
Identification of miRNAs Involving Potato-Phytophthora infestans Interaction

Abstract
:1. Introduction
2. Results
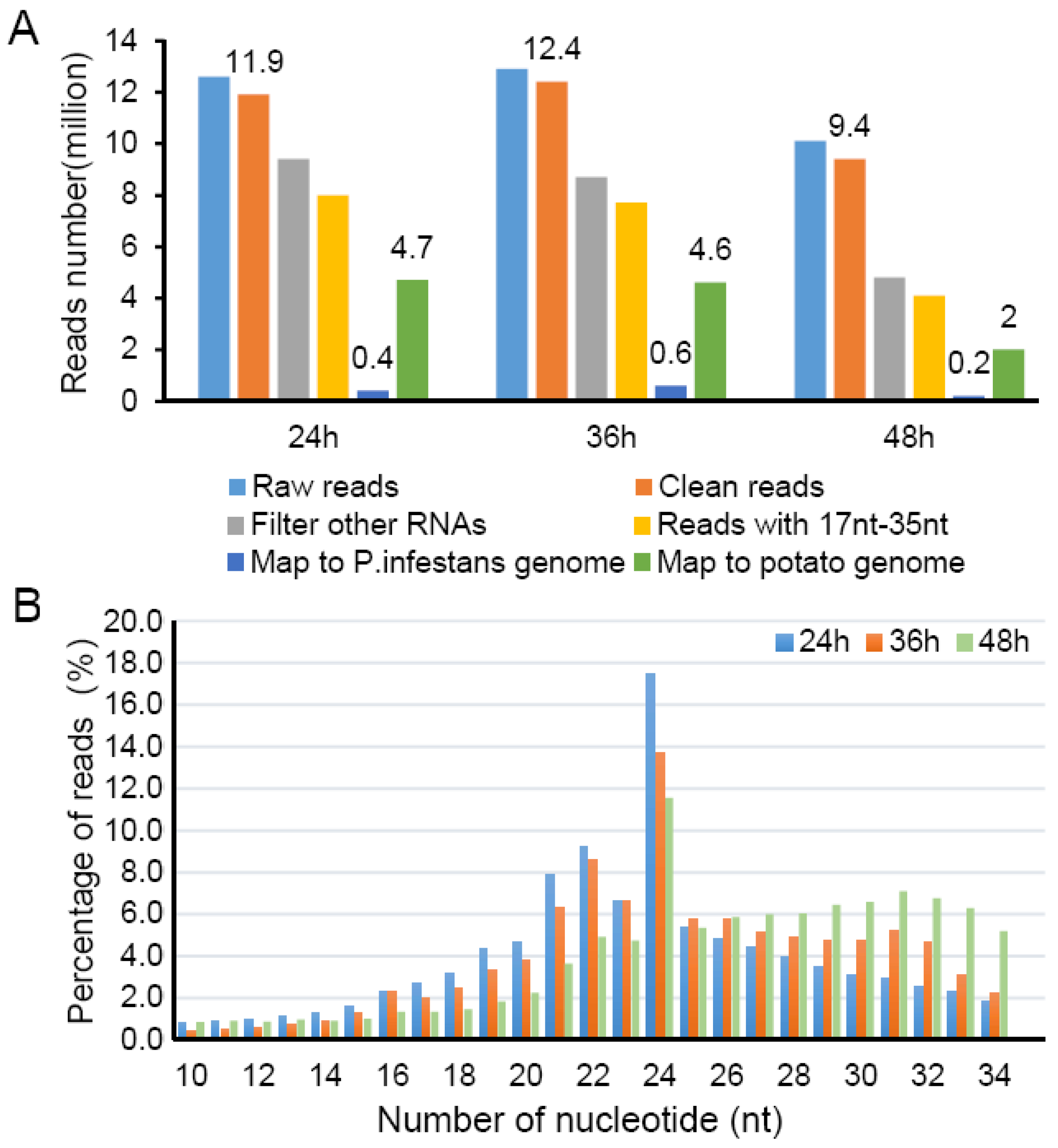
2.1. sRNA Analysis during Potato-P. infestans Interaction
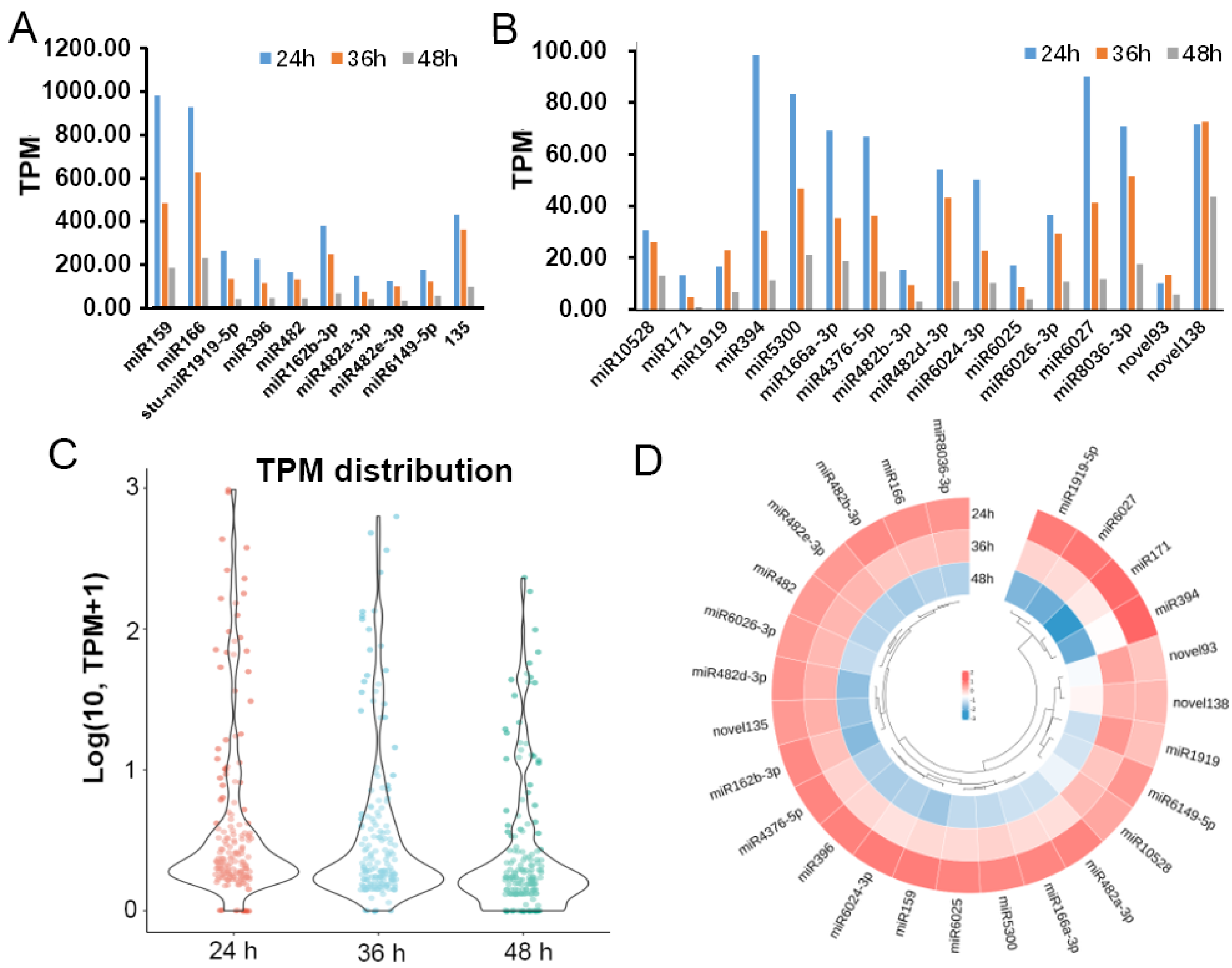
2.2. Prediction of miRNAs Expressed during Potato-P. infestans Interaction
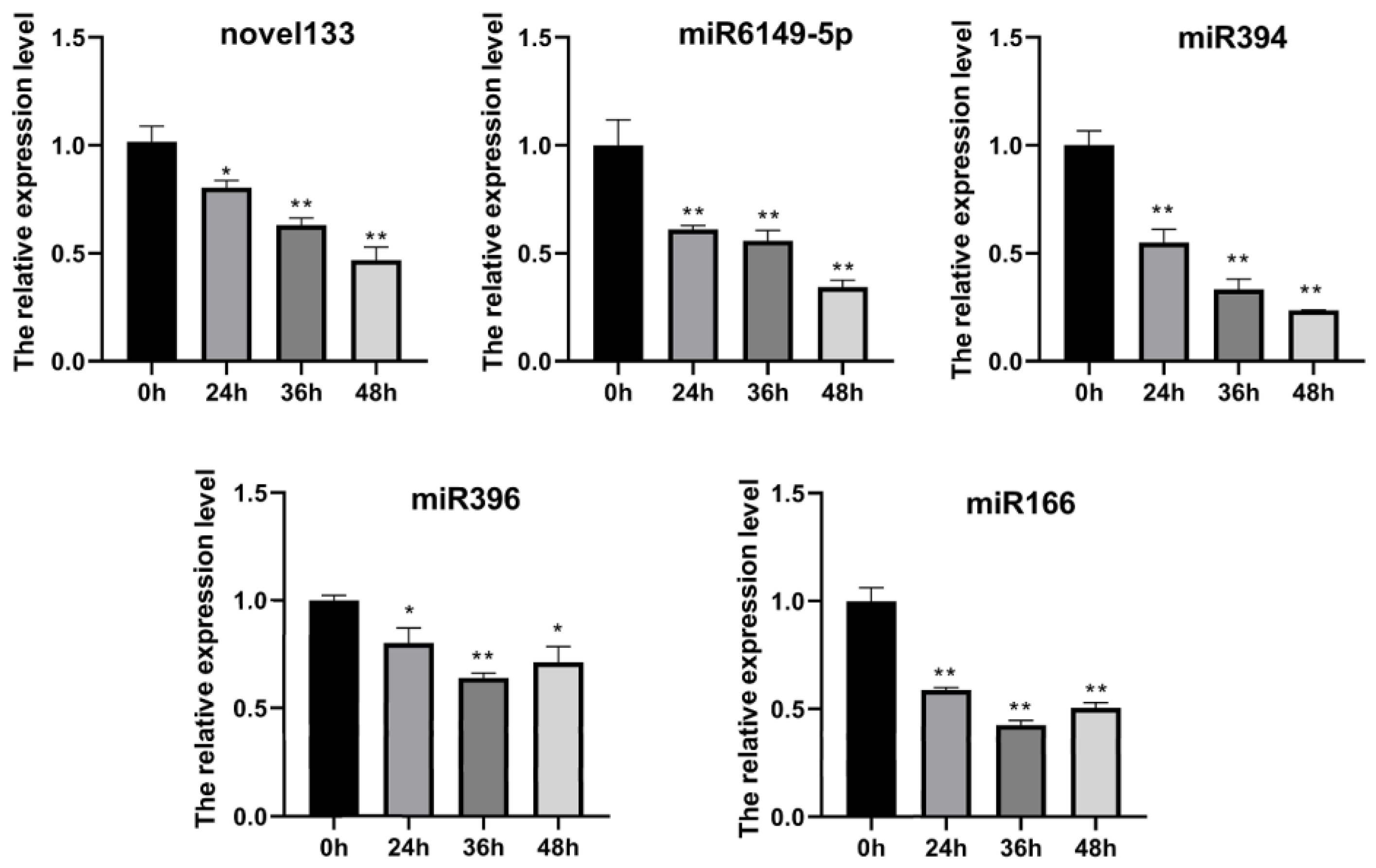
2.3. Most of Potato miRNAs Were Expressed in the Early Stage of Infection
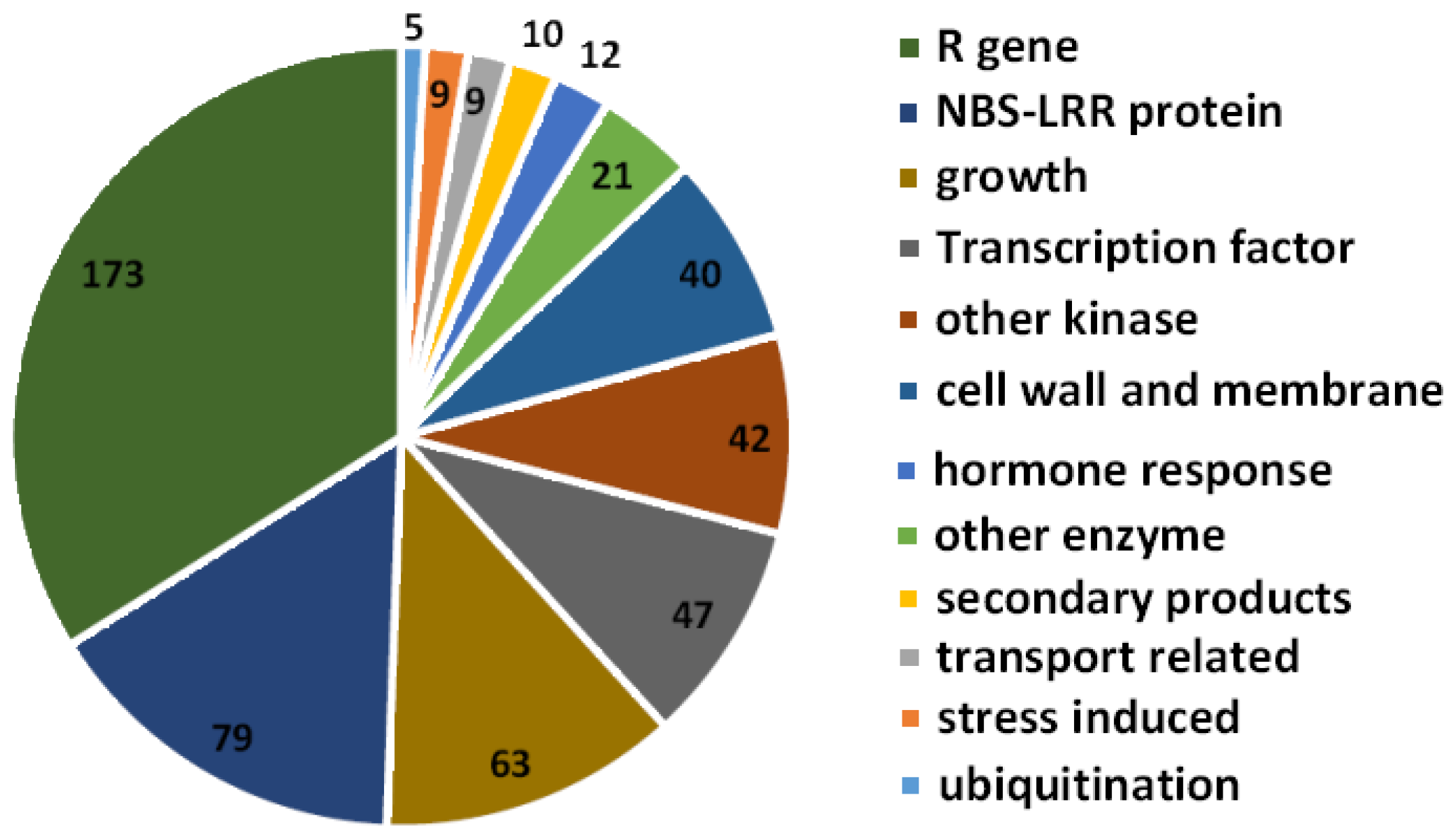
2.4. Targets Prediction of Potato-Derived miRNAs in Potato Genome
2.5. Targets Prediction of Potato-Derived miRNAs in P. infestans Genome
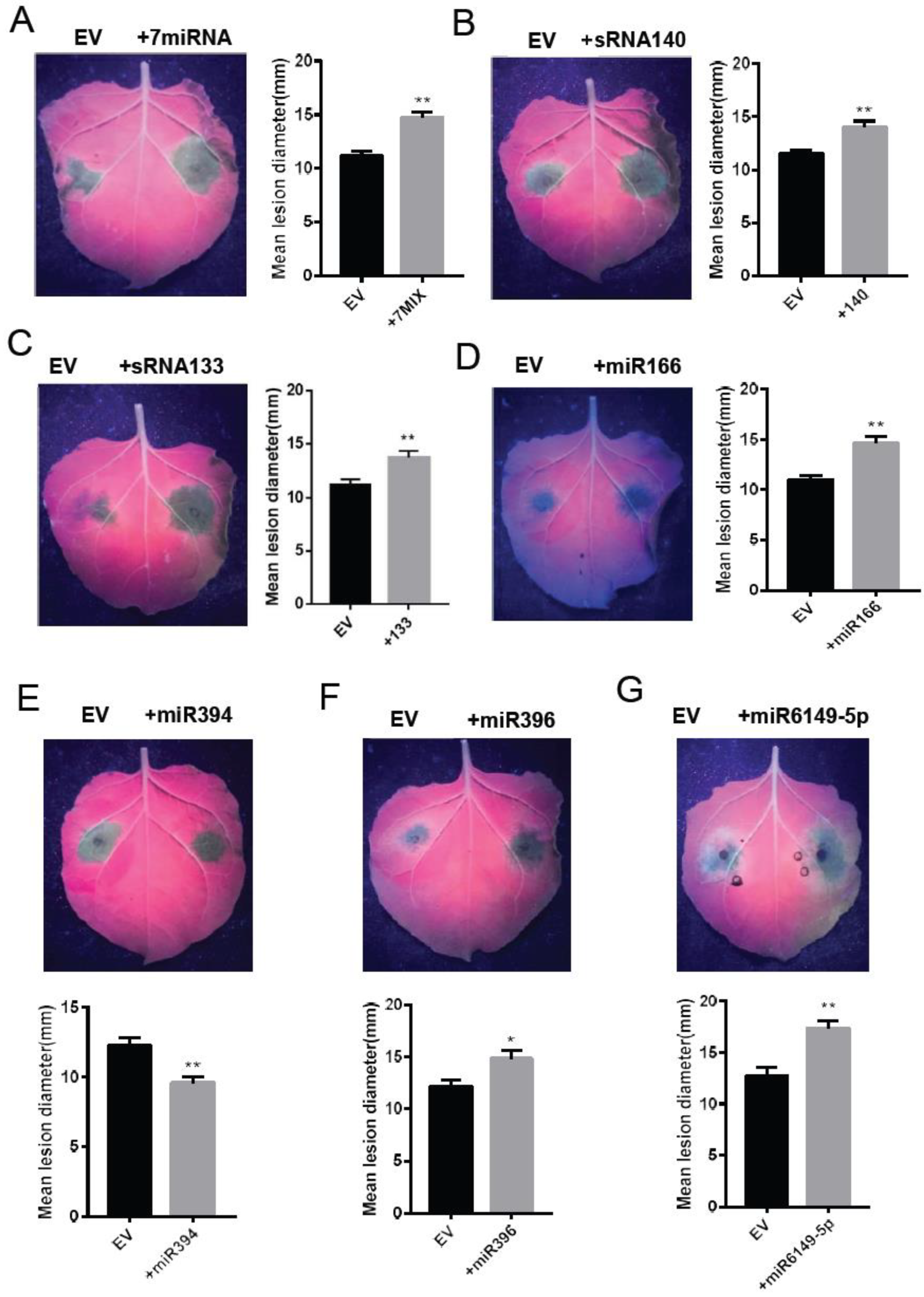
2.6. miRNAs Regulate Plant Resistance against P. infestans
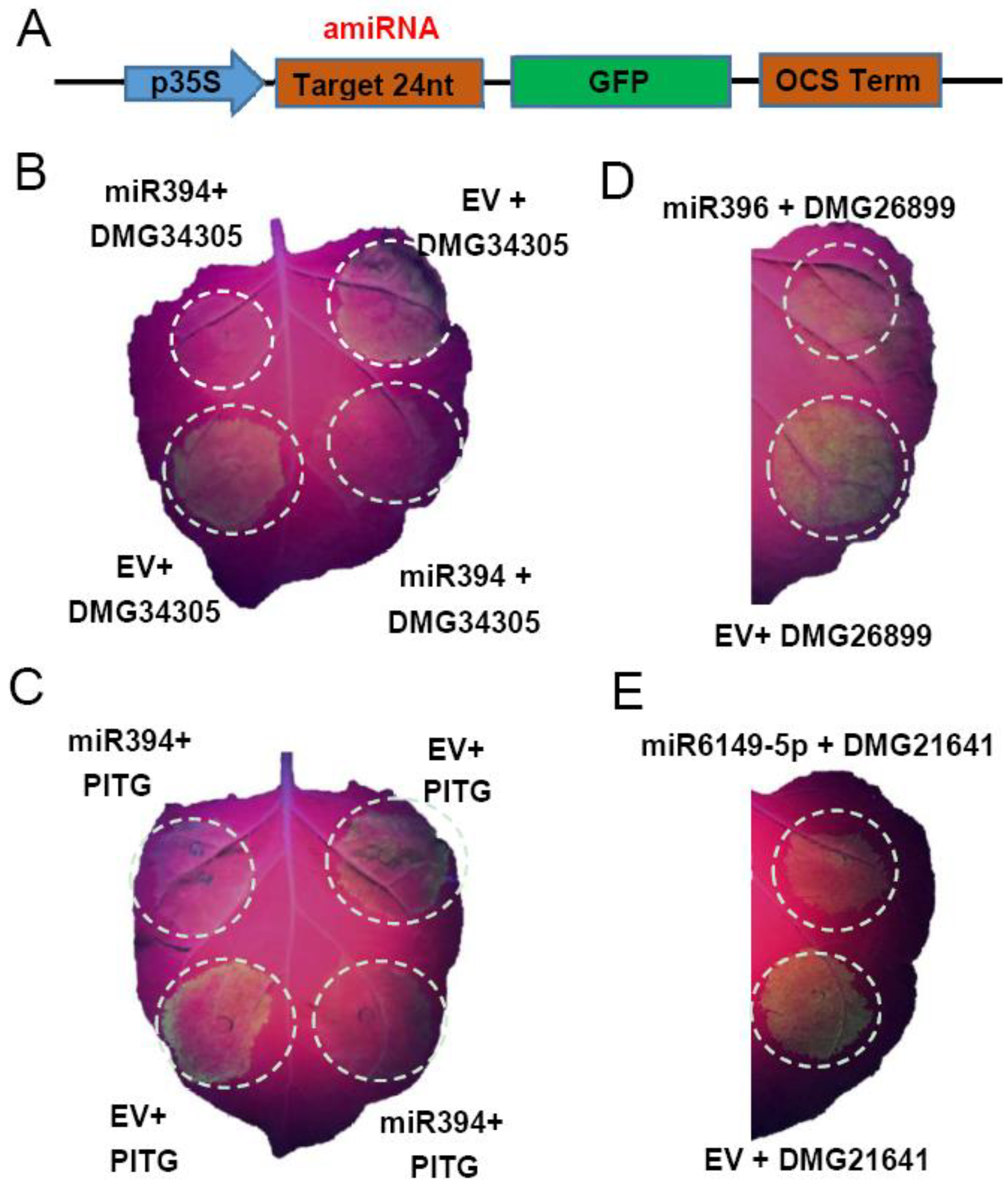
2.7. Confirmation of the Degradation Ability of miRNA by amiRNAs Strategy
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. P. infestans Inoculation and Infection Assay
4.3. sRNA-seq and Bioinformatics Analysis
4.4. Plasmid Constructs
4.5. Agrobacterium-Mediated Transient Gene Expression Assays
4.6. RNA Extraction and Quantitative Real-Time RT-PCR Analysis
4.7. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Finnegan, E.J.; Matzke, M.A. The small RNA world. J. Cell Sci. 2003, 116, 4689–4693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondhare, K.R.; Malankar, N.N.; Devani, R.S.; Banerjee, A.K. Genome-wide transcriptome analysis reveals small RNA profiles involved in early stages of stolon-to-tuber transitions in potato under photoperiodic conditions. BMC Plant Biol. 2018, 18, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Luo, Y.; Gong, X.; Zeng, W.; Li, S. Computational identification of 48 potato microRNAs and their targets. Comput. Biol. Chem. 2009, 33, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Bonar, N.; Liney, M.; Zhang, R.; Austin, C.; Dessoly, J.; Davidson, D.; Stephens, J.; McDougall, G.; Taylor, M.; Bryan, G.J.; et al. Potato miR828 Is associated with purple tuber skin and flesh color. Front. Plant Sci. 2018, 9, 1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, E.W.; Shin, S.J.; Yu, B.K.; Byun, M.O.; Kwon, H.B. miR171 family members are involved in drought response in Solanum tuberosum. J. Plant Biol. 2011, 54, 43–48. [Google Scholar] [CrossRef]
- Chen, L.; Luan, Y.; Zhai, J. Sp-miR396a-5p acts as a stress-responsive genes regulator by conferring tolerance to abiotic stresses and susceptibility to Phytophthora nicotianae infection in transgenic tobacco. Plant Cell Rep. 2015, 34, 2013–2025. [Google Scholar] [CrossRef]
- Islam, W.; Noman, A.; Qasim, M.; Wang, L. Plant Responses to Pathogen Attack: Small RNAs in Focus. Int. J. Mol. Sci. 2018, 19, 515. [Google Scholar] [CrossRef] [Green Version]
- Kuan, T.; Zhai, Y.; Ma, W. Small RNAs regulate plant responses to filamentous pathogens. Semin. Cell Dev. Biol. 2016, 56, 190–200. [Google Scholar] [CrossRef] [Green Version]
- Luan, Y.; Cui, J.; Zhai, J.; Li, J.; Han, L.; Meng, J. High-throughput sequencing reveals differential expression of miRNAs in tomato inoculated with Phytophthora infestans. Planta 2015, 241, 1405–1416. [Google Scholar] [CrossRef]
- Li, Y.; Hu, X.; Chen, J.; Wang, W.; Xiong, X.; He, C. Integrated mRNA and microRNA transcriptome analysis reveals miRNA regulation in response to PVA in potato. Sci. Rep. 2017, 7, 16925. [Google Scholar] [CrossRef]
- Bhavani, N.; Harpreet, S.K.; Prajakta, G.; Nilam, M.; Aarthy, T.; Sundaresha, S.; Hirekodathakallu, V.T.; Swarup, K.C.; Anjan, K.B. miRNA160 is associated with local defense and systemic acquired resistance against infection in potato. J. Exp. Bot. 2018, 69, 2023–2036. [Google Scholar]
- Salvador-Guirao, R.; Baldrich, P.; Weigel, D.; Rubio-Somoza, I.; San Segundo, B. The MicroRNA miR773 is involved in the Arabidopsis immune response to fungal pathogens. Mol. Plant Microbe. Interact. 2018, 31, 249–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Meng, J.; He, X.L.; Zhang, M.; Luan, Y.S. Solanum lycopersicum microRNA1916 targets multiple target genes and negatively regulates the immune response in tomato. Plant Cell Environ. 2019, 42, 1393–1407. [Google Scholar] [CrossRef] [PubMed]
- Rhoades, M.W.; Reinhart, B.J.; Lim, L.P.; Burge, C.B.; Bartel, B.; Bartel, D.P. Prediction of plant microRNA targets. Cell 2002, 110, 513–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, Y.; Cui, J.; Li, J.; Jiang, N.; Liu, P.; Meng, J. Effective enhancement of resistance to Phytophthora infestans by overexpression of miR172a and b in Solanum lycopersicum. Planta 2018, 247, 127–138. [Google Scholar] [CrossRef]
- Li, F.; Pignatta, D.; Bendix, C.; Brunkard, J.O.; Cohn, M.M.; Tung, J.; Sun, H.; Kumar, P.; Baker, B. MicroRNA regulation of plant innate immune receptors. Proc. Natl. Acad. Sci. USA 2012, 109, 1790–1795. [Google Scholar] [CrossRef] [Green Version]
- Jiang, N.; Meng, J.; Cui, J.; Sun, G.X.; Luan, Y.S. Function identification of miR482b, a negative regulator during tomato resistance to Phytophthora infestans. Hortic. Res. 2018, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, S.; Park, G.; Atamian, H.S.; Han, C.S.; Stajich, J.E.; Kaloshian, I.; Borkovich, K.A. MicroRNAs suppress NB domain genes in tomato that confer resistance to Fusarium oxysporum. PLoS Pathog. 2014, 10, e1004464. [Google Scholar] [CrossRef]
- Wang, C.; He, X.; Wang, X.; Zhang, S.; Guo, X. ghr-miR5272a-mediated regulation of GhMKK6 gene transcription contributes to the immune response in cotton. J. Exp. Bot. 2017, 68, 5895–5906. [Google Scholar] [CrossRef] [Green Version]
- de Vries, S.; Kukuk, A.; von Dahlen, J.K.; Schnake, A.; Kloesges, T.; Rose, L.E. Expression profiling across wild and cultivated tomatoes supports the relevance of early miR482/2118 suppression for Phytophthora resistance. Proc. Biol. Sci. 2018, 285, 20172560. [Google Scholar] [CrossRef] [Green Version]
- Eckardt, N.A. A microRNA cascade in plant defense. Plant Cell. 2012, 24, 840. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of microRNA biogenesis, mechanisms of actions, and circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Liang, H.; Zhang, J.; Zen, K.; Zhang, C.Y. Secreted microRNAs: A new form of intercellular communication. Trends Cell Biol. 2012, 22, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.Y.; Wang, H.; Hu, P.; Hamby, R.; Jin, H. Small RNAs—Big players in Plant-Microbe Interactions. Cell Host Microbe 2019, 26, 173–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timmons, L.; Fire, A. Specific interference by ingested dsRNA. Nature 1998, 395, 854. [Google Scholar] [CrossRef]
- Whangbo, J.S.; Hunter, C.P. Environmental RNA interference. Trends Genet. 2008, 24, 297–305. [Google Scholar] [CrossRef]
- Wang, W.; Galili, G. Tuning the Orchestra: miRNAs in plant immunity. Trends Plant Sci. 2019, 24, 189–191. [Google Scholar] [CrossRef]
- Cai, Q.; Qiao, L.; Wang, M.; He, B.; Lin, F.M.; Palmquist, J.; Huang, S.D.; Jin, H. Plants send small RNAs in extracellular vesicles to fungal pathogen to silence virulence genes. Science 2018, 360, 1126–1129. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Persson Hodén, K.; Liao, Z.; Åsman, A.; Dixelius, C. Phytophthora infestans Ago1-associated miRNA promotes potato late blight disease. New Phytol. 2022, 233, 443–457. [Google Scholar] [CrossRef]
- Weiberg, A.; Wang, M.; Lin, F.M.; Zhao, H.; Zhang, Z.; Kaloshian, I.; Huang, H.D.; Jin, H. Fungal small RNAs suppress plant immunity by hijacking host RNA interference pathways. Science 2013, 342, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Feng, H.; Lu, X.; Wang, C.; Yang, W.; Li, F. An asymmetric bulge enhances artificial microRNA-mediated virus resistance. Plant Biotechnol. J. 2020, 18, 608–610. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Lu, W.; Zhong, C.; Zhou, R.; Xu, J.; Liu, W.; Gou, X.; Wang, Q.; Yin, J.; Xu, C.; et al. The 25-26 nt Small RNAs in Phytophthora parasitica are associated with efficient silencing of homologous endogenous genes. Front. Microbiol. 2017, 8, 773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Åsman, A.K.; Vetukuri, R.R.; Jahan, S.N.; Fogelqvist, J.; Corcoran, P.; Avrova, A.O.; Whisson, S.C.; Dixelius, C. Fragmentation of tRNA in Phytophthora infestans asexual life cycle stages and during host plant infection. BMC Microbiol. 2014, 14, 308. [Google Scholar]
- Sun, T.; Zhou, Q.; Zhou, Z.; Song, Y.; Li, Y.; Wang, H.B.; Liu, B. SQUINT positively regulates resistance to the pathogen Botrytis cinerea via miR156-SPL9 module in Arabidopsis. Plant Cell Physiol. 2022, 63, 1414–1432. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Zhang, Q.; Zhao, Y.Q.; Yang, J.; He, H.B.; Jia, G.X. The lre-miR159a-LrGAMYB pathway mediates resistance to grey mould infection in Lilium regale. Mol. Plant Pathol. 2020, 21, 749–760. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Zhang, Q.; Cui, R.; Xu, X.; Zhu, F.; Cheng, Q.; Li, X. High-throughput sequencing reveals the regulatory networks of transcriptome and small RNAs during the defense against Marssonina brunnea in poplar. Front. Plant Sci. 2021, 12, 719549. [Google Scholar] [CrossRef]
- Zhang, Y.; Waseem, M.; Zeng, Z.; Xu, J.; Chen, C.; Liu, Y.; Zhai, J.; Xia, R. MicroRNA482/2118, a miRNA superfamily essential for both disease resistance and plant development. New Phytol. 2022, 233, 2047–2057. [Google Scholar] [CrossRef]
- Dey, S.; Sarkar, A.; Chowdhury, S.; Singh, R.; Mukherjee, A.; Ghosh, Z.; Kundu, P. Heightened miR6024-NLR interactions facilitate necrotrophic pathogenesis in tomato. Plant Mol. Biol. 2022, 109, 717–739. [Google Scholar] [CrossRef]
- Deng, Y.; Liu, M.; Li, X.; Li, F. microRNA-mediated R gene regulation: Molecular scabbards for double-edged swords. Sci. China Life Sci. 2018, 61, 138–147. [Google Scholar] [CrossRef]
- Wang, J.; Long, X.; Chern, M.; Chen, X. Understanding the molecular mechanisms of trade-offs between plant growth and immunity. Sci. China Life Sci. 2021, 64, 234–241. [Google Scholar] [CrossRef]
- Cui, C.; Wang, J.J.; Zhao, J.H.; Fang, Y.Y.; He, X.F.; Guo, H.S.; Duan, C.G. A brassica miRNA regulates plant growth and immunity through distinct modes of action. Mol. Plant 2020, 13, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhao, Y.L.; Zhao, J.H.; Wang, S.; Jin, Y.; Chen, Z.Q.; Fang, Y.Y.; Hua, C.L.; Ding, S.W.; Guo, H.S. Cotton plants export microRNAs to inhibit virulence gene expression in a fungal pathogen. Nat. Plants 2016, 2, 16153. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Cai, Q.; Qiao, L.; Huang, C.Y.; Wang, S.; Miao, W.; Ha, T.; Wang, Y.; Jin, H. RNA-binding proteins contribute to small RNA loading in plant extracellular vesicles. Nat. Plants 2021, 7, 342–352. [Google Scholar] [CrossRef]
- Wang, M.; Weiberg, A.; Dellota, E., Jr.; Yamane, D.; Jin, H. Botrytis small RNA Bc-siR37 suppresses plant defense genes by cross-kingdom RNAi. RNA Biol. 2017, 14, 421–428. [Google Scholar] [PubMed] [Green Version]
- Shahid, S.; Kim, G.; Johnson, N.R.; Wafula, E.; Wang, F.; Coruh, C.; Bernal-Galeano, V.; Phifer, T.; dePamphilis, C.W.; Westwood, J.H.; et al. MicroRNAs from the parasitic plant Cuscuta campestris target host messenger RNAs. Nature 2018, 553, 82–85. [Google Scholar] [CrossRef]
- Song, J.B.; Gao, S.; Sun, D.; Li, H.; Shu, X.X.; Yang, Z.M. miR394 and LCR are involved in Arabidopsis salt and drought stress responses in an abscisic acid-dependent manner. BMC Plant Biol. 2013, 13, 210. [Google Scholar] [CrossRef] [Green Version]
- Qu, L.; Lin, L.B.; Xue, H.W. Rice miR394 suppresses leaf inclination through targeting an F-box gene, LEAF INCLINATION 4. J. Integr. Plant Biol. 2019, 61, 406–416. [Google Scholar] [CrossRef]
- Song, J.B.; Shu, X.X.; Shen, Q.; Li, B.W.; Song, J.; Yang, Z.M. Altered fruit and seed development of transgenic rapeseed (Brassica napus) over-expressing MicroRNA394. PLoS ONE 2015, 10, e0125427. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Song, L.; Wang, Y.; Jin, W.; Tong, F.; Wu, F. miR394 acts as a negative regulator of Arabidopsis resistance to B. cinerea Infection by targeting LCR. Front. Plant Sci. 2018, 9, 903. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.Y.; Hong, Y.H.; Liu, Y.R.; Cui, J.; Luan, Y.S. Function identification of miR394 in tomato resistance to Phytophthora infestans. Plant Cell Rep. 2021, 40, 1831–1844. [Google Scholar] [CrossRef]
- Chand, S.K.; Nanda, S.; Joshi, R.K. Regulation of miR394 in response to Fusarium oxysporum f. sp. cepae (FOC) infection in garlic (Allium sativum L). Front. Plant Sci. 2016, 7, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto-Suárez, M.; Baldrich, P.; Weigel, D.; Rubio-Somoza, I.; San Segundo, B. The Arabidopsis miR396 mediates pathogen-associated molecular pattern-triggered immune responses against fungal pathogens. Sci. Rep. 2017, 7, 44898. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Li, Z.; Yuan, N.; Hu, Q.; Zhou, M.; Zhao, J.; Li, D.; Luo, H. MiR396 is involved in plant response to vernalization and flower development in Agrostis stolonifera. Hortic. Res. 2020, 7, 173. [Google Scholar] [CrossRef] [PubMed]
- Gou, M.; Su, N.; Zheng, J.; Huai, J.; Wu, G.; Zhao, J.; He, J.; Tang, D.; Yang, S.; Wang, G. An F-box gene, CPR30, functions as a negative regulator of the defense response in Arabidopsis. Plant J. 2009, 60, 757–770. [Google Scholar] [CrossRef] [PubMed]
- Gou, M.; Shi, Z.; Zhu, Y.; Bao, Z.; Wang, G.; Hua, J. The F-box protein CPR1/CPR30 negatively regulates R protein SNC1 accumulation. Plant J. 2012, 69, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Rabuma, T.; Gupta, O.P.; Chhokar, V. Recent advances and potential applications of cross-kingdom movement of miRNAs in modulating plant’s disease response. RNA Biol. 2022, 19, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Qi, Y.; Mu, Y.; Zhou, J.; Lu, W.; Tian, Z. Potato StLecRK-IV.1 negatively regulates late blight resistance by affecting the stability of a positive regulator StTET8. Hortic. Res. 2022, 9, uhac010. [Google Scholar] [CrossRef]
- Diambra, L.A. Genome sequence and analysis of the tuber crop potato. Nature 2011, 475, 189–195. [Google Scholar]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.J.; Ma, Y.K.; Chen, T.; Wang, M.; Wang, X.J. PsRobot: A web-based plant small RNA meta-analysis toolbox. Nucleic Acids Res. 2012, 40, W22–W28. [Google Scholar] [CrossRef]
- Yasir, M.; Motawaa, M.; Wang, Q.W.; Zhang, X.; Khalid, A.; Cai, X.K.; Li, F. Simple webserver-facilitated method to design and synthesize artificial miRNA gene and its application in engineering viral resistance. Plants 2022, 11, 2125. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Qi, Y.; Nie, J.; Guo, L.; Luo, M.; McLellan, H.; Boevink, P.C.; Birch, P.R.J.; Tian, Z. A Phytophthora effector promotes homodimerization of host transcription factor StKNOX3 to enhance susceptibility. J. Exp. Bot. 2022, 73, 6902–6915. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | miRNA | 24 h | 36 h | 48 h | No. | miRNA | 24 h | 36 h | 48 h |
|---|---|---|---|---|---|---|---|---|---|
| 1 | miR10528 | 29.89932 | 25.37756 | 12.92606 | 23 | miR482a-3p | 148.6614 | 74.19915 | 40.70108 |
| 2 | miR156 | 0.918694 | 0.805637 | 0.320481 | 24 | miR482b-3p | 15.11669 | 9.506514 | 3.311636 |
| 3 | miR159 | 980.3301 | 482.8181 | 185.3448 | 25 | miR482d-3p | 52.5326 | 41.97368 | 10.89635 |
| 4 | miR162b-3p | 378.669 | 248.7001 | 67.72831 | 26 | miR482e-3p | 124.1072 | 99.8184 | 33.00954 |
| 5 | miR162b-5p | 8.351764 | 4.511566 | 1.709232 | 27 | miR530 | 1.4198 | 0.241691 | 0 |
| 6 | miR166 | 927.2129 | 624.3685 | 230.5326 | 28 | miR5300 | 80.59453 | 45.43792 | 20.72443 |
| 7 | miR166a-3p | 67.06467 | 34.32013 | 18.37424 | 29 | miR6024-3p | 48.69079 | 22.23558 | 10.25539 |
| 8 | miR166b | 6.764929 | 3.06142 | 1.06827 | 30 | miR6025 | 16.70353 | 8.620314 | 4.166252 |
| 9 | miR167b-3p | 2.338494 | 1.852965 | 0.427308 | 31 | miR6026-3p | 35.66203 | 28.60011 | 10.78953 |
| 10 | miR167d-3p | 3.090153 | 3.464238 | 1.06827 | 32 | miR6027 | 87.02538 | 40.12071 | 11.64414 |
| 11 | miR171 | 13.19579 | 4.994948 | 1.281924 | 33 | miR6149-5p | 175.9717 | 121.9734 | 56.51147 |
| 12 | miR171b-3p | 2.422012 | 1.691837 | 0.640962 | 34 | miR7122-3p | 1.252765 | 0.322255 | 0.320481 |
| 13 | miR1919 | 16.28594 | 22.47727 | 6.836927 | 35 | miR8011a-3p | 0.835176 | 0.483382 | 0.106827 |
| 14 | miR319-3p | 2.338494 | 8.056368 | 11.75097 | 36 | miR8016 | 7.767141 | 1.611274 | 1.602405 |
| 15 | miR394 | 94.95956 | 29.64743 | 11.21683 | 37 | miR8033-5p | 1.586835 | 0.644509 | 0.320481 |
| 16 | miR396 | 226.4163 | 115.6089 | 47.21753 | 38 | miR8036-3p | 68.56798 | 50.03005 | 17.30597 |
| 17 | miR397b-3p | 0.918694 | 0.563946 | 0.320481 | 39 | miR8036-5p | 0.668141 | 0.161127 | 0.320481 |
| 18 | miR398 | 2.422012 | 2.578038 | 0.427308 | 40 | miR8044-3p | 1.002212 | 0.8862 | 0.106827 |
| 19 | miR398b-3p | 3.006635 | 5.317203 | 0.961443 | 41 | miR8051-5p | 0.584623 | 0.402818 | 0.213654 |
| 20 | miR4376-5p | 64.72617 | 35.28689 | 14.52847 | 42 | miR1919-5p | 264.5004 | 134.3802 | 41.12839 |
| 21 | miR477b-5p | 1.002212 | 0.725073 | 0.534135 | 43 | miR8039 | 1.586835 | 2.336347 | 1.495578 |
| 22 | miR482 | 164.6133 | 132.0439 | 44.86733 |
| GO ID | Description | NO. | Bg | p.Value | |
|---|---|---|---|---|---|
| CC | GO:0005669 | Transcription factor TFIID complex | 2 | 7 | 3.30 × 10−1 |
| GO:0030127 | COPII vesicle coat | 2 | 14 | 3.83 × 10−2 | |
| GO:0005886 | Plasma membrane | 96 | 2741 | 2.40 × 10−4 | |
| MF | GO:0003955 | NAD(P)H dehydrogenase (quinone) activity | 2 | 3 | 3.64 × 10−2 |
| GO:0015152 | Glucose-6-phosphate transmembrane transporter activity | 2 | 3 | 3.64 × 10−2 | |
| GO:0010542 | Nitrate efflux transmembrane transporter activity | 3 | 6 | 7.48 × 10−3 | |
| GO:0015562 | Efflux transmembrane transporter activity | 5 | 36 | 4.71 × 10−2 | |
| GO:0004872 | Receptor activity | 36 | 192 | 2.27 × 10−18 | |
| BP | GO:0048263 | Determination of dorsal identity | 2 | 2 | 3.91 × 10−2 |
| GO:0010072 | Primary shoot apical meristem specification | 5 | 20 | 1.05 × 10−2 | |
| GO:0009814 | Defense response, incompatible interaction | 20 | 171 | 6.20 × 10−6 | |
| GO:0009626 | Plant-type hypersensitive response | 63 | 173 | 1.08 × 10−50 | |
| GO:0045087 | Innate immune response | 71 | 412 | 2.68 × 10−33 | |
| GO:0007154 | Cell communication | 80 | 1757 | 6.63 × 10−4 | |
| GO:0012501 | Programmed cell death | 171 | 410 | 1.13 × 10−156 | |
| GO:0008219 | Cell death | 173 | 475 | 8.70 × 10−147 | |
| GO:0006952 | Defense response | 240 | 1529 | 1.09 × 10−116 |
| Potato miRNA | P. infestans Gene | Annotation |
|---|---|---|
| novel 72 | PITG_15260 | Conserved hypothetical protein |
| PITG_15263 | Transmembrane protein | |
| novel 133 | PITG_05949 | Conserved hypothetical protein |
| PITG_07309 | Flagellar radial spoke protein | |
| PITG_09123 | Conserved hypothetical protein | |
| PITG_12774 | Conserved hypothetical protein | |
| novel 140 | PITG_08205 | Methionyl-trna formyltransferase |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, M.; Sun, X.; Xu, M.; Tian, Z. Identification of miRNAs Involving Potato-Phytophthora infestans Interaction. Plants 2023, 12, 461. https://doi.org/10.3390/plants12030461
Luo M, Sun X, Xu M, Tian Z. Identification of miRNAs Involving Potato-Phytophthora infestans Interaction. Plants. 2023; 12(3):461. https://doi.org/10.3390/plants12030461
Chicago/Turabian StyleLuo, Ming, Xinyuan Sun, Meng Xu, and Zhendong Tian. 2023. "Identification of miRNAs Involving Potato-Phytophthora infestans Interaction" Plants 12, no. 3: 461. https://doi.org/10.3390/plants12030461