Procera: Comparative Chemical Profiles, Antimicrobial Activity, and Allelopathic Potential on Weeds. Molecules 2020, 25, 5203. [Google Scholar] [CrossRef]
- Pattnaik, P.K.; Kar, D.; Chhatoi, H.; Shahbazi, S.; Ghosh, G.; Kuanar, A. Chemometric Profile & Antimicrobial Activities of Leaf Extract of Calotropis procera and Calotropis Gigantea. Nat. Prod. Res. 2017, 31, 1954–1957. [Google Scholar]
- Chundattu, S.J.; Agrawal, V.K.; Ganesh, N. Phytochemical Investigation of Calotropis Procera. Arab. J. Chem. 2016, 9, S230–S234. [Google Scholar] [CrossRef] [Green Version]
- Harada, H.; Yamashita, U.; Kurihara, H.; Fukushi, E.; Kawabata, J.; Kamei, Y. Antitumor Activity of Palmitic Acid Found as a Selective Cytotoxic Substance in a Marine Red Alga. Anticancer Res. 2002, 22, 2587–2590. [Google Scholar]
- Librán-Pérez, M.; Pereiro, P.; Figueras, A.; Novoa, B. Antiviral Activity of Palmitic Acid via Autophagic Flux Inhibition in Zebrafish (Danio Rerio). Fish Shellfish Immunol. 2019, 95, 595–605. [Google Scholar] [CrossRef]
- DeWitte-Orr, S.; Cai, J.; Huang, X.; Qin, Q.; Yu, Y.; Li, C.; Liu, J.; Zhu, F.; Wei, S.; Huang, Y. Palmitic Acid Promotes Virus Replication in Fish Cell by Modulating Autophagy Flux and TBK1-IRF3/7 Pathway. Front. Immunol. 2020, 11, 1764. [Google Scholar] [CrossRef]
- Ashraf, R.; Bhatti, H.N. Stigmasterol. In A Centum of Valuable Plant Bioactives; Academic Press: Cambridge, MA, USA, 2021; pp. 212–232. [Google Scholar] [CrossRef]
- Yusuf, A.J.; Abdullahi, M.I.; Aleku, G.A.; Ibrahim, I.A.; Alebiosu, C.O.; Yahaya, M.; Adamu, H.W.; Sanusi, A.; Mailafiya, M.M.; Abubakar, H. Antimicrobial Activity of Stigmasterol from the Stem Bark of Neocarya Macrophylla. J. Med. Plants Econ. Dev. 2018, 2, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Gade, S.; Rajamanikyam, M.; Vadlapudi, V.; Nukala, K.M.; Aluvala, R.; Giddigari, C.; Karanam, N.J.; Barua, N.C.; Pandey, R.; Upadhyayula, V.S.v.; et al. Acetylcholinesterase Inhibitory Activity of Stigmasterol & Hexacosanol Is Responsible for Larvicidal and Repellent Properties of Chromolaena odorata. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2017, 1861, 541–550. [Google Scholar] [CrossRef]
- Bhattacharjee, B.; Pal, P.K.; Chattopadhyay, A.; Bandyopadhyay, D. Oleic Acid Protects against Cadmium Induced Cardiac and Hepatic Tissue Injury in Male Wistar Rats: A Mechanistic Study. Life Sci. 2020, 244, 117324. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.-C.; Yen, P.-L.; Chang, S.-T.; Cheng, P.-L.; Lo, Y.-C.; Hsiu-Chuan Liao, V. Antioxidative Activities of Both Oleic Acid and Camellia Tenuifolia Seed Oil Are Regulated by the Transcription Factor DAF-16/FOXO in Caenorhabditis Elegans. PLoS ONE 2016, 11, e0157195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.T.; de Alencar, M.V.O.B.; da Conceição Machado, K.; da Conceição Machado, K.; de Carvalho Melo-Cavalcante, A.A.; de Sousa, D.P.; de Freitas, R.M. Phytol in a Pharma-Medico-Stance. Chem. Biol. Interact. 2015, 240, 60–73. [Google Scholar] [CrossRef]
- Islam, M.T.; Ali, E.S.; Uddin, S.J.; Shaw, S.; Islam, M.A.; Ahmed, M.I.; Chandra Shill, M.; Karmakar, U.K.; Yarla, N.S.; Khan, I.N.; et al. Phytol: A Review of Biomedical Activities. Food Chem. Toxicol. 2018, 121, 82–94. [Google Scholar] [CrossRef] [PubMed]
- do Nascimento, K.F.; Moreira, F.M.F.; Alencar Santos, J.; Kassuya, C.A.L.; Croda, J.H.R.; Cardoso, C.A.L.; Vieira, M. do C.; Góis Ruiz, A.L.T.; Ann Foglio, M.; de Carvalho, J.E.; et al. Antioxidant, Anti-Inflammatory, Antiproliferative and Antimycobacterial Activities of the Essential Oil of Psidium Guineense Sw. and Spathulenol. J. Ethnopharmacol. 2018, 210, 351–358. [Google Scholar] [CrossRef]
- Ounaissia, K.; Bennadja, S.; Aliane, L.; Djahoudi, A. Phytochemical screening and anti-bacterial activity of methanolic extracts of the aerial parts of Atriplex halimus L., from biskra (Algeria). Asian J. Res. Chem. 2020, 13, 365–372. [Google Scholar]
- Kabbash, A.; Shoeib, N. Chemical and Biological Investigation of Some Secondary Metabolites in Atriplex halimus Growing in Egypt. Nat. Prod. Commun. 2012, 7, 1465–1468. [Google Scholar] [CrossRef] [Green Version]
- Zennaf, I.; Tir touil, A.; Meddah, B.; Mokhtar, M. Ethnobotanical and Phytochemical Study of the Medicinal Plant Atriplex halimus and Its Importance in the Traditional Algerian Pharmacopoeia. Fr.-Ukr. J. Chem. 2022, 10, 60–69. [Google Scholar] [CrossRef]
- Chhetri, D.R. Myo-Inositol and Its Derivatives: Their Emerging Role in the Treatment of Human Diseases. Front. Pharmacol. 2019, 10, 1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Hidalgo, M.; León-González, A.J.; Gálvez-Peralta, M.; González-Mauraza, N.H.; Martin-Cordero, C. d-Pinitol: A cyclitol with versatile biological and pharmacological activities. Phytochem. Rev. 2020, 20, 211–224. [Google Scholar] [CrossRef]
- Luo, J.; Huang, K.; Zhou, X.; Xu, Y. Preparation of Highly Flexible and Sustainable Lignin-Rich Nanocellulose Film Containing Xylonic Acid (XA), and Its Application as an Antibacterial Agent. Int. J. Biol. Macromol. 2020, 163, 1565–1571. [Google Scholar] [CrossRef] [PubMed]
- Rossi, E.; La Rosa, R.; Bartell, J.A.; Marvig, R.L.; Haagensen, J.A.; Sommer, L.M.; Molin, S.; Johansen, H.K. Pseudomonas aeruginosa adaptation and evolution in patients with cystic fibrosis. Nat. Rev. Microbiol. 2021, 19, 331–342. [Google Scholar] [CrossRef]
- Armbruster, C.E.; Mobley, H.L.T.; Pearson, M.M. Pathogenesis of Proteus Mirabilis Infection. EcoSal Plus 2018, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Zanichelli, V.; Huttner, A.; Harbarth, S.; Kronenberg, A.; Huttner, B. Swiss Centre For Antibiotic Resistance Anresis. Antimicrobial Resistance Trends in Escherichia coli, Klebsiella pneumoniae and Proteus mirabilis Urinary Isolates from Switzerland: Retrospective Analysis of Data from a National Surveillance Network over an 8-Year Period (2009–2016). Swiss Med. Wkly. 2019, 149, w20110. [Google Scholar] [CrossRef] [Green Version]
- Papagiannitsis, C.C.; Hrabak, J.; Maria D’andrea, M.; Reslan, L.; Naas, T.; Girlich, D.; Bonnin, R.A.; Dortet, L. Genetics of Acquired Antibiotic Resistance Genes in Proteus spp. Front. Microbiol. 2020, 11, 256. [Google Scholar] [CrossRef] [Green Version]
- Nenaah, G. Antimicrobial activity of Calotropis procera Ait. (Asclepiadaceae) and isolation of four flavonoid glycosides as the active constituents. World J. Microbiol. Biotechnol. 2013, 29, 1255–1262. [Google Scholar] [CrossRef]
- Bakry, F.A. Use of Some Plant Extracts to Control Biomphalaria Alexandrina Snails with Emphasis on Some Biological Effects. Pestic. Biochem. Physiol. 2009, 95, 159–165. [Google Scholar] [CrossRef]
- Abo Zaid, K.H.; El-Wakil, H.; El-Hussein, A.; Jomaa, S.; Shohayeb, M. Evaluation of the Molluscicidal Activity of Punica Granatum, Calotropis procera, Solanum incanum and Citrullus colocynthis against Biomphalaria arabica. World Appl. Sci. J. 2013, 26, 873–879. [Google Scholar] [CrossRef]
- Seif El-Din, S.H.; El-Lakkany, N.M.; Mohamed, M.A.; Hamed, M.M.; Sterner, O.; Botros, S.S. Potential Effect of the Medicinal Plants Calotropis procera, Ficus Elastica and Zingiber Officinale against Schistosoma Mansoni in Mice. Pharm Biol. 2014, 52, 144–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mettwally, W.S.A.; Zahran, H.A.; Khayyal, A.E.; Ahmed, M.M.E.; Allam, R.M.; Saleh, D.O. Calotropis procera (Aiton) Seeds Fixed Oil: Physicochemical Analysis, GC–MS Profiling and Evaluation of Its in-Vivo Anti-Inflammatory and in-Vitro Antiparasitic Activities. Arab. J. Chem. 2022, 15, 104085. [Google Scholar] [CrossRef]
- Ali, B.; Musaddiq, S.; Iqbal, S.; Rehman, T.; Shafiq, N.; Hussain, A. The Therapeutic Properties, Ethno Pharmacology and Phytochemistry of Atriplex Species: A Review. Pak. J. Biochem. Biotechnol. 2021, 2, 49–64. [Google Scholar] [CrossRef]
- Ibrahim, A.M.; Ahmed, A.K.; Bakry, F.A.; Abdel-Ghaffar, F. Hematological, Physiological and Genotoxicological Effects of Match 5% EC Insecticide on Biomphalaria Alexandrina Snails. Ecotoxicol. Env. Saf. 2018, 147, 1017–1022. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.F.; Steinert, S. Use of the Single Cell Gel Electrophoresis/Comet Assay for Detecting DNA Damage in Aquatic (Marine and Freshwater) Animals. Mutat. Res./Rev. Mutat. Res. 2003, 544, 43–64. [Google Scholar] [CrossRef] [PubMed]
- Harris, L.; O’Byrne-Ring, N.; Lambkin, H. Characterisation Of Cell Types In Abalone (Haliotis spp.) Tissues Using Immunohistochemical Techniques. Articles 2006, 261, 1413–1421. [Google Scholar] [CrossRef] [Green Version]
- Nakopoulou, L.; Janinis, J.; Panagos, G.; Comin, G.; Davaris, P. The Immunohistochemical Expression of Proliferating Cell Nuclear Antigen (PCNA/Cyclin) in Malignant and Benign Epithelial Ovarian Neoplasms and Correlation with Prognosis. Eur. J. Cancer 1993, 29, 1599–1601. [Google Scholar] [CrossRef]
- Zhang, L.; Zou, Z. Molluscicidal Activity of Fatty Acids in the Kernel of Chimonanthus Praecox Cv. Luteus against the Golden Apple Snail Pomacea Canaliculata. Pestic. Biochem. Physiol. 2020, 167, 104620. [Google Scholar] [CrossRef]
- Ohta, Y.; Ichimura, K. Proliferation Markers, Proliferating Cell Nuclear Antigen, Ki67, 5-Bromo-2′-Deoxyuridine, and Cyclin D1 in Mouse Olfactory Epithelium. Ann. Otol. Rhinol. Laryngol. 2000, 109, 1046–1048. [Google Scholar] [CrossRef]
- Alao, J.P. The Regulation of Cyclin D1 Degradation: Roles in Cancer Development and the Potential for Therapeutic Invention. Mol. Cancer 2007, 6, 24. [Google Scholar] [CrossRef] [Green Version]
- Bartkova, J.; Lukas, J.; Strauss, M.; Bartek, J. Cell Cycle-Related Variation and Tissue-Restricted Expression of Human Cyclin D1 Protein. J. Pathol. 1994, 172, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.M.; Bakry, F.A. Assessment of the Molluscicidal Impact of Extracted Chlorophyllin on Some Biochemical Parameters in the Nervous Tissue and Histological Changes in Biomphalaria Alexandrina and Lymnaea Natalensis Snails. Invertebr. Neurosci. 2019, 19, 7. [Google Scholar] [CrossRef] [PubMed]
- Abonyi, C.U.; Ezugwu, A.L.; Eze, J.C.; Nwankwo, N.E.; Okafor, F.C. Toxicological Impact of Picralima Nitida (Pile Plant) Extracts on the Gastropod Lanistes Varicus (Freshwater Snail), as a Control Measure against Trematodes Infections. Biologia 2022, 77, 2093–2101. [Google Scholar] [CrossRef]
- Saad, A.-H.A.; Aly, R.H.; Ragab, F.M.A.; Abdel-Wareth, M.T.A. Effect of Cestrum diurnum and Casimiroa edulis plants on the digestive gland and its related enzymes in the freshwater snail Biomphalaria alexandrina. Toxicol. Environ. Health Sci. 2012, 4, 109–114. [Google Scholar] [CrossRef]
- Boulos, L. Flora of Egypt; Al Hadara Publishing: Cairo, Egypt, 2002; Volume 3. [Google Scholar]
- Hamad, D.; El-Sayed, H.; Ahmed, W.; Sonbol, H.; Ramadan, M.A.H. GC-MS Analysis of Potentially Volatile Compounds of Pleurotus Ostreatus Polar Extract: In Vitro Antimicrobial, Cytotoxic, Immunomodulatory, and Antioxidant Activities. Front. Microbiol. 2022, 13, 396. [Google Scholar] [CrossRef]
- WHO. Report of the Scientific Working Group on Plant Molluscicide & Guidelines for Evaluation of Plant Molluscicides; WHO: Geneva, Switzerland, 1983. [Google Scholar]
- WHO Molluscicide Screening and Evaluation. Bull. WHO 1965, 33, 567–581.
- Bergmeyer, H. Methods of Enzymatic Analysis, Volume 8: Metabolites 3: Lipids, Amino Acids and Related Compounds; Verlag Chemie: Basel, Switzerland, 1985. [Google Scholar]
- Singh, D.K.; Agarwal, R.A. Action Sites of Cypermethrin, a Synthetic Pyrethroid in the Snail Lymnaea Acuminata. Acta Hydrochim. Hydrobiol. 1991, 19, 425–430. [Google Scholar] [CrossRef]
- Doumas, B.T. Standards for Total Serum Protein Assays—A Collaborative Study. Clin. Chem. 1975, 21, 1159–1166. [Google Scholar] [CrossRef]
- Gustafsson, J. Improved Specificity of Serum Albumin Determination and Estimation of “Acute Phase Reactants” by Use of the Bromcresol Green Reaction. Clin. Chem. 1976, 22, 616–622. [Google Scholar] [CrossRef]
- Reitman, S.; Frankel, S. A Colorimetric Method for the Determination of Serum Glutamic Oxalacetic and Glutamic Pyruvic Transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef]
- Grazeffe, V.S.; Tallarico, L.D.F.; Pinheiro, A.D.S.; Kawano, T.; Suzuki, M.; Okazaki, K.; Pereira, C.A.D.B.; Nakano, E. Establishment of the Comet Assay in the Freshwater Snail Biomphalaria Glabrata (Say, 1818). Mutat. Res. /Genet. Toxicol. Environ. Mutagen. 2008, 654, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Carleton, M.; Drury, R.A.B.; Willington, E.A.; Cammeron, R. Carleton’s Histological Technique; Oxford University Press: New York, NY, USA, 1967. [Google Scholar]
- Ishikawa, K.; Mihara, Y.; Gondoh, K.; Suzuki, E.; Asano, Y. X-ray structures of a novel acid phosphatase from Escherichia blattae and its complex with the transition-state analog molybdate. EMBO J. 2000, 19, 2412–2423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.E.; Wyckoff, H.W. Reaction mechanism of alkaline phosphatase based on crystal structures: Two-metal ion catalysis. J. Mol. Biol. 1991, 218, 449–464. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Tempel, W.; Shah, A.; Chen, L.; Liu, Z.-J.; Lee, D.; Lin, D.; Chang, S.-H.; Brereton, P.S.; Izumi, M.; et al. Alanine Aminotransferase from Pyrococcus furiosus Pfu-1397077-001. Southeast Collaboratory for Structural Genomics (SECSG). 2004. Available online: https://www.wwpdb.org/pdb?id=pdb_00001xi9 (accessed on 10 November 2022).




{kind=link}
{kind=link}
{kind=link}
| Compound Name | Molecular Formula | Molecular Weight (g/mol) | Retention Time (min) | Area % | |
|---|---|---|---|---|---|
| 1 | Formaldehyde | CH2O | 30.02 | 4.16 | 1.3 |
| 2 | Pentitol | C5H12O5 | 152.15 | 5.42 | 0.3 |
| 3 | L-Valine | C5H11NO2 | 117.15 | 6.59 | 0.64 |
| 4 | L-Leucine | C6H13NO2 | 131.17 | 8.25 | 0.28 |
| 5 | L-Isoleucine | C6H13NO2 | 131.17 | 8.76 | 0.35 |
| 6 | Glycerol | C3H8O3 | 92.09 | 11.37 | 2.65 |
| 7 | Butanedioic acid | C4H6O4 | 118.09 | 12.35 | 6.19 |
| 8 | Malic acid | C4H6O5 | 134.09 | 16.76 | 2.34 |
| 9 | L-Proline | C5H9NO2 | 115.13 | 17.33 | 1.21 |
| 10 | DL-Phenylalanine | C9H11NO2 | 165.19 | 17.89 | 0.83 |
| 11 | Spathulenol | C15H24O | 220 | 18.71 | 0.71 |
| 12 | Methyl-β-D-glucopyranoside | C7H14O6 | 194.18 | 20.48 | 0.26 |
| 13 | L-Fucitol | C6H14O5 | 166.17 | 22.42 | 0.58 |
| 14 | 4-Coumaric acid | C9H8O3 | 164.16 | 23.36 | 0.23 |
| 15 | Azelaic acid | C9H16O4 | 188.22 | 23.61 | 0.21 |
| 16 | D-Fructofuranose | C6H12O6 | 180.16 | 23.80 | 2.40 |
| 17 | D-Tagatofuranose | C6H12O6 | 180.16 | 23.97 | 0.45 |
| 18 | Methyl D-glucofuranoside | C7H14O6 | 194.18 | 24.60 | 0.75 |
| 19 | α-L-Arabinopyranose | C5H10O5 | 150.130 | 24.68 | 0.32 |
| 20 | Dulcitol | C6H14O6 | 182.17 | 25.51 | 0.36 |
| 21 | α-d-glucopyranose | C6H12O6 | 180.16 | 25.86 | 0.52 |
| 22 | Hexadecanoic acid, methyl ester | C17H34O2 | 270.4 | 26.19 | 0.23 |
| 23 | D-Xylofuranose | C5H10O5 | 150.13 | 26.96 | 0.54 |
| 24 | α-D-Allopyranose | C6H12O6 | 180.16 | 27.41 | 0.31 |
| 25 | D-Allofuranose | C6H12O6 | 180.16 | 27.60 | 0.45 |
| 26 | Palmitic Acid | C16H32O2 | 256.42 | 28.52 | 10.74 |
| 27 | 10-Octadecenoic acid, methyl ester | C19H36O3 | 312.5 | 29.47 | 0.51 |
| 28 | Heptadecanoic acid | C17H34O2 | 270.5 | 30.31 | 0.32 |
| 29 | Phytol | C20H40O | 296.5 | 30.82 | 3.91 |
| 30 | 9,12-Octadecadienoic acid (alpha-Linoleic acid) | C18H32O2 | 280.4 | 31.42 | 1.79 |
| 31 | Oleic Acid | C18H34O2 | 282.46 | 31.54 | 8.04 |
| 32 | Petroselinic acid | C18H34O2 | 282.46 | 31.68 | 0.92 |
| 33 | Stearic acid | C18H36O2 | 284.48 | 32.04 | 4.24 |
| 34 | 5,8,11-Eicosatriynoicacid | C20H28O2 | 300.4 | 32.76 | 0.30 |
| 35 | D-Trehalose | C12H22O11 | 342.3 | 36.63 | 3.23 |
| 36 | D-(+)-Turanose | C12H22O11 | 342.3 | 37.15 | 1.82 |
| 37 | Sucrose | C12H22O11 | 342.3 | 37.61 | 3.29 |
| 38 | Dasycarpidan-1-methanol, acetate (ester) | C20H26N2O2 | 326.4 | 39.17 | 1.46 |
| 39 | Trilinolein | C57H98O6 | 879.4 | 39.81 | 0.25 |
| 40 | Dasycarpidan-1-methanol, acetate (ester) | C20H26N2O2 | 326 | 40.02 | 0.52 |
| 41 | Oleic acid, eicosyl ester | C38H74O2 | 562.9 | 40.47 | 0.41 |
| 42 | Ser-Asp-Gly-Arg-Gly | C17H30N8O9 | 490 | 41.47 | 0.96 |
| 43 | Ursolic aldehyde | C30H48O2 | 440.7 | 42.45 | 1.31 |
| 44 | 2-Butenoic acid, 2-methyl-, 2-(acetyloxy)-1,1a,2,3,4,6,7,10,11,11 a-decahydro-7,10-dihydroxy-1,1,3,6, 9-pentamethyl-4a,7a-epoxy-5H-cyclo penta[a]cyclopropa[f]cycloundecen-1 1-yl ester | C27H38O8 | 490 | 43.02 | 0.22 |
| 45 | α-Tocopherol | C29H50O2 | 430.7 | 43.11 | 1.34 |
| 46 | L-Arabinitol pentaacetate | C15H22O10 | 362 | 43.44 | 0.29 |
| 47 | α-Carotene | C40H56 | 536 | 43.98 | 1.76 |
| 48 | Campesterol | C28H48O | 400.7 | 44.67 | 8.13 |
| 49 | Stigmasterol | C29H48O | 412.7 | 45.08 | 8.48 |
| 50 | Oleyl oleate | C36H68O2 | 532.9 | 45.41 | 1.41 |
| 51 | (Z)-Icos-11-en-1-yl oleate | C38H72O2 | 560.9 | 45.59 | 0.43 |
| 52 | 2-Hydroxy-3-[(9E)-9-octadecenoyloxy] propyl | C39H72O5 | 620.5 | 45.76 | 0.22 |
| Total | 90.7% |
| Compound Name | Molecular Formula | Molecular Weight | Retention Time (min) | Area % | |
|---|---|---|---|---|---|
| 1 | Propionic acid | C3H6O2 | 74.07 | 5.98 | 0.65 |
| 2 | Glycolic acid | C2H4O3 | 76.05 | 6.35 | 0.35 |
| 3 | L-Alanine | C3H7NO2 | 89.09 | 6.97 | 1.18 |
| 4 | Hydracrylic acid | C3H6O3 | 90.08 | 7.99 | 0.26 |
| 5 | L-Valine | C5H11NO2 | 117.15 | 9.77 | 0.31 |
| 6 | Urea | CH4N2O | 60.05 | 10.58 | 0.85 |
| 7 | Glycerol | C3H8O3 | 92.09 | 11.37 | 3.43 |
| 8 | L-Proline | C5H9NO2 | 115.13 | 11.79 | 0.64 |
| 9 | Butanedioic acid | C4H6O4 | 118.09 | 12.35 | 2.04 |
| 10 | L-Serine | C3H7NO3 | 105.09 | 13.56 | 0.38 |
| 11 | Homoserine | C4H9NO3 | 119.12 | 15.78 | 0.23 |
| 12 | L-5-Oxoproline | C5H7NO3 | 129.11 | 17.34 | 1.58 |
| 13 | L-Aspartic acid | C4H7NO4 | 133.1 | 17.49 | 0.23 |
| 14 | Methyl alpha-D-galactopyranoside | C7H14O6 | 194.18 | 17.75 | 0.32 |
| 15 | L-Threonic acid | C4H8O5 | 136.1 | 18.06 | 0.26 |
| 16 | 2,3,4-Trihydroxybutyric acid | C4H8O5 | 136.1 | 18.49 | 0.76 |
| 17 | Spathulenol | C15H24O | 220.35 | 18.71 | 0.61 |
| 18 | L-Asparagine | C4H8N2O3 | 132.12 | 19.20 | 0.45 |
| 19 | Pentanedioic acid | C5H8O4 | 132.11 | 19.37 | 0.36 |
| 20 | L-Phenylalanine | C9H11NO2 | 165.19 | 19.76 | 0.21 |
| 21 | Xylonic acid | C5H10O6 | 166.13 | 19.86 | 0.91 |
| 22 | D-(+)-Arabitol | C5H12O5 | 152.15 | 21.51 | 0.36 |
| 23 | L-Fucitol | C6H14O5 | 166.17 | 22.41 | 0.28 |
| 24 | Ribonic acid | C5H10O6 | 166.13 | 22.68 | 0.75 |
| 25 | L-(+)-Tartaric acid | C4H6O6 | 150.09 | 22.93 | 0.28 |
| 26 | D-Xylofuranose | C5H10O5 | 150.13 | 23.06 | 0.30 |
| 27 | D-Pinitol | C7H14O6 | 194.18 | 23.14 | 1.63 |
| 28 | α -D-Glucopyranosiduronic acid | C42H71NO19 | 894 | 23.61 | 0.60 |
| 29 | D-(-)-Fructofuranose | C6H12O6 | 180.16 | 23.80 | 2.72 |
| 30 | D-Psicofuranose | C6H12O6 | 180.16 | 23.9 | 2.54 |
| 31 | Citric acid | C6H8O7 | 192.12 | 24.10 | 4.05 |
| 32 | Myo-Inositol | C6H12O6 | 180.16 | 24.25 | 5.14 |
| 33 | Methyl-D-glucofuranoside | C7H14O6 | 194.18 | 24.60 | 2.20 |
| 34 | D-Mannonic acid | C6H12O7 | 196.16 | 24.98 | 1.52 |
| 35 | α-D-(+)-Talopyranose | C6H12O6 | 180.16 | 25.54 | 1.51 |
| 36 | 1,5-Anhydrohexitol | C6H12O5 | 164.16 | 25.69 | 0.32 |
| 37 | α –Lyxopyranose | C5H10O5 | 150.13 | 25.86 | 0.61 |
| 38 | Methyl palmitate | C17H34O2 | 270.5 | 26.20 | 0.75 |
| 39 | D-Lyxofuranose | C5H10O5 | 150.13 | 26.95 | 1.03 |
| 40 | D-(+)-Talofuranose | C6H12O6 | 180.16 | 27.75 | 0.44 |
| 41 | Palmitic Acid | C16H32O2 | 256.42 | 28.52 | 6.47 |
| 42 | D-Allofuranose | C6H12O6 | 180.16 | 28.92 | 0.42 |
| 43 | Linoleic acid ethyl ester | C20H36O2 | 308 | 29.32 | 0.80 |
| 44 | cis-13-Octadecenoic acid, methyl ester | C19H36O2 | 296 | 29.46 | 1.21 |
| 45 | Methyl stearate | C19H38O2 | 298 | 29.98 | 0.36 |
| 46 | Phytol | C20H40O | 296.5 | 30.82 | 0.25 |
| 47 | 9,12-Octadecadienoic acid | C18H32O2 | 280.4 | 31.42 | 1.55 |
| 48 | Oleic Acid | C18H34O2 | 282.5 | 31.54 | 5.25 |
| 49 | cis-11-Octadecenoic acid | C18H34O2 | 282.5 | 31.68 | 0.82 |
| 50 | Stearic acid | C18H36O2 | 284.5 | 32.05 | 4.01 |
| 51 | Linoelaidic acid | C18H32O2 | 280.4 | 33.07 | 0.32 |
| 52 | D-(+)-Galacturonic acid | C6H10O7 | 194.14 | 34.36 | 0.33 |
| 53 | 11-Eicosenoic acid | C20H38O2 | 310.5 | 34.86 | 0.42 |
| 54 | á-D-Galactopyranoside | C6H12O6 | 180.16 | 35.30 | 0.33 |
| 55 | Sucrose | C12H22O11 | 342.3 | 36.63 | 2.24 |
| 56 | D-Trehalose | C12H22O11 | 342.3 | 37.15 | 0.85 |
| 57 | Oleic acid, eicosyl ester | C38H74O2 | 562 | 39.10 | 0.45 |
| 58 | Dasycarpidan-1-methanol, acetate (ester) | C20H26N2O2 | 326 | 39.17 | 0.19 |
| 59 | Fumaric acid | C4H4O4 | 116.07 | 39.22 | 0.19 |
| 60 | 2-Oleoylglycerol | C21H40O4 | 356.5 | 39.32 | 0.42 |
| 61 | 2-Hydroxy-3-[(9E)-9-octadecenoyloxy]propyl(9E)-9-octadecenoate | C39H72O5 | 620 | 40.03 | 0.45 |
| 62 | Dasycarpidan-1-methanol, acetate (ester) | C20H26N2O2 | 326 | 40.74 | 0.54 |
| 63 | 9-Octadecenoic acid, (2-phenyl-1,3-dioxolan-4-YL) Methyl ester | C28H44O4 | 444 | 43.14 | 0.50 |
| 64 | Stigmasterol | C29H48O | 412.7 | 45.06 | 0.36 |
| 65 | (Z)-Icos-11-en-1-yl oleate | C38H72O2 | 560 | 45.39 | 1.11 |
| 66 | E,E,Z-1,3,12-Nonadecatriene-5,14-d Iol | C19H34O2 | 294 | 45.62 | 0.83 |
| Total | 73.7% |
| Bacterial Species | Inhibition Clear Zone Diameter (mm) | |||
|---|---|---|---|---|
| Atriplex halimus | Calitropis procera | Gentamycin (10 μg/disc) | Ethyl Acetate | |
| Staphylococcus aureus ATCC 25923 | -ve | -ve | 17 ± 0.2 | -ve |
| Escherichia coli ATCC 25922 | -ve | 10 ± 0.1 b | 15 ± 0.8 a | -ve |
| Pseudomonas aeruginosa ATCC 7853 | 14 ± 0.5 c | 18 ± 0.3 a | 17 ± 0.6 b | -ve |
| Proteus mirabilis ATCC 29906 | -ve | 18 ± 0.2 a | 10 ± 0.0 b | -ve |
| Klebsiella pneumoniae ATCC 700721 | -ve | -ve | 12 ± 0.5 | -ve |
| Slope | LC90 (mg/L) | LC50 (mg/L) | LC25 (mg/L) | LC10 (mg/L) | Plants |
|---|---|---|---|---|---|
| 1.1 | 260.4 | 223.8 | 204.5 | 187.2 | Atriplex halimus |
| 1.0 | 148.5 | 135 | 127.8 | 121.4 | Calitropis procera |
| Weeks | Survival Rate (%) | ||
|---|---|---|---|
| Control | A. halimus | C. procera | |
| 1 | 99 c | 80 b | 55 a |
| 2 | 95 c | 60 b | 30 a |
| 3 | 95 c | 40 b | 15 a |
| 4 | 90 c | 20 b | 5 a |
| Group | % Hatchability | % Mortality |
|---|---|---|
| Control | 100 c | 0 c |
| Atriplex halimus | 60 b | 40 b |
| Calitropis procera | 30 a | 70 a |
| Olive Tail Moment | Tail Length (px) | % DNA in Tail | Tail Moment | |
|---|---|---|---|---|
| Control | 1.71 | 4.62 ± 0.58 c | 16.39 ± 4.25 b | 0.94 ± 0.31 c |
| Atriplex halimus (LC25) | 2.11 | 6.24 ± 0.12 b | 16.21 ± 1.11 b | 1.23 ± 1.13 b |
| Calotropis procera(LC25) | 2.99 | 8.35 ± 0.92 a | 20.25 ± 0.21 a | 2.14 ± 0.72 a |
| Alkaline Phosphatase (μmole/mg) | Acid Phosphatase (μmole/mg) | Total Protein g/100 mL | Albumin g/100 mL | Alanine Aminotransfersa (ALT) U/L | |
|---|---|---|---|---|---|
| Control | 105.7 ± 0.05 c | 125 ± 0.2 b | 5.8 ± 0.11 b | 3.4 ± 0.1 b | 68.2 ± 0.5 c |
| LC25 Atriplex halimus | 75.4 ± 0.1 b | 95.2 ± 0.4 a | 3.9 ± 0.12 a | 3.1 ± 0.1 b | 88.5 ± 0.6 b |
| LC25 Calotropis procera | 60.5 ± 0.3 a | 80.5 ± 0.2 a | 3.6 ± 0.23 a | 2.4 ± 0.3 a | 107.2 ± 0.4 a |
| PDB ID | Docking Score (Kcal/mol) | Interaction Type | Amino Acid Residue Involved in Docking |
|---|---|---|---|
| AP (1D2T) | −1.1 | H-donor H-acceptor H-acceptor | ALA 68 |
| −4.3 | GLN 137 | ||
| −2.0 | ASP 138 | ||
| ALP (1alk) | −6.3 | H- donor H-acceptor H-acceptor | ASP 55 ARG 62 ARG 62 |
| −2.6 | |||
| −2.1 | |||
| ALT (1XI9) | −0.7 −3.1 | H-acceptor H-acceptor | ASN 177 ARG 371 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morad, M.Y.; El-Sayed, H.; El-Khadragy, M.F.; Abdelsalam, A.; Ahmed, E.Z.; Ibrahim, A.M. Metabolomic Profiling, Antibacterial, and Molluscicidal Properties of the Medicinal Plants Calotropis procera and Atriplex halimus: In Silico Molecular Docking Study. Plants 2023, 12, 477. https://doi.org/10.3390/plants12030477
Morad MY, El-Sayed H, El-Khadragy MF, Abdelsalam A, Ahmed EZ, Ibrahim AM. Metabolomic Profiling, Antibacterial, and Molluscicidal Properties of the Medicinal Plants Calotropis procera and Atriplex halimus: In Silico Molecular Docking Study. Plants. 2023; 12(3):477. https://doi.org/10.3390/plants12030477
Chicago/Turabian StyleMorad, Mostafa Y., Heba El-Sayed, Manal F. El-Khadragy, Asmaa Abdelsalam, Eman Zakaria Ahmed, and Amina M. Ibrahim. 2023. "Metabolomic Profiling, Antibacterial, and Molluscicidal Properties of the Medicinal Plants Calotropis procera and Atriplex halimus: In Silico Molecular Docking Study" Plants 12, no. 3: 477. https://doi.org/10.3390/plants12030477