

Small Talk: On the Possible Role of Trans-Kingdom Small RNAs during Plant–Virus–Vector Tritrophic Communication

Abstract
:1. Introduction
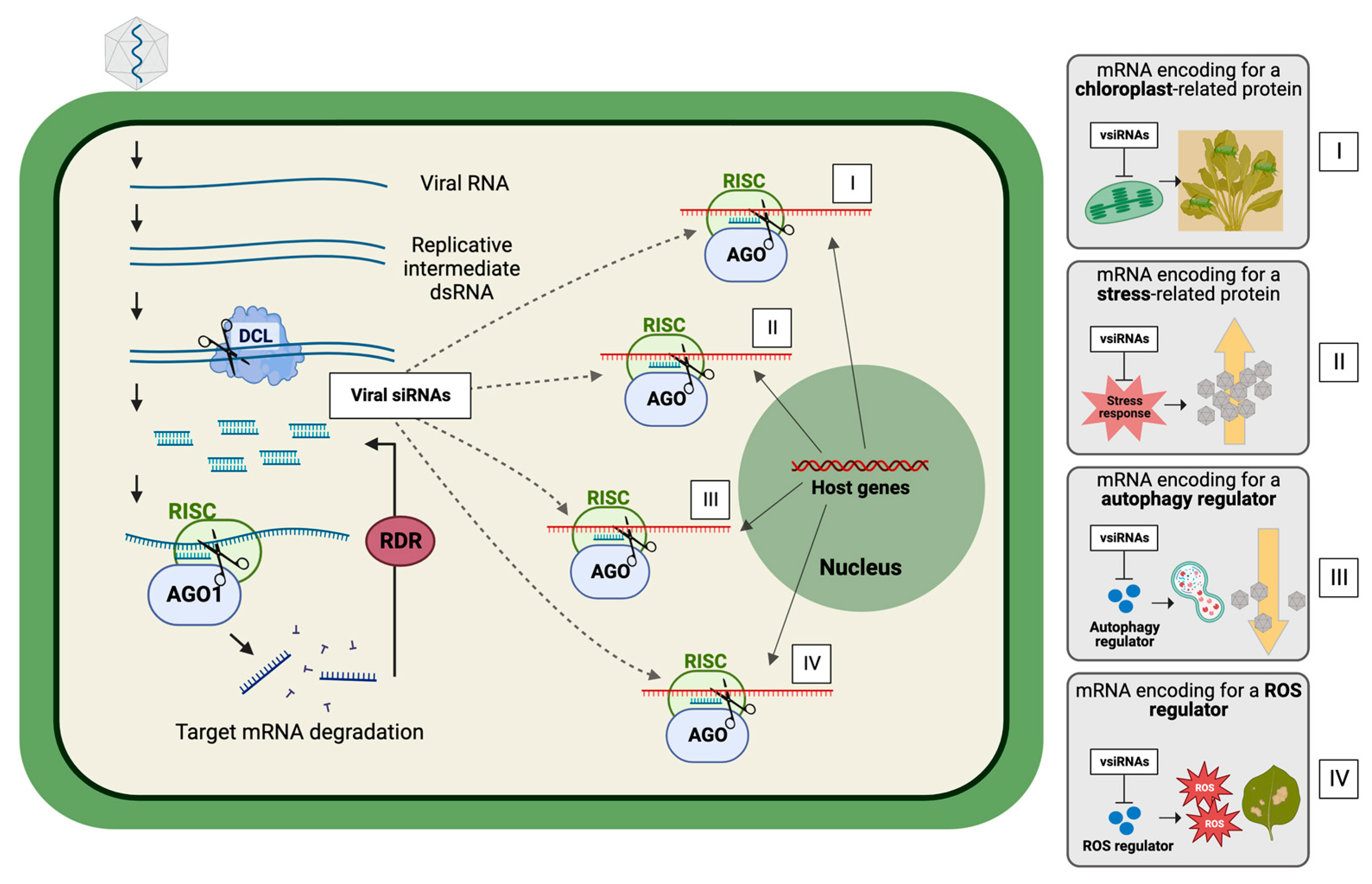
2. Virus-Derived Small RNAs as Modulators of Virus–Plant Interactions
Virus-Derived sRNAs Regulating Plant Host Genes Expression
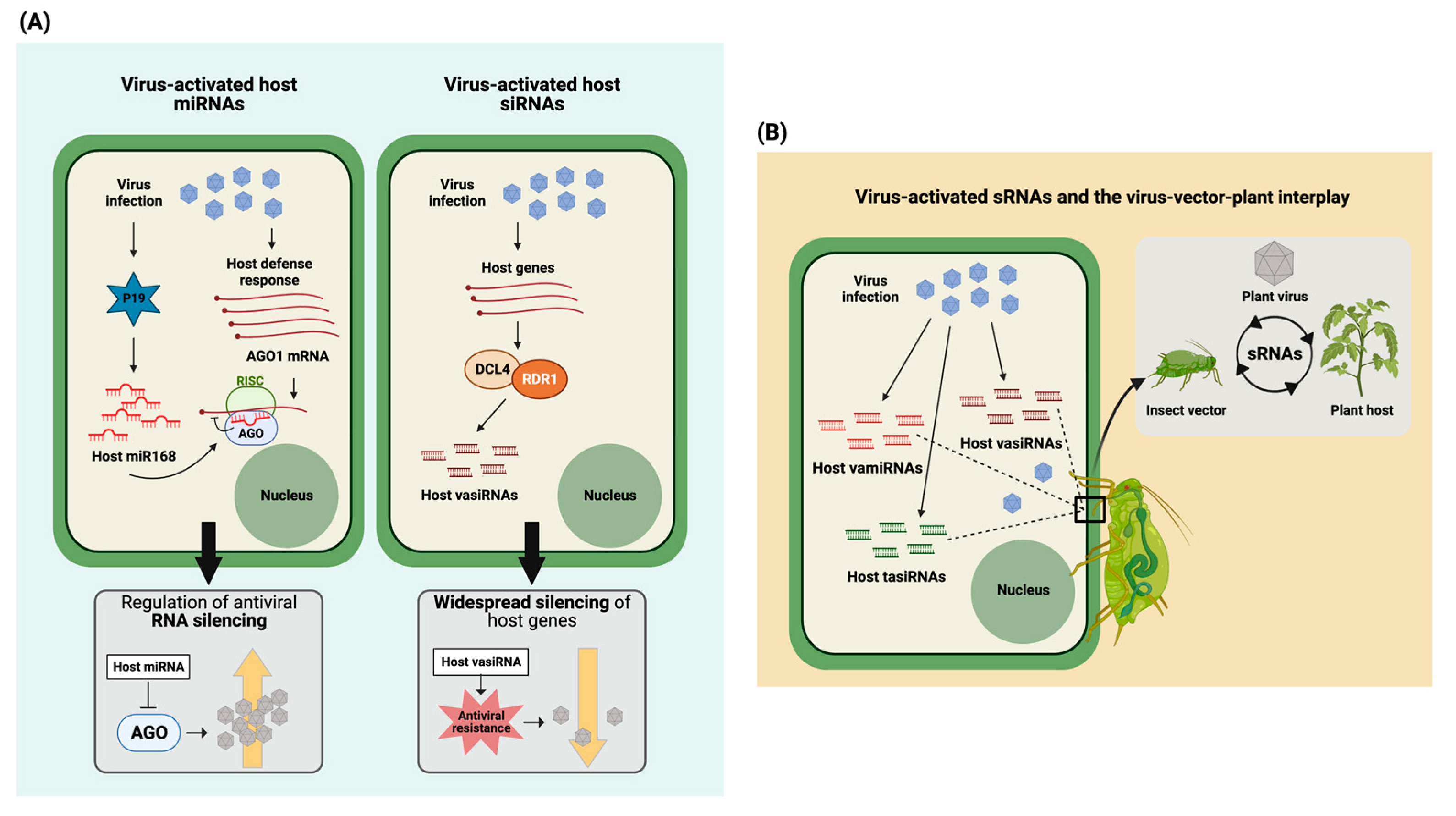
3. Virus-Activated Plant Endogenous Small RNAs as Modulators of Virus–Plant Interactions
4. Crosstalk between Virus and Insect Transmission Vector: Antiviral Defense Meets Herbivore Defense in Host Plant
5. Viruses as Modulators of sRNA-Based Communication between Vectors and Host Plant, with Implications on Virus Spread
6. Perspectives
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baulcombe, D. RNA silencing in plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef]
- Borges, F.; Martienssen, R.A. The expanding world of small RNAs in plants. Nat. Rev. Mol. Cell Biol. 2015, 12, 727–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, J.; Yang, M.; Lu, L.; Zhang, X. Diverse Functions of Small RNAs in Different Plant-Pathogen Communications. Front. Microbiol. 2016, 7, 1552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Z.; Ma, L.; Zhang, P.; Zhu, H. Small RNAs Participate in Plant-Virus Interaction and Their Application in Plant Viral Defense. Int. J. Mol. Sci. 2022, 23, 696. [Google Scholar] [CrossRef] [PubMed]
- Hudzik, C.; Hou, Y.; Ma, W.; Axtell, M.J. Exchange of Small Regulatory RNAs between Plants and Their Pests. Plant Physiol. 2020, 182, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Ding, S.W.; Voinnet, O. Antiviral immunity directed by small RNAs. Cell 2007, 130, 413–426. [Google Scholar] [CrossRef] [Green Version]
- Katiyar-Agarwal, S.; Jin, H. Role of small RNAs in host-microbe interactions. Annu. Rev. Phytopathol. 2010, 48, 225–246. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.Y.; Wang, H.; Hu, P.; Hamby, R.; Jin, H. Small RNAs—Big Players in Plant-Microbe Interactions. Cell Host Microbe 2019, 26, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Guleria, P.; Mahajan, M.; Bhardwaj, J.; Yadav, S.K. Plant small RNAs: Biogenesis, mode of action and their roles in abiotic stresses. Genom. Proteom. Bioinform. 2011, 9, 183–199. [Google Scholar] [CrossRef] [Green Version]
- Voinnet, O. Fly antiviral RNA silencing and miRNA biogenesis claim ARS2. Cell Host Microbe 2009, 6, 99–101. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, S.V.; Yogindran, S.; Gnanasekaran, P.; Chakraborty, S.; Winter, S.; Pappu, H.R. Virus and Viroid-Derived Small RNAs as Modulators of Host Gene Expression: Molecular Insights Into Pathogenesis. Front. Microbiol. 2021, 11, 614231. [Google Scholar] [CrossRef]
- Tang, Y.; Yan, X.; Gu, C.; Yuan, X. Biogenesis, Trafficking, and Function of Small RNAs in Plants. Front. Plant Sci. 2022, 13, 825477. [Google Scholar] [CrossRef]
- Annacondia, M.L.; Martinez, G. Reprogramming of RNA silencing triggered by cucumber mosaic virus infection in Arabidopsis. Genome Biol. 2021, 22, 340. [Google Scholar] [CrossRef]
- Díaz-Pendón, J.A.; Ding, S.W. Direct and indirect roles of viral suppressors of RNA silencing in pathogenesis. Annu. Rev. Phytopathol. 2008, 46, 303–326. [Google Scholar] [CrossRef] [PubMed]
- Csorba, T.; Kontra, L.; Burgyán, J. Viral silencing suppressors: Tools forged to fine-tune host-pathogen coexistence. Virology 2015, 479–480, 85–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daròs, J.A. Viral suppressors: Combatting RNA silencing. Nat. Plants 2017, 3, 17098. [Google Scholar] [CrossRef]
- Zhou, C.J.; Zhang, X.Y.; Liu, S.Y.; Wang, Y.; Li, D.W.; Yu, J.L.; Han, C.G. Synergistic infection of BrYV and PEMV 2 increases the accumulations of both BrYV and BrYV-derived siRNAs in Nicotiana benthamiana. Sci. Rep. 2017, 7, 45132. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Zhao, Y.L.; Zhao, J.H.; Wang, S.; Jin, Y.; Chen, Z.Q.; Fang, Y.Y.; Hua, C.L.; Ding, S.W.; Guo, H.S. Cotton plants export microRNAs to inhibit virulence gene expression in a fungal pathogen. Nat. Plants 2016, 2, 16153. [Google Scholar] [CrossRef] [PubMed]
- Gualtieri, C.; Leonetti, P.; Macovei, A. Plant miRNA Cross-Kingdom Transfer Targeting Parasitic and Mutualistic Organisms as a Tool to Advance Modern Agriculture. Front. Plant Sci. 2020, 11, 930. [Google Scholar] [CrossRef]
- Hou, Y.; Ma, W. Natural Host-Induced Gene Silencing Offers New Opportunities to Engineer Disease Resistance. Trends Microbiol. 2020, 28, 109–117. [Google Scholar] [CrossRef]
- Shahid, S.; Kim, G.; Johnson, N.R.; Wafula, E.; Wang, F.; Coruh, C.; Bernal-Galeano, V.; Phifer, T.; de Pamphilis, C.W.; Westwood, J.H.; et al. MicroRNAs from the parasitic plant Cuscuta campestris target host messenger RNAs. Nature 2018, 553, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Maule, A.; Leh, V.; Lederer, C. The dialogue between viruses and hosts in compatible interactions. Curr. Opin. Plant Biol. 2002, 5, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Whitham, S.A.; Quan, S.; Chang, H.S.; Cooper, B.; Estes, B.; Zhu, T.; Wang, X.; Hou, Y.M. Diverse RNA viruses elicit the expression of common sets of genes in susceptible Arabidopsis thaliana plants. Plant J. 2003, 33, 271–283. [Google Scholar] [CrossRef] [PubMed]
- Aparicio, F.; Thomas, C.L.; Lederer, C.; Niu, Y.; Wang, D.; Maule, A.J. Virus induction of heat shock protein 70 reflects a general response to protein accumulation in the plant cytosol. Plant Physiol. 2005, 138, 529–536. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Guo, R.; Jie, F.; Nettleton, D.; Peng, J.; Carr, T.; Yeakley, J.M.; Fan, J.B.; Whitham, S.A. Spatial analysis of arabidopsis thaliana gene expression in response to Turnip mosaic virus infection. Mol. Plant-Microbe Interact. 2007, 20, 358–370. [Google Scholar] [CrossRef] [Green Version]
- Pitzalis, N.; Amari, K.; Graindorge, S.; Pflieger, D.; Donaire, L.; Wassenegger, M.; Llave, C.; Heinlein, M. Turnip mosaic virus in oilseed rape activates networks of sRNA-mediated interactions between viral and host genomes. Commun. Biol. 2020, 3, 702. [Google Scholar] [CrossRef] [PubMed]
- Bizabani, C.; Rogans, S.J.; Rey, M.E.C. Differential miRNA profiles in South African cassava mosaic virus-infected cassava landraces reveal clues to susceptibility and tolerance to cassava mosaic disease. Virus Res. 2021, 303, 198400. [Google Scholar] [CrossRef]
- Lopez-Gomollon, S.; Baulcombe, D.C. Roles of RNA silencing in viral and non-viral plant immunity and in the crosstalk between disease resistance systems. Nat. Rev. Mol. Cell Biol. 2022, 23, 645–662. [Google Scholar] [CrossRef]
- Hu, Q.; Hollunder, J.; Niehl, A.; Kørner, C.J.; Gereige, D.; Windels, D.; Arnold, A.; Kuiper, M.; Vazquez, F.; Poooggin, M.; et al. Specific Impact of Tobamovirus Infection on the Arabidopsis Small RNA Profile. PLoS ONE 2011, 6, e19549. [Google Scholar] [CrossRef] [Green Version]
- Cao, M.; Du, P.; Wang, X.; Yu, Y.Q.; Qiu, Y.H.; Li, W.; Gal-On, A.; Zhou, C.; Li, Y.; Ding, S.W. Virus infection triggers widespread silencing of host genes by a distinct class of endogenous siRNAs in Arabidopsis. Proc. Natl. Acad. Sci. USA 2014, 111, 14613–14618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Z.; Zhao, Z.; Jiao, Z.; Xu, T.; Wu, Y.; Zhou, T.; Fan, Z. Virus-Derived Small Interfering RNAs Affect the Accumulations of Viral and Host Transcripts in Maize. Viruses 2018, 10, 664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimura, H.; Pantaleo, V.; Ishihara, T.; Myojo, N.; Inaba, J.; Sueda, K.; Burgyán, J.; Masuta, C. A viral satellite RNA induces yellow symptoms on tobacco by targeting a gene involved in chlorophyll biosynthesis using the RNA silencing machinery. PLoS Pathog. 2011, 7, e1002021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, N.A.; Eamens, A.L.; Wang, M.B. Viral small interfering RNAs target host genes to mediate disease symptoms in plants. PLoS Pathog. 2011, 7, e1002022. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Liu, T.; Shen, D.; Wang, J.; Ling, X.; Hu, Z.; Chen, T.; Hu, J.; Huang, J.; Yu, W.; et al. Tomato yellow leaf curl virus intergenic siRNAs target a host long noncoding RNA to modulate disease symptoms. PLoS Pathog. 2019, 15, e1007534. [Google Scholar] [CrossRef]
- Guo, S.; Wong, S.M. Small RNA derived from Tobacco mosaic virus targets a host C2-domain abscisic acid-related (CAR) 7-like protein gene. Phytopathol. Res. 2020, 2, 15. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, T.; Li, J.; Wu, N.; Wu, G.; Yang, J.; Chen, X.; He, L.; Chen, J. Chinese wheat mosaic virus-derived vsiRNA-20 can regulate virus infection in wheat through inhibition of vacuolar-(H+ )-PPase induced cell death. New Phytol. 2020, 226, 205–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, B.; Lin, L.; Wang, S.; Guo, Q.; Zhou, H.; Rong, L.; Li, J.; Peng, J.; Lu, Y.; Zheng, H.; et al. Identification and regulation of host genes related to Rice stripe virus symptom production. New Phytol. 2016, 209, 1106–1119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yin, Y.; Su, Y.; Jia, Z.; Jiang, L.; Lu, Y.; Zheng, H.; Peng, J.; Rao, S.; Wu, G.; et al. eIF4A, a target of siRNA derived from rice stripe virus, negatively regulates antiviral autophagy by interacting with ATG5 in Nicotiana benthamiana. PLoS Pathog. 2021, 17, e1009963. [Google Scholar] [CrossRef]
- Liu, P.; Zhang, X.; Zhang, F.; Xu, M.; Ye, Z.; Wang, K.; Liu, S.; Han, X.; Cheng, Y.; Zhong, K.; et al. A virus-derived siRNA activates plant immunity by interfering with ROS scavenging. Mol. Plant 2021, 14, 1088–1103. [Google Scholar] [CrossRef] [PubMed]
- Miozzi, L.; Gambino, G.; Burgyan, J.; Pantaleo, V. Genome-wide identification of viral and host transcripts targeted by viral siRNAs in Vitis vinifera. Mol. Plant Pathol. 2013, 14, 30–43. [Google Scholar] [CrossRef]
- Xu, D.; Zhou, G. Characteristics of siRNAs derived from Southern rice black-streaked dwarf virus in infected rice and their potential role in host gene regulation. Virol. J. 2017, 14, 27. [Google Scholar] [CrossRef] [Green Version]
- Maghuly, F.; Ramkat, R.C.; Laimer, M. Virus versus host plant microRNAs: Who determines the outcome of the interaction? PLoS ONE. 2014, 9, e98263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Niu, X.; Fan, M. Genome-wide identification of cucumber green mottle mosaic virus-responsive microRNAs in watermelon. Arch. Virol. 2017, 162, 2591–2602. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, C.; Anburaj, J.; Prabu, G. Identification and validation of sugarcane streak mosaic virus-encoded microRNAs and their targets in sugarcane. Plant Cell Rep. 2014, 33, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Moyo, L.; Ramesh, S.V.; Kappagantu, M.; Mitter, N.; Sathuvalli, V.; Pappu, H.R. The effects of potato virus Y-derived virus small interfering RNAs of three biologically distinct strains on potato (Solanum tuberosum) transcriptome. Virol. J. 2017, 14, 129. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zheng, H.; Zhang, C.; Han, K.; Wang, S.; Peng, J.; Lu, Y.; Zhao, J.; Xu, P.; Wu, X.; et al. Different Virus-Derived siRNAs Profiles between Leaves and Fruits in Cucumber Green Mottle Mosaic Virus-Infected Lagenaria siceraria Plants. Front. Microbiol. 2016, 7, 1797. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Xu, Z.; Zhao, W.; Liu, Q.; Li, Q.; Lu, L.; Liu, R.; Zhang, X.; Cui, F. Rice stripe virus-derived siRNAs play different regulatory roles in rice and in the insect vector Laodelphax striatellus. BMC Plant Biol. 2018, 18, 219. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, S.V.; Williams, S.; Kappagantu, M.; Mitter, N.; Pappu, H.R. Transcriptome-wide identification of host genes targeted by tomato spotted wilt virus-derived small interfering RNAs. Virus Res. 2017, 238, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Xia, Z.; Peng, J.; Li, Y.; Chen, L.; Li, S.; Zhou, T.; Fan, Z. Characterization of small interfering RNAs derived from Sugarcane mosaic virus in infected maize plants by deep sequencing. PLoS ONE 2014, 9, e97013. [Google Scholar] [CrossRef]
- Xia, Z.; Zhao, Z.; Chen, L.; Li, M.; Zhou, T.; Deng, C.; Zhou, Q.; Fan, Z. Synergistic infection of two viruses MCMV and SCMV increases the accumulations of both MCMV and MCMV-derived siRNAs in maize. Sci. Rep. 2016, 6, 20520. [Google Scholar] [CrossRef]
- Qiu, Y.; Zhang, Y.; Hu, F.; Zhu, S. Characterization of siRNAs derived from cucumber mosaic virus in infected tobacco plants. Arch. Virol. 2017, 162, 2077–2082. [Google Scholar] [CrossRef]
- Wang, H.; Jiao, X.; Kong, X.; Hamera, S.; Wu, Y.; Chen, X.; Fang, R.; Yan, Y. A Signaling Cascade from miR444 to RDR1 in Rice Antiviral RNA Silencing Pathway. Plant Physiol. 2016, 170, 2365–2377. [Google Scholar] [CrossRef] [Green Version]
- Pai, H.; Jean, W.H.; Lee, Y.S.; Chang, Y.A.; Lin, N.S. Genome-wide analysis of small RNAs from Odontoglossum ringspot virus and Cymbidium mosaic virus synergistically infecting Phalaenopsis. Mol. Plant Pathol. 2020, 21, 188–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, R.; Liu, P.; Wong, S.M. Identification of a plant viral RNA genome in the nucleus. PLoS ONE 2012, 7, e48736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sankaranarayanan, R.; Palani, S.N.; Kumar, A.; PunithaSelvakumar, A.S.; Tennyson, J. Prediction and experimental confirmation of banana bract mosaic virus encoding miRNAs and their targets. ExRNA 2020, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Shazia, I. Role of Viral and Host microRNAs in Plant Virus Interaction. Ph.D. Thesis, The University of Queensland, Brisbane, Australia, 2010. [Google Scholar]
- Mengistu, A.A.; Tenkegna, T.A. The role of miRNA in plant-virus interaction: A review. Mol. Biol. Rep. 2021, 48, 2853–2861. [Google Scholar] [CrossRef]
- Bazzini, A.A.; Almasia, N.I.; Manacorda, C.A.; Mongelli, V.C.; Conti, G.; Maroniche, G.A.; Rodriguez, M.C.; Distéfano, A.J.; Hopp, H.E.; del Vas, M.; et al. Virus infection elevates transcriptional activity of miR164a promoter in plants. BMC Plant Biol. 2009, 9, 152. [Google Scholar] [CrossRef] [Green Version]
- Bazzini, A.A.; Hopp, H.E.; Beachy, R.N.; Asurmendi, S. Infection and coaccumulation of tobacco mosaic virus proteins alter microRNA levels, correlating with symptom and plant development. Proc. Natl. Acad. Sci. USA 2007, 104, 12157–12162. [Google Scholar] [CrossRef] [Green Version]
- Katiyar-Agarwal, S.; Morgan, R.; Dahlbeck, D.; Borsani, O.; Villegas, A., Jr.; Zhu, J.K.; Staskawicz, B.J.; Jin, H. A pathogen-inducible endogenous siRNA in plant immunity. Proc. Natl. Acad. Sci. USA 2006, 103, 18002–18007. [Google Scholar] [CrossRef] [Green Version]
- Várallyay, E.; Válóczi, A.; Agyi, A.; Burgyán, J.; Havelda, Z. Plant virus-mediated induction of miR168 is associated with repression of ARGONAUTE1 accumulation. EMBO J. 2010, 29, 3507–3519. [Google Scholar] [CrossRef] [Green Version]
- Leonetti, P.; Pantaleo, V. Plant virus-derived siRNAs “turn on” broad-spectrum plant immunity in wheat. Mol. Plant 2021, 14, 1038–1040. [Google Scholar] [CrossRef]
- Fletcher, S.J.; Peters, J.R.; Olaya, C.; Persley, D.M.; Dietzgen, R.G.; Carroll, B.J.; Pappu, H.; Mitter, N. Tospoviruses Induce Small Interfering RNAs Targeting Viral Sequences and Endogenous Transcripts in Solanaceous Plants. Pathogens 2022, 11, 745. [Google Scholar] [CrossRef]
- Guo, Z.; Lu, J.; Wang, X.; Zhan, B.; Li, W.; Ding, S.W. Lipid flippases promote antiviral silencing and the biogenesis of viral and host siRNAs in Arabidopsis. Proc. Natl. Acad. Sci. USA 2017, 114, 1377–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Z.; Wang, X.B.; Wang, Y.; Li, W.X.; Gal-On, A.; Ding, S.W. Identification of a New Host Factor Required for Antiviral RNAi and Amplification of Viral siRNAs. Plant Physiol. 2018, 176, 1587–1597. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Mazur, M.J.; Tao, X.; Kormelink, R. Cellular RNA Hubs: Friends and Foes of Plant Viruses. Mol. Plant Microbe Interact. 2020, 33, 40–54. [Google Scholar] [CrossRef]
- Prigigallo, M.I.; Križnik, M.; Paola, D.; Catalano, D.; Gruden, K.; Finetti-Sialer, M.M.; Cillo, F. Potato Virus Y Infection Alters Small RNA Metabolism and Immune Response in Tomato. Viruses 2019, 11, 1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, H.; Jia, Z.; Gao, X.; Gui, M.; Li, Y.; Liu, Y. Analysis of the miRNA expression profile involved in the tomato spotted wilt orthotospovirus-pepper interaction. Virus Res. 2022, 312, 198710. [Google Scholar] [CrossRef]
- Su, Y.; Peng, Q.; Ling, H.; You, C.; Wu, Q.; Xu, L.; Que, Y. Systematic identification of miRNA-regulatory networks unveils their potential roles in sugarcane response to Sorghum mosaic virus infection. BMC Plant Biol. 2022, 22, 247. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; He, H.; Yan, M.; Zhao, C.; Lei, C.; Li, J.; Yan, F. Widely targeted analysis of metabolomic changes of Cucumis sativus induced by cucurbit chlorotic yellows virus. BMC Plant Biol. 2022, 22, 158. [Google Scholar] [CrossRef] [PubMed]
- Ontiveros, I.; López-Moya, J.J.; Díaz-Pendón, J.A. Coinfection of Tomato Plants with Tomato yellow leaf curl virus and Tomato chlorosis virus Affects the Interaction with Host and Whiteflies. Phytopathology 2022, 112, 944–952. [Google Scholar] [CrossRef]
- Nebapure, S.M.; Shankarganesh, K.; Rajna, S.; Naga, K.C.; Pandey, D.; Gambhir, S.; Praveen, K.V.; Subramanian, S. Dynamic changes in virus-induced volatiles in cotton modulate the orientation and oviposition behavior of the whitefly Bemisia tabaci. Front. Physiol. 2022, 13, 1017948. [Google Scholar] [CrossRef]
- van Houte, S.; Ros, V.I.; Mastenbroek, T.G.; Vendrig, N.J.; Hoover, K.; Spitzen, J.; van Oers, M.M. Protein tyrosine phosphatase-induced hyperactivity is a conserved strategy of a subset of baculoviruses to manipulate lepidopteran host behavior. PLoS ONE 2012, 7, e46933. [Google Scholar] [CrossRef]
- Ros, V.I.; van Houte, S.; Hemerik, L.; van Oers, M.M. Baculovirus-induced tree-top disease: How extended is the role of egt as a gene for the extended phenotype? Mol. Ecol. 2015, 24, 249–258. [Google Scholar] [CrossRef]
- Han, Y.; van Houte, S.; Drees, G.F.; van Oers, M.M.; Ros, V.I. Parasitic Manipulation of Host Behaviour: Baculovirus SeMNPV EGT Facilitates Tree-Top Disease in Spodoptera exigua Larvae by Extending the Time to Death. Insects 2015, 6, 716–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chesnais, Q.; Golyaev, V.; Velt, A.; Rustenholz, C.; Brault, V.; Pooggin, M.M.; Drucker, M. Comparative Plant Transcriptome Profiling of Arabidopsis thaliana Col-0 and Camelina sativa var. Celine Infested with Myzus persicae Aphids Acquiring Circulative and Noncirculative Viruses Reveals Virus- and Plant-Specific Alterations Relevant to Aphid Feeding Behavior and Transmission. Microbiol. Spectr. 2022, 10, e0013622. [Google Scholar] [PubMed]
- Minato, N.; Hatori, S.; Okawa, A.; Nakagawa, K.; Hironaka, M. Manipulation of Insect Vectors’ Host Selection Behavior by Barley Yellow Dwarf Virus Is Dependent on the Host Plant Species and Viral Co-Infection. Life 2022, 12, 644. [Google Scholar] [CrossRef] [PubMed]
- Todd, J.C.; Stewart, L.R.; Redinbaugh, M.G.; Wilson, J.R. Soybean Aphid (Hemiptera: Aphididae) Feeding Behavior is Largely Unchanged by Soybean Mosaic Virus but Significantly Altered by the Beetle-Transmitted Bean Pod Mottle Virus. J. Econ. Entomol. 2022, 115, 1059–1068. [Google Scholar] [CrossRef]
- Marmonier, A.; Velt, A.; Villeroy, C.; Rustenholz, C.; Chesnais, Q.; Brault, V. Differential gene expression in aphids following virus acquisition from plants or from an artificial medium. BMC Genom. 2022, 23, 333. [Google Scholar] [CrossRef]
- Patton, M.F.; Hansen, A.K.; Casteel, C.L. Potato leafroll virus reduces Buchnera aphidocola titer and alters vector transcriptome responses. Sci. Rep. 2021, 11, 23931. [Google Scholar] [CrossRef]
- Ghorbel, M.; Brini, F.; Sharma, A.; Landi, M. Role of jasmonic acid in plants: The molecular point of view. Plant Cell Rep. 2021, 40, 1471–1494. [Google Scholar] [CrossRef]
- Mostafa, S.; Wang, Y.; Zeng, W.; Jin, B. Plant Responses to Herbivory, Wounding, and Infection. Int. J. Mol. Sci. 2022, 23, 7031. [Google Scholar] [CrossRef]
- Parmagnani, A.S.; Maffei, M.E. Calcium Signaling in Plant-Insect Interactions. Plants 2022, 11, 2689. [Google Scholar] [CrossRef]
- Stroud, E.A.; Jayaraman, J.; Templeton, M.D.; Rikkerink, E.H.A. Comparison of the pathway structures influencing the temporal response of salicylate and jasmonate defence hormones in Arabidopsis thaliana. Front. Plant Sci. 2022, 13, 952301. [Google Scholar] [CrossRef] [PubMed]
- Pamplona, R.S.; Kim, J.; Lee, J.W.; Kim, C.S.; Boo, K.-H. Comparative transcriptome analysis of spinach in response to insect herbivory. Plant Biotechnol. Rep. 2022, 16, 43–55. [Google Scholar] [CrossRef]
- Tripathi, D.; Tanaka, K. A crosstalk between extracellular ATP and jasmonate signaling pathways for plant defense. Plant Signal. Behav. 2018, 13, e1432229. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gong, Q.; Wu, Y.; Huang, F.; Ismayil, A.; Zhang, D.; Li, H.; Gu, H.; Ludman, M.; Fátyol, K.; et al. A calmodulin-binding transcription factor links calcium signaling to antiviral RNAi defense in plants. Cell Host Microbe 2021, 29, 1393–1406.e7. [Google Scholar] [CrossRef] [PubMed]
- Baulcombe, D.C. The Role of Viruses in Identifying and Analyzing RNA Silencing. Annu. Rev. Virol. 2022, 9, 353–373. [Google Scholar] [CrossRef]
- Hu, J.; Huang, J.; Xu, H.; Wang, Y.; Li, C.; Wen, P.; You, X.; Zhang, X.; Pan, G.; Li, Q.; et al. Rice stripe virus suppresses jasmonic acid-mediated resistance by hijacking brassinosteroid signaling pathway in rice. PLoS Pathog. 2020, 16, e1008801. [Google Scholar] [CrossRef]
- Ji, M.; Zhao, J.; Han, K.; Cui, W.; Wu, X.; Chen, B.; Lu, Y.; Peng, J.; Zheng, H.; Rao, S.; et al. Turnip mosaic virus P1 suppresses JA biosynthesis by degrading cpSRP54 that delivers AOCs onto the thylakoid membrane to facilitate viral infection. PLoS Pathog. 2021, 17, e1010108. [Google Scholar] [CrossRef]
- Li, P.; Liu, H.; Li, F.; Liao, X.; Ali, S.; Hou, M. A virus plays a role in partially suppressing plant defenses induced by the viruliferous vectors. Sci. Rep. 2018, 8, 9027. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Weldegergis, B.T.; Li, J.; Jung, C.; Qu, J.; Sun, Y.; Qian, H.; Tee, C.; van Loon, J.J.; Dicke, M.; et al. Virulence factors of geminivirus interact with MYC2 to subvert plant resistance and promote vector performance. Plant Cell 2014, 26, 4991–5008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westwood, J.H.; Lewsey, M.G.; Murphy, A.M.; Tungadi, T.; Bates, A.; Gilligan, C.A.; Carr, J.P. Interference with jasmonic acid-regulated gene expression is a general property of viral suppressors of RNA silencing but only partly explains virus-induced changes in plant-aphid interactions. J. Gen. Virol. 2014, 95 Pt 3, 733–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Zhang, J.; Wang, J.; Liu, W.; Wang, K.; Chen, X.; Wen, Y.; Tian, S.; Pu, Y.; Fan, G.; et al. Tobacco mosaic virus hijacks its coat protein-interacting protein IP-L to inhibit NbCML30, a calmodulin-like protein, to enhance its infection. Plant J. 2022, 112, 677–693. [Google Scholar] [CrossRef]
- Schnettler, E.; Hemmes, H.; Huismann, R.; Goldbach, R.; Prins, M.; Kormelink, R. Diverging affinity of tospovirus RNA silencing suppressor proteins, NSs, for various RNA duplex molecules. J. Virol. 2010, 84, 11542–11554. [Google Scholar] [CrossRef] [Green Version]
- Hedil, M.; de Ronde, D.; Kormelink, R. Biochemical analysis of NSs from different tospoviruses. Virus Res. 2017, 242, 149–155. [Google Scholar] [CrossRef]
- Zhai, Y.; Gnanasekaran, P.; Pappu, H.R. Identification and Characterization of Plant-Interacting Targets of Tomato Spotted Wilt Virus Silencing Suppressor. Pathogens 2021, 10, 27. [Google Scholar] [CrossRef]
- Li, F.; Huang, C.; Li, Z.; Zhou, X. Suppression of RNA silencing by a plant DNA virus satellite requires a host calmodulin-like protein to repress RDR6 expression. PLoS Pathog. 2014, 10, e1003921. [Google Scholar] [CrossRef]
- Kamal, H.; Minhas, F.A.; Tripathi, D.; Abbasi, W.A.; Hamza, M.; Mustafa, R.; Khan, M.Z.; Mansoor, S.; Pappu, H.R.; Amin, I. βC1, pathogenicity determinant encoded by Cotton leaf curl Multan betasatellite, interacts with calmodulin-like protein 11 (Gh-CML11) in Gossypium hirsutum. PLoS ONE 2019, 14, e0225876. [Google Scholar] [CrossRef]
- Anandalakshmi, R.; Marathe, R.; Ge, X.; Herr, J.M., Jr.; Mau, C.; Mallory, A.; Pruss, G.; Bowman, L.; Vance, V.B. A calmodulin-related protein that suppresses posttranscriptional gene silencing in plants. Science 2000, 290, 142–144. [Google Scholar] [CrossRef] [PubMed]
- Jeon, E.J.; Tadamura, K.; Murakami, T.; Inaba, J.I.; Kim, B.M.; Sato, M.; Atsumi, G.; Kuchitsu, K.; Masuta, C.; Nakahara, K.S. rgs-CaM Detects and Counteracts Viral RNA Silencing Suppressors in Plant Immune Priming. J. Virol. 2017, 91, e00761-e17. [Google Scholar] [CrossRef] [Green Version]
- Timmons, L.; Fire, A. Specific interference by ingested dsRNA. Nature 1998, 395, 854. [Google Scholar] [CrossRef] [PubMed]
- Hua, C.; Zhao, J.H.; Guo, H.S. Trans-Kingdom RNA Silencing in Plant-Fungal Pathogen Interactions. Mol. Plant 2018, 11, 235–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaloner, T.; van Kan, J.A.L.; Grant-Downton, R.G.T. RNA ‘Information Warfare’ in pathogenic and mutualistic interactions. Trends Plant Sci. 2016, 21, 738–748. [Google Scholar] [CrossRef]
- van den Heuvel, J.F.; Bruyère, A.; Hogenhout, S.A.; Ziegler-Graff, V.; Brault, V.; Verbeek, M.; van der Wilk, F.; Richards, K. The N-terminal region of the luteovirus readthrough domain determines virus binding to Buchnera GroEL and is essential for virus persistence in the aphid. J. Virol. 1997, 71, 7258–7265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallis, S.; Alyokhin, A.; Manley, B.; Rodrigues, T.B.; Buzza, A.; Barnes, E.; Narva, K. Toxicity of a novel dsRNA-based insecticide to the Colorado potato beetle in laboratory and field trials. Pest Manag. Sci. 2022, 78, 3836–3848. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.; Zeng, Q.; She, L.; Yuan, H.; Luo, Y.; Wang, R.; She, Y.; Wang, W.; Wang, C.; Pan, X. WolbachiaUtilizes lncRNAs to Activate the Anti-Dengue Toll Pathway and Balance Reactive Oxygen Species Stress in Aedes aegypti Through a Competitive Endogenous RNA Network. Front. Cell Infect. Microbiol. 2022, 11, 823403. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Wei, Y.; Zhao, C.; Li, S.; Gao, S.; Zhang, Y.; Wu, Y.; Lu, C. Facultative symbionts are potential agents of symbiont-mediated RNAi in aphids. Front. Microbiol. 2022, 13, 1020461. [Google Scholar] [CrossRef]
- Lefebvre, F.A.; Lécuyer, E. Small Luggage for a Long Journey: Transfer of Vesicle-Enclosed Small RNA in Interspecies Communication. Front. Microbiol. 2017, 8, 377. [Google Scholar] [CrossRef] [Green Version]
- Kormelink, R.; Verchot, J.; Tao, X.; Desbiez, C. The Bunyavirales: The Plant-Infecting Counterparts. Viruses 2021, 13, 842. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Virus | Studied Host | Target | Implicated Role | Reference |
|---|---|---|---|---|
| Cucumber mosaic virus (CMV) Y satellite RNA (Y-sat) | Nicotiana tabacum | Magnesium protoporphyrin chelatase subunit I gene (ChlI) | Disorder in chlorophyll synthesis; yellowing of the leaves | [32,33] |
| Tobacco mosaic virus (TMV) | Nicotiana benthamiana | C2-domain abscisic acid (ABA)-related (CAR) 7-like protein | Interfere with the levels of abscisic acid (ABA) | [35] |
| Chinese wheat mosaic virus (CWMV) | Triticum aestivum | Vacuolar (H+)-PPases (VPs) | Suppression of cell death induced by VPs | [36] |
| Rice stripe tenuivirus (RSV) | Nicotiana benthamiana; Rice | Several chloroplast-related genes; Eukaryotic translation initiation factor 4A (eIF4E) | Leaf-twisting and stunting; activation of antiviral autophagy | [37,38] |
| Wheat yellow mosaic virus (WYMV) | Wheat | Thioredoxin-like gene (TaAAED1) | Interfere with the scavenging of reactive oxygen species (ROS) | [39] |
| Plant Biological Process | Virus-Derived sRNA | Potential Modulatory Effect | Reference |
|---|---|---|---|
| Stress responses | vsiRNAs; vmiRNAs | Decrease expression of stress resistance genes leading to increased susceptibility of the host to other (a)biotic stimuli. | [11,40,41,42] |
| Protein synthesis | vsiRNAs | Decrease expression of plant proteins involved in the antiviral response. | [40,43] |
| Chloroplast functions | vsiRNAs | Induce development of leaf-associated disease symptoms | [41,44] |
| Development | vsiRNAs | Modulate genes involved in pollen or anatomical structure development. | [41,45] |
| Cellular structure | vsiRNAs | Modulate expression of cellular structural proteins, such as kinesins and ribosomal proteins, interfering with general aspects of the cellular functioning. | [46,47,48] |
| Cell metabolism | vsiRNAs; vmiRNAs | Modulate expression of enzymes involved in biosynthesis of secondary metabolites, amino acids, starch and sucrose metabolism, and carbon metabolism. | [42,46,47,48,49,50,51] |
| Host defense pathways | vsiRNAs; vmiRNAs | Modulate expression of transcription factors (TFs), nucleotide binding site–leucine-rich repeats (NBS LRR), and receptor-like protein (RLP) kinase involved in host defense pathways. | [11,41,43,45,46,47,49,52] |
| Host RNA silencing pathway | vsiRNAs | Modulate expression of RNA silencing-associated factors (e.g., DCLs and AGO) involved in antiviral RNA-based response. | [41] |
| Synergistic infection | vsiRNAs; vmiRNAs | Modulate pathways to enhance synergistic infection and virulence of multiple viruses. | [41,50,53] |
| Virus (and Inducer) | vamiRNAs | Host | Targets | Implicated Role | Reference |
|---|---|---|---|---|---|
| CymRSV (p19), CrTMV, PVX, TEV, TCV, RMV, SHMV | miRNA168 | Arabidopsis thaliana, N. benthamiana, Medicago truncalata, Solanum lycopersicum | AGO1 | Translational repression of AGO1; alleviates antiviral RNAi pressure, and benefits viral infections. | [61] |
| PVY | miRNAs | N. tabacum | Genes from a broad range of cellular processes | Regulation of plant resistance to PVY. | [64] |
| PVY | miRNAs | S. lycopersicum | R genes, MAPKs and disease-responsive genes | A functional role of sRNA-mediated defenses in the recovery phenotype of tomato. | [67] |
| RSV | miRNA444 | Oryza sativa | MIKCC-type MADS box proteins OsMADS23, OsMADS27a, and OsMADS57 | Relieve the repression of RDR1 by MADS box proteins. | [52] |
| TSWV | miRNAs | Capsicum | ND | ND | [68] |
| SrMV | miRNAs | Saccharum spp. | Various | Associated with early immune response upon virus infection. | [69] |
| Virus (and Inducer) | vasiRNAs | Host | Targets | Implicated Role | Reference |
| ORMV | miRNAs, ml-siRNA, vasiRNAs | A. thaliana | Various | At the time point of sRNA analysis, no major changes in RNA targets were observed. | [29] |
| CMV, TuMv | vasiRNAs | A. thaliana | Various; genes responsive to biotic and abiotic stimuli are significantly enriched | Broad spectrum viral defense. | [30] |
| CaMV | vasiRNAs | A. thaliana, Brassica rapa, B. napus | 13 loci (targets) out of 15, from which vasiRNAs derive, perfectly match those from Cao et al. [30]. | Loci/CDS targets encode factors involved in photosynthesis and stress response (e.g., Rubisco activase (RCA), senescence-associated protein, heat shock protein HSP70, light harvesting complex, and membrane-related protein CP5). During infection, the expression of these factors is significantly downregulated, suggesting that their silencing is a central component of the plant’s response to virus infections. | [62] |
| TSWV, CaCV | vasiRNAs | Capsicum annuum cv. Yolo Wonder, N. benthamiana LAB (RDR1-deficient), and N. benthamiana WA, Tomato Marglobe (Sw5−), Red Defender (Sw5+) | Similar to those reported by Cao et al. [30] and enriched for ribosomal protein-encoding genes and genes involved in protein processing at the endoplasmic reticulum. | [63] | |
| TuMV | vasiRNAs | B. napus (local) | Host genes and viral sequences | vasiRNAs, hsiRNAs and vsiRNAs are observed to target host as well as vRNA molecules to potentially control gene homeostasis during virus infection, and therefore contributing to virus–host compatibility. | [26] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsumura, E.E.; Kormelink, R. Small Talk: On the Possible Role of Trans-Kingdom Small RNAs during Plant–Virus–Vector Tritrophic Communication. Plants 2023, 12, 1411. https://doi.org/10.3390/plants12061411
Matsumura EE, Kormelink R. Small Talk: On the Possible Role of Trans-Kingdom Small RNAs during Plant–Virus–Vector Tritrophic Communication. Plants. 2023; 12(6):1411. https://doi.org/10.3390/plants12061411
Chicago/Turabian StyleMatsumura, Emilyn E., and Richard Kormelink. 2023. "Small Talk: On the Possible Role of Trans-Kingdom Small RNAs during Plant–Virus–Vector Tritrophic Communication" Plants 12, no. 6: 1411. https://doi.org/10.3390/plants12061411