
DDM1-Mediated TE Silencing in Plants


Abstract
:1. Introduction
2. DDM1 in Trait Regulation
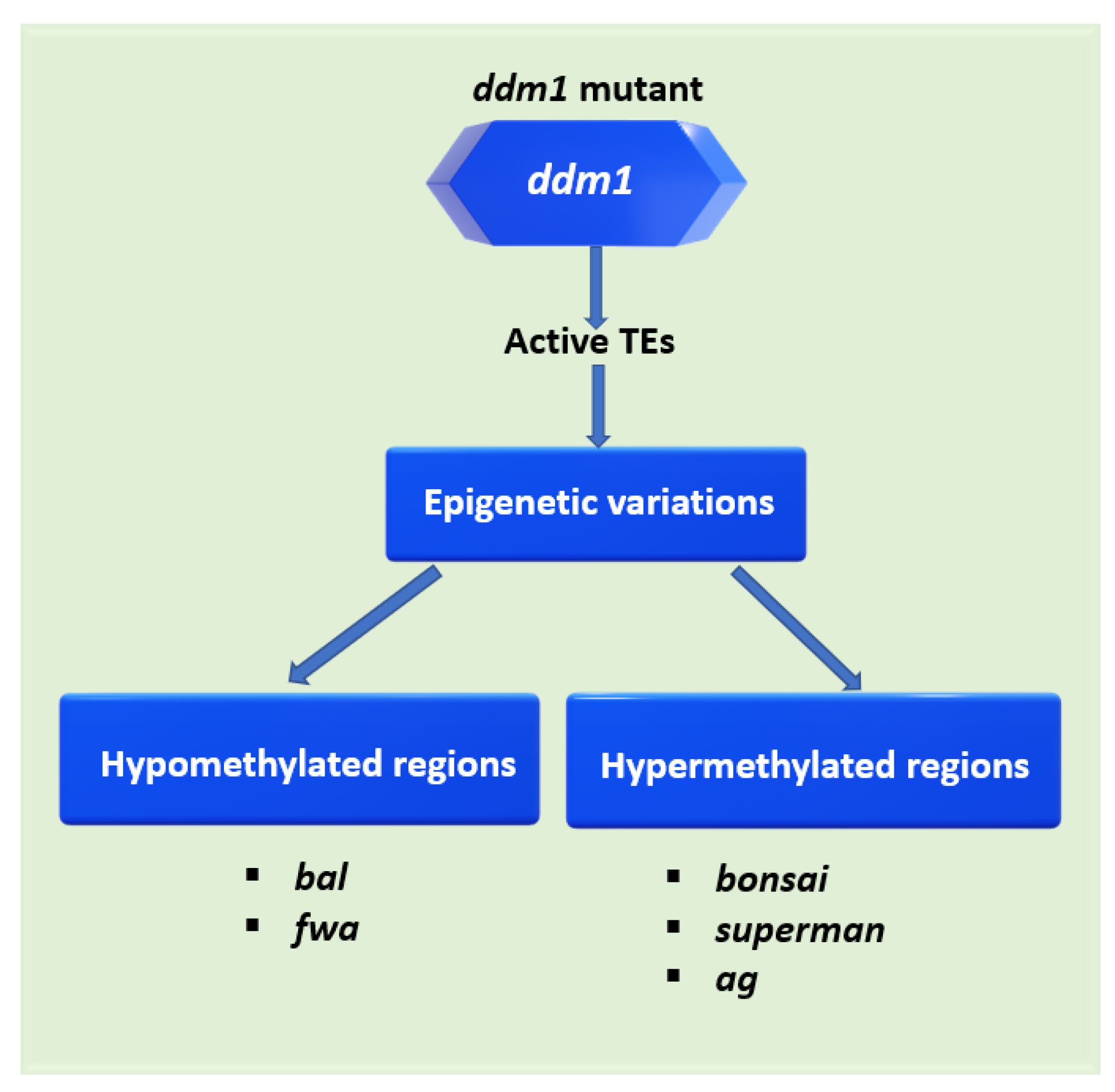
2.1. DDM1 Participates in a Broad Range of Plant Traits
2.2. DDM1 Is Genetically Distinct from DDM2
3. DDM1 in the RdDM Context
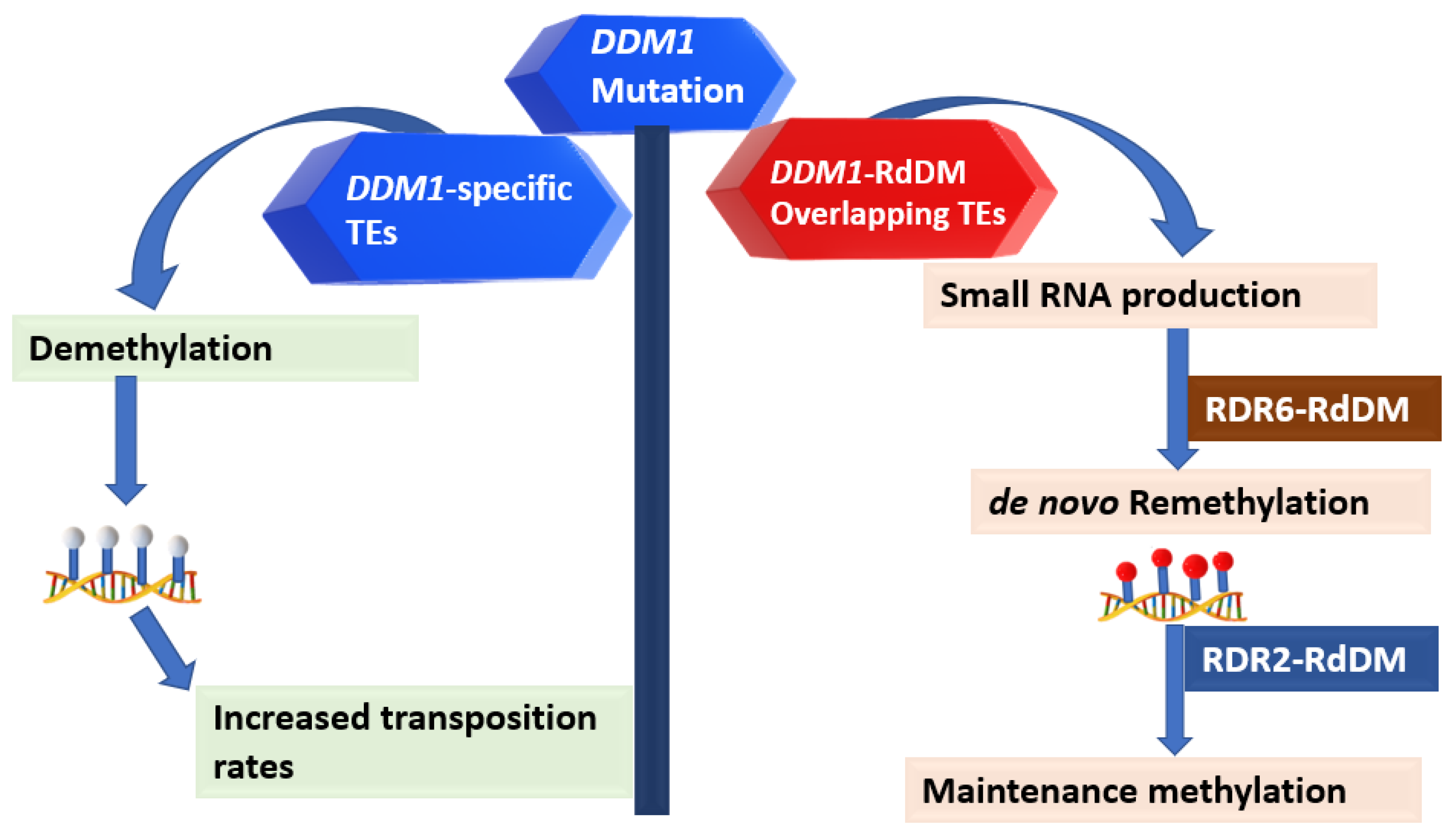
3.1. DDM1 Collaborates with RdDM in Whole-Genome TE Silencing
3.2. The Collaboration between DDM1 and the RdDM Is Linked to TE Activity and Small RNA Dynamics in the Genome
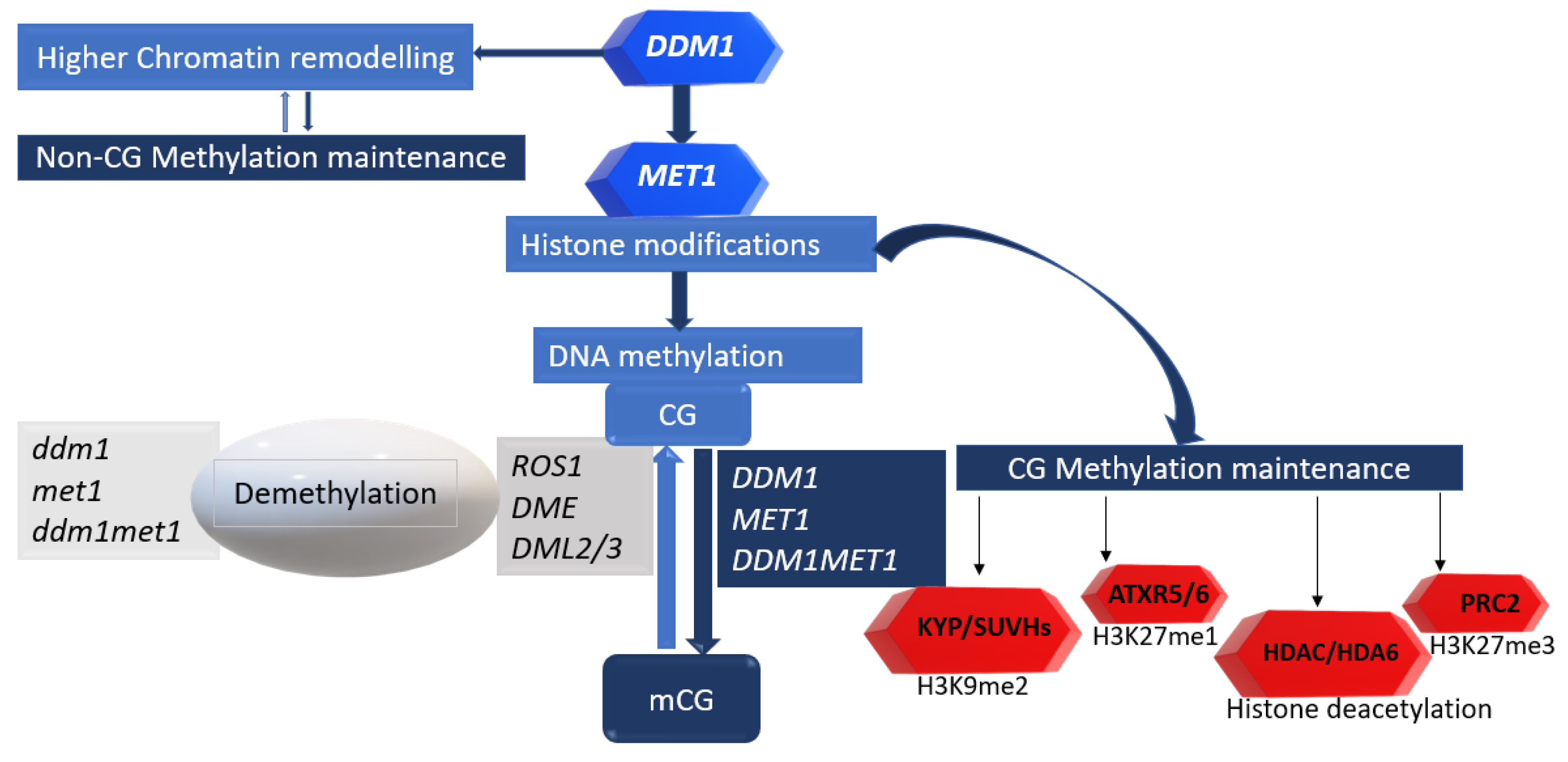
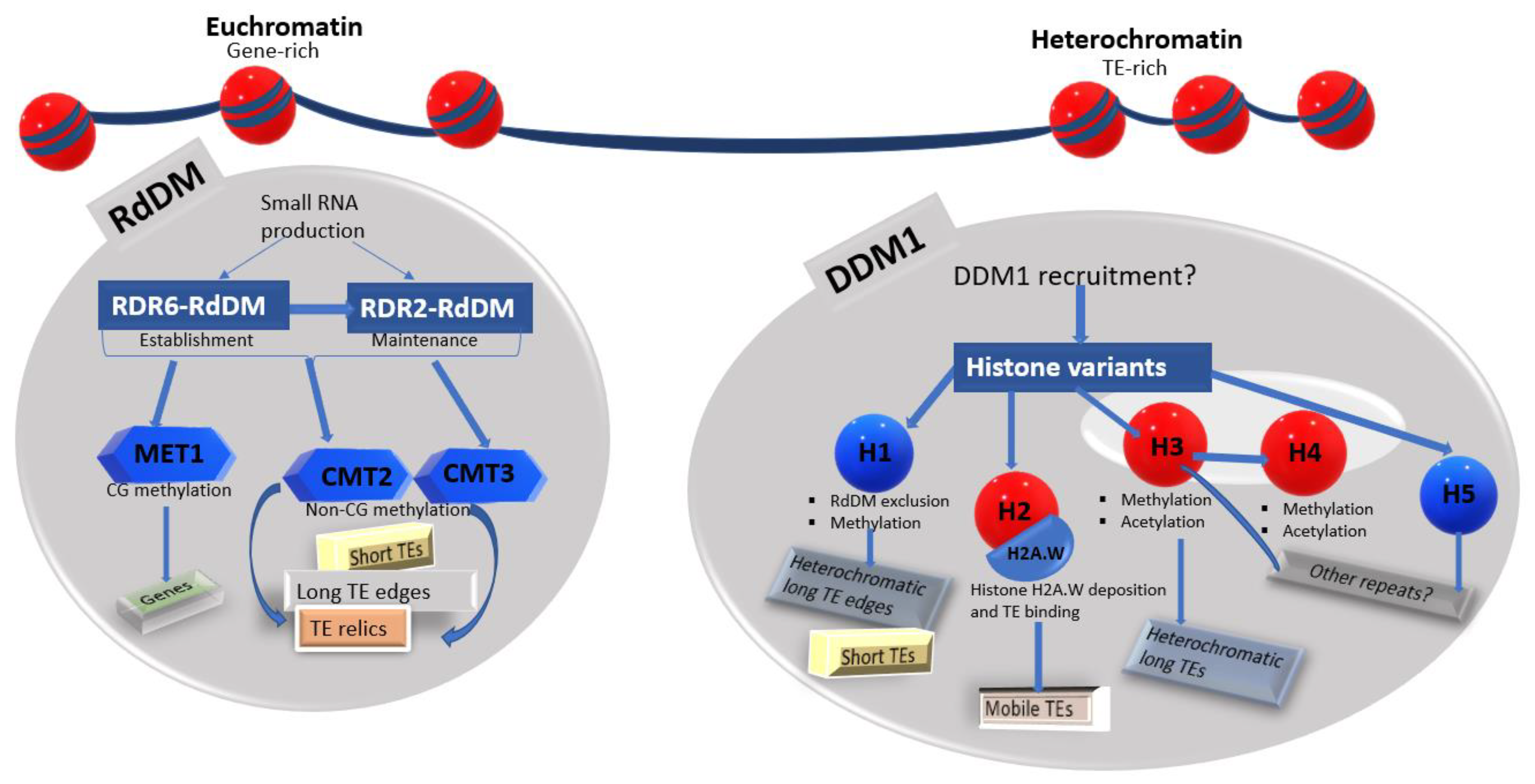
4. DDM1 Is More Active in the Heterochromatin
4.1. DDM1 Is Required to Preserve Heterochromatic Features, and This Is Independent of the RdDM Pathway
4.2. DDM1 Activity in the Heterochromatin Extends to the Histone Core to Silence the Majority of Mobile TEs
5. Conclusions and Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- O’Neill, K.; Brocks, D.; Hammell, M.G. Mobile genomics: Tools and techniques for tackling transposons. Philos. Trans. Biol. Sci. 2020, 375, 20190345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wessler, S.R. Transposable Elements and the Evolution of Eukaryotic Genomes. Proc. Natl. Acad. Sci. USA 2006, 103, 17600–17601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourque, G.; Burns, K.H.; Gehring, M.; Gorbunova, V.; Seluanov, A.; Hammell, M.; Imbeault, M.; Izsvák, Z.; Levin, H.L.; Macfarlan, T.S.; et al. Ten things you should know about transposable elements. Genome Biol. 2018, 19, 199. [Google Scholar] [CrossRef]
- Hallet, B.; Sherratt, D.J. Transposition and site-specific recombination: Adapting DNA cut-and-paste mechanisms to a variety of genetic rearrangements. FEMS Microbiol. Rev. 1997, 21, 157–178. [Google Scholar] [CrossRef] [PubMed]
- Lisch, D. How important are transposons for plant evolution? Nat. Rev. Genet. 2013, 14, 49–61. [Google Scholar] [CrossRef]
- Bennetzen, J.L.; Wang, H. The contributions of transposable elements to the structure, function, and evolution of plant genomes. Annu. Rev. Plant Biol. 2014, 65, 505–530. [Google Scholar] [CrossRef] [PubMed]
- Grewal, S.I.S.; Jia, S. Heterochromatin revisited. Nat. Rev. Genet. 2007, 8, 35–46. [Google Scholar] [CrossRef]
- Baidouri, M.E.; Carpentier, M.-C.; Cooke, R.; Gao, D.; Lasserre, E.; Llauro, C.; Mirouze, M.; Picault, N.; Jackson, S.A.; Panaud, O. Widespread and frequent horizontal transfers of transposable elements in plants. Genome Res. 2014, 24, 831–838. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.-H.; Peccoud, J.; Xu, M.-R.-X.; Zhang, X.-G.; Gilbert, C. Horizontal transfer and evolution of transposable elements in vertebrates. Nat. Commun. 2020, 11, 1362. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Liang, G.; Molloy, P.L.; Jones, P.A. DNA methylation enables transposable element-driven genome expansion. Proc. Natl. Acad. Sci. USA 2020, 117, 19359–19366. [Google Scholar] [CrossRef]
- Zemach, A.; Kim, M.Y.; Hsieh, P.-H.; Coleman-Derr, D.; Eshed-Williams, L.; Thao, K.; Harmer, S.L.; Zilberman, D. The Arabidopsis Nucleosome Remodeler DDM1 Allows DNA Methyltransferases to Access H1-Containing Heterochromatin. Cell 2013, 153, 193–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erdmann, R.M.; Picard, C.L. RNA-directed DNA Methylation. PLoS Genet. 2020, 16, e1009034. [Google Scholar] [CrossRef] [PubMed]
- Sammarco, I.; Pieters, J.; Salony, S.; Toman, I.; Zolotarov, G.; Lafon Placette, C. Epigenetic targeting of transposon relics: Beating the dead horses of the genome? Epigenetics 2022, 17, 1331–1344. [Google Scholar] [CrossRef] [PubMed]
- Papareddy, R.K.; Páldi, K.; Paulraj, S.; Kao, P.; Lutzmayer, S.; Nodine, M.D. Chromatin regulates expression of small RNAs to help maintain transposon methylome homeostasis in Arabidopsis. Genome Biol. 2020, 21, 251. [Google Scholar] [CrossRef]
- McCue, A.D.; Panda, K.; Nuthikattu, S.; Choudury, S.G.; Thomas, E.N.; Slotkin, R.K. ARGONAUTE 6 bridges transposable element mRNA-derived siRNAs to the establishment of DNA methylation. EMBO J. 2015, 34, 20–35. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, T.; Kobayashi, A.; Saze, H.; Kakutani, T. RNAi-independent de novo DNA methylation revealed in Arabidopsis mutants of chromatin remodeling gene DDM1: RNAi-independent de novo DNA methylation. Plant J. Cell Mol. Biol. 2012, 70, 750–758. [Google Scholar] [CrossRef]
- Czajka, K.; Mehes-Smith, M.; Nkongolo, K. DNA methylation and histone modifications induced by abiotic stressors in plants. Genes Genom. 2022, 44, 279–297. [Google Scholar] [CrossRef]
- Scott, R.J.; Spielman, M.; Bailey, J.; Dickinson, H.G. Parent-of-origin effects on seed development in Arabidopsis thaliana. Development 1998, 125, 3329–3341. [Google Scholar] [CrossRef]
- Satyaki, P.R.V.; Gehring, M. Paternally Acting Canonical RNA-Directed DNA Methylation Pathway Genes Sensitize Arabidopsis Endosperm to Paternal Genome Dosage. Plant Cell 2019, 31, 1563–1578. [Google Scholar] [CrossRef] [Green Version]
- Kankel, M.W.; Ramsey, D.E.; Stokes, T.L.; Flowers, S.K.; Haag, J.R.; Jeddeloh, J.A.; Riddle, N.C.; Verbsky, M.L.; Richards, E.J. Arabidopsis MET1 Cytosine Methyltransferase Mutants. Genetics 2003, 163, 1109–1122. [Google Scholar] [CrossRef]
- Pavlopoulou, A.; Kossida, S. Plant cytosine-5 DNA methyltransferases: Structure, function, and molecular evolution. Genomics 2007, 90, 530–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osakabe, A.; Jamge, B.; Axelsson, E.; Montgomery, S.A.; Akimcheva, S.; Kuehn, A.L.; Pisupati, R.; Lorković, Z.J.; Yelagandula, R.; Kakutani, T.; et al. The chromatin remodeler DDM1 prevents transposon mobility through deposition of histone variant H2A.W. Nat. Cell Biol. 2021, 23, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Lyons, D.B.; Zilberman, D. DDM1 and lsh remodelers allow methylation of DNA wrapped in nucleosomes. Elife 2017, 6, e30674. [Google Scholar] [CrossRef]
- Soppe, W.J.J.; Jasencakova, Z.; Houben, A.; Kakutani, T.; Meister, A.; Huang, M.S.; Jacobsen, S.E.; Schubert, I.; Fransz, P.F. DNA methylation controls histone H3 lysine 9 methylation and heterochromatin assembly in Arabidopsis. EMBO J. 2002, 21, 6549–6559. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Li, Y.; Xu, T.; Srivastava, A.K.; Wang, D.; Zeng, L.; Yang, L.; He, L.; Zhang, H.; Zheng, Z.; et al. The chromatin remodeler DDM1 promotes hybrid vigor by regulating salicylic acid metabolism. Cell Discov. 2016, 2, 16027. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.H.; Ryu, T.H.; Kim, J.-I.; Lee, S.; Lee, S.S.; Kim, J.-H. Mutation in DDM1 inhibits the homology directed repair of double strand breaks. PLoS ONE 2019, 14, e0211878. [Google Scholar] [CrossRef] [PubMed]
- Scheid, O.M.; Afsar, K.; Paszkowski, J. Release of epigenetic gene silencing by trans-acting mutations in Arabidopsis. Proc. Natl. Acad. Sci. USA 1998, 95, 632–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaked, H.; Avivi-Ragolsky, N.; Levy, A.A. Involvement of the Arabidopsis SWI2/SNF2 Chromatin Remodeling Gene Family in DNA Damage Response and Recombination. Genetics 2006, 173, 985–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stokes, T.L.; Kunkel, B.N.; Richards, E.J. Epigenetic variation in Arabidopsis disease resistance. Genes Dev. 2002, 16, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Saze, H.; Kakutani, T. Heritable epigenetic mutation of a transposon-flanked Arabidopsis gene due to lack of the chromatin-remodeling factor DDM1. EMBO J. 2007, 26, 3641–3652. [Google Scholar] [CrossRef]
- Liu, F.; Gao, C.; Chen, M.; Tang, G.; Sun, Y.; Li, K. The impacts of flowering phenology on the reproductive success of the narrow endemic Nouelia insignis Franch. (Asteraceae). Ecol. Evol. 2021, 11, 9396–9409. [Google Scholar] [CrossRef] [PubMed]
- Zu, P.; Schiestl, F.P. The effects of becoming taller: Direct and pleiotropic effects of artificial selection on plant height in Brassica rapa. Plant J. Cell Mol. Biol. 2017, 89, 1009–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soppe, W.J.J.; Jacobsen, S.E.; Alonso-Blanco, C.; Jackson, J.P.; Kakutani, T.; Koornneef, M.; Peeters, A.J.M. The Late Flowering Phenotype of fwa Mutants Is Caused by Gain-of-Function Epigenetic Alleles of a Homeodomain Gene. Mol. Cell 2000, 6, 791–802. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, S.E.; Sakai, H.; Finnegan, E.J.; Cao, X.; Meyerowitz, E.M. Ectopic hypermethylation of flower-specific genes in Arabidopsis. Curr. Biol. 2000, 10, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Jacobsen, S.E. Hypermethylated SUPERMAN epigenetic alleles in Arabidopsis. Science 1997, 277, 1100–1104. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Eichten, S.R.; Hermanson, P.J.; Zaunbrecher, V.M.; Song, J.; Wendt, J.; Rosenbaum, H.; Madzima, T.F.; Sloan, A.E.; Huang, J.; et al. Genetic Perturbation of the Maize Methylome. Plant Cell 2014, 26, 4602–4616. [Google Scholar] [CrossRef] [Green Version]
- Corem, S.; Doron-Faigenboim, A.; Jouffroy, O.; Maumus, F.; Arazi, T.; Bouché, N. Redistribution of CHH methylation and small interfering RNAs across the genome of tomato ddm1 mutants. Plant Cell 2018, 30, 1628–1644. [Google Scholar] [CrossRef] [Green Version]
- Higo, A.; Saihara, N.; Miura, F.; Higashi, Y.; Yamada, M.; Tamaki, S.; Ito, T.; Tarutani, Y.; Sakamoto, T.; Fujiwara, M.; et al. DNA methylation is reconfigured at the onset of reproduction in rice shoot apical meristem. Nat. Commun. 2020, 11, 4079. [Google Scholar] [CrossRef]
- Long, J.C.; Xia, A.A.; Liu, J.H.; Jing, J.L.; Wang, Y.Z.; Qi, C.Y.; He, Y. Decrease in DNA methylation 1 (DDM1) is required for the formation of mCHH islands in maize. J. Integr. Plant Biol. 2019, 61, 749–764. [Google Scholar] [CrossRef] [Green Version]
- Niederhuth, C.E.; Schmitz, R.J. Putting DNA methylation in context: From genomes to gene expression in plants. Biochim. Biophys. Acta Gene Regul. Mech. 2017, 1860, 149–156. [Google Scholar] [CrossRef]
- Blevins, T.; Pontes, O.; Pikaard, C.S.; Meins, F.J. Heterochromatic siRNAs and DDM1 independently silence aberrant 5S rDNA transcripts in Arabidopsis. PLoS ONE 2009, 4, e5932. [Google Scholar] [CrossRef] [PubMed]
- Stroud, H.; Do, T.; Du, J.; Zhong, X.; Feng, S.; Johnson, L.; Patel, D.J.; Jacobsen, S.E. Non-CG methylation patterns shape the epigenetic landscape in Arabidopsis. Nat. Struct. Mol. Biol. 2014, 21, 64–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakutani, T.; Jeddeloh, J.A.; Flowers, S.K.; Munakata, K.; Richards, E.J. Developmental abnormalities and epimutations associated with DNA hypomethylation mutations. Proc. Natl. Acad. Sci. USA 1996, 93, 12406–12411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawanabe, T.; Ishikura, S.; Miyaji, N.; Sasaki, T.; Wu, L.M.; Itabashi, E.; Takada, S.; Shimizu, M.; Takasaki-Yasuda, T.; Osabe, K.; et al. Role of DNA methylation in hybrid vigor in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2016, 113, E6704–E6711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rougée, M.; Quadrana, L.; Zervudacki, J.; Hure, V.; Colot, V.; Navarro, L.; Deleris, A. Polycomb mutant partially suppresses DNA hypomethylation-associated phenotypes in Arabidopsis. Life Sci. Alliance 2021, 4, e202000848. [Google Scholar] [CrossRef] [PubMed]
- Stroud, H.; Hale, C.J.; Feng, S.; Caro, E.; Jacob, Y.; Michaels, S.D.; Jacobsen, S.E. DNA methyltransferases are required to induce heterochromatic re-replication in Arabidopsis. PLoS Genet. 2012, 8, e1002808. [Google Scholar] [CrossRef]
- Liu, X.; Yu, C.-W.; Duan, J.; Luo, M.; Wang, K.; Tian, G.; Cui, Y.; Wu, K. HDA6 Directly Interacts with DNA Methyltransferase MET1 and Maintains Transposable Element Silencing in Arabidopsis. Plant Physiol. 2012, 158, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Melamed-Bessudo, C.; Levy, A.A. Deficiency in DNA methylation increases meiotic crossover rates in euchromatic but not in heterochromatic regions in Arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, E981–E988. [Google Scholar] [CrossRef] [Green Version]
- He, X.-J.; Chen, T.; Zhu, J.-K. Regulation and function of DNA methylation in plants and animals. Cell Res. 2011, 21, 442–465. [Google Scholar] [CrossRef]
- Haag, J.R.; Ream, T.S.; Marasco, M.; Nicora, C.D.; Norbeck, A.D.; Pasa-Tolic, L.; Pikaard, C.S. In Vitro Transcription Activities of Pol IV, Pol V, and RDR2 Reveal Coupling of Pol IV and RDR2 for dsRNA Synthesis in Plant RNA Silencing. Mol. Cell 2012, 48, 811–818. [Google Scholar] [CrossRef]
- Blevins, T.; Podicheti, R.; Mishra, V.; Marasco, M.; Wang, J.; Rusch, D.; Tang, H.; Pikaard, C.S. Identification of pol IV and RDR2-dependent precursors of 24nt siRNAs guiding de novo DNA methylation in Arabidopsis. eLife 2015, 4, e09591. [Google Scholar] [CrossRef] [PubMed]
- Zhai, J.; Bischof, S.; Wang, H.; Feng, S.; Lee, T.-F.; Teng, C.; Chen, X.; Park, S.Y.; Liu, L.; Gallego-Bartolome, J.; et al. A One Precursor One siRNA Model for Pol IV-Dependent siRNA Biogenesis. Cell 2015, 163, 445–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, J.; Mishra, V.; Wang, F.; Huang, H.-Y.; Pikaard, C.S. Reaction Mechanisms of Pol IV, RDR2, and DCL3 Drive RNA Channeling in the siRNA-Directed DNA Methylation Pathway. Mol. Cell 2019, 75, 576–589.e575. [Google Scholar] [CrossRef]
- Pikaard, C.S.; Haag, J.R.; Pontes, O.M.F.; Blevins, T.; Cocklin, R. A transcription fork model for pol IV and pol V-dependent RNA-directed DNA methylation. Cold Spring Harb. Symp. Quant. Biol. 2012, 77, 205–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Zhao, C.; Zhang, Q.; Zinta, G.; Wang, D.; Lozano-Durán, R.; Zhu, J.-K. Pathway conversion enables a double-lock mechanism to maintain DNA methylation and genome stability. Proc. Natl. Acad. Sci. USA 2021, 118, e2107320118. [Google Scholar] [CrossRef]
- Greenberg, M.V.C.; Ausin, I.; Chan, S.W.L.; Cokus, S.J.; Cuperus, J.T.; Feng, S.; Law, J.A.; Chu, C.; Pellegrini, M.; Carrington, J.C.; et al. Identification of genes required for de novo DNA methylation in Arabidopsis. Epigenetics 2011, 6, 344–354. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.C.; Ernst, E.; Berube, B.; Borges, F.; Parent, J.-S.; Ledon, P.; Schorn, A.; Martienssen, R.A. Arabidopsis retrotransposon virus-like particles and their regulation by epigenetically activated small RNA. Genome Res. 2020, 30, 576–588. [Google Scholar] [CrossRef]
- Henderson, I.R.; Deleris, A.; Wong, W.; Zhong, X.; Chin, H.G.; Horwitz, G.A.; Kelly, K.A.; Pradhan, S.; Jacobsen, S.E. The De novo cytosine methyltransferase DRM2 requires intact UBA domains and a catalytically mutated paralog DRM3 during RNA-directed DNA methylation in Arabidopsis thaliana. PLoS Genet. 2010, 6, e1001182. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Zhong, X.; Bernatavichute, Y.V.; Stroud, H.; Feng, S.; Caro, E.; Vashisht, A.A.; Terragni, J.; Chin, H.G.; Tu, A.; et al. Dual Binding of Chromomethylase Domains to H3K9me2-Containing Nucleosomes Directs DNA Methylation in Plants. Cell 2012, 151, 167–180. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Johnson, L.M.; Jacobsen, S.E.; Patel, D.J. DNA methylation pathways and their crosstalk with histone methylation. Nat. Rev. Mol. Cell Biol. 2015, 16, 519–532. [Google Scholar] [CrossRef]
- Sigman, M.J.; Slotkin, R.K. The First Rule of Plant Transposable Element Silencing: Location, Location, Location. Plant Cell 2016, 28, 304–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeddeloh, J.A.; Bender, J.; Richards, E.J. The DNA methylation locus DDM1 is required for maintenance of gene silencing in Arabidopsis. Genes Dev. 1998, 12, 1714–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirochika, H.; Okamoto, H.; Kakutani, T. Silencing of Retrotransposons in Arabidopsis and Reactivation by the ddm1 Mutation. Plant Cell 2000, 12, 357–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panda, K.; Ji, L.; Neumann, D.A.; Daron, J.; Schmitz, R.J.; Slotkin, R.K. Full-length autonomous transposable elements are preferentially targeted by expression-dependent forms of RNA-directed DNA methylation. Genome Biol. 2016, 17, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, T.; Tarutani, Y.; To, T.K.; Kassam, M.; Duvernois-Berthet, E.; Cortijo, S.; Takashima, K.; Saze, H.; Toyoda, A.; Fujiyama, A. Genome-Wide Negative Feedback Drives Transgenerational DNA Methylation Dynamics in Arabidopsis. PLoS Genet. 2015, 11, e1005154. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Hatlen, A.; Kelly, L.J.; Becher, H.; Wang, W.; Kovarik, A.; Leitch, I.J.; Leitch, A.R. Angiosperms are unique among land plant lineages in the occurrence of key genes in the RNA-Directed DNA methylation (RdDM) Pathway. Genome Biol. Evol. 2015, 7, 2648–2662. [Google Scholar] [CrossRef] [Green Version]
- Basenko, E.Y.; Kamei, M.; Ji, L.; Schmitz, R.J.; Lewis, Z.A. The LSH/DDM1 Homolog MUS-30 Is Required for Genome Stability, but Not for DNA Methylation in Neurospora crassa. PLoS Genet. 2016, 12, e1005790. [Google Scholar] [CrossRef]
- Fu, F.-F.; Dawe, R.K.; Gent, J.I. Loss of RNA-directed DNA methylation in maize chromomethylase and DDM1-type nucleosome remodeler mutants. Plant Cell 2018, 30, 1617–1627. [Google Scholar] [CrossRef] [Green Version]
- El-Sappah, A.H.; Yan, K.; Huang, Q.; Islam, M.M.; Li, Q.; Wang, Y.; Khan, M.S.; Zhao, X.; Mir, R.R.; Li, J.; et al. Comprehensive Mechanism of Gene Silencing and Its Role in Plant Growth and Development. Front. Plant Sci. 2021, 12, 705249. [Google Scholar] [CrossRef]
- Vaucheret, H.; Fagard, M. Transcriptional gene silencing in plants: Targets, inducers and regulators. Trends Genet. 2001, 17, 29–35. [Google Scholar] [CrossRef]
- Morel, J.-B.; Mourrain, P.; Béclin, C.; Vaucheret, H. DNA methylation and chromatin structure affect transcriptional and post-transcriptional transgene silencing in Arabidopsis. Curr. Biol. 2000, 10, 1591–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, F.; Lu, Y.; Jiang, W.; Wu, T.; Zhang, R.; Zhao, Y.; Zhou, D.-X. DDM1 Represses Noncoding RNA Expression and RNA-Directed DNA Methylation in Heterochromatin. Plant Physiol. 2018, 177, 1187–1197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Gent, J.I.; Zynda, G.; Song, J.; Makarevitch, I.; Hirsch, C.D.; Hirsch, C.N.; Dawe, R.K.; Madzima, T.F.; McGinnis, K.M.; et al. RNA-directed DNA methylation enforces boundaries between heterochromatin and euchromatin in the maize genome. Proc. Natl. Acad. Sci. USA 2015, 112, 14728–14733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murcott, B.; Pawluk, R.J.; Protasio, A.V.; Akinmusola, R.Y.; Lastik, D.; Hunt, V.L. stepRNA: Identification of Dicer cleavage signatures and passenger strand lengths in small RNA sequences. Front. Bioinform. 2022, 2, 994871. [Google Scholar] [CrossRef] [PubMed]
- Henderson, I.R.; Zhang, X.; Lu, C.; Johnson, L.; Meyers, B.C.; Green, P.J.; Jacobsen, S.E. Dissecting Arabidopsis thaliana DICER function in small RNA processing, gene silencing and DNA methylation patterning. Nat. Genet. 2006, 38, 721–725. [Google Scholar] [CrossRef] [PubMed]
- Bourguet, P.; Picard, C.L.; Yelagandula, R.; Pélissier, T.; Lorković, Z.J.; Feng, S.; Pouch-Pélissier, M.-N.; Schmücker, A.; Jacobsen, S.E.; Berger, F.; et al. The histone variant H2A.W and linker histone H1 co-regulate heterochromatin accessibility and DNA methylation. Nat. Commun. 2021, 12, 2683. [Google Scholar] [CrossRef] [PubMed]
- Sabot, F.; Schulman, A.H. Parasitism and the retrotransposon life cycle in plants: A hitchhiker’s guide to the genome. Heredity 2006, 97, 381–388. [Google Scholar] [CrossRef] [Green Version]
- Blumenstiel, J.P. Birth, School, Work, Death, and Resurrection: The Life Stages and Dynamics of Transposable Element Proliferation. Genes 2019, 10, 336. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, I.; Sarazin, A.; Bowler, C.; Colot, V.; Quesneville, H. Genome-wide evidence for local DNA methylation spreading from small RNA-targeted sequences in Arabidopsis. Nucleic Acids Res. 2011, 39, 6919–6931. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Lyons, D.B.; Zilberman, D. Histone H1 prevents non-cg methylation-mediated small RNA biogenesis in arabidopsis heterochromatin. eLife 2021, 10, e72676. [Google Scholar] [CrossRef]
- Colomé Tatché, M.; Cortijo, S.; Wardenaar, R.; Monteiro Morgado, L.; Lahouze, B.; Sarazin, A.; Etcheverry, M.; Martin, A.; Feng, S.; Duvernois-Berthet, E.; et al. Features of the Arabidopsis recombination landscape resulting from the combined loss of sequence variation and DNA methylation. Proc. Natl. Acad. Sci. USA 2012, 109, 16240–16245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirouze, M.; Lieberman-Lazarovich, M.; Aversano, R.; Bucher, E.; Nicolet, J.; Reinders, J.; Paszkowski, J. Loss of DNA methylation affects the recombination landscape in Arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, 5880–5885. [Google Scholar] [CrossRef] [Green Version]
- Si, W.; Yuan, Y.; Huang, J.; Zhang, X.; Zhang, Y.; Zhang, Y.; Tian, D.; Wang, C.; Yang, Y.; Yang, S. Widely distributed hot and cold spots in meiotic recombination as shown by the sequencing of rice F2 plants. New Phytol. 2015, 206, 1491–1502. [Google Scholar] [CrossRef] [PubMed]
- Nosaka, M.; Itoh, J.-I.; Nagato, Y.; Ono, A.; Ishiwata, A.; Sato, Y. Role of Transposon-Derived Small RNAs in the Interplay between Genomes and Parasitic DNA in Rice. PLoS Genet. 2012, 8, e1002953. [Google Scholar] [CrossRef] [PubMed]
- Slotkin, R.K.; Vaughn, M.; Borges, F.; Tanurdžić, M.; Becker, J.D.; Feijó, J.A.; Martienssen, R.A. Epigenetic Reprogramming and Small RNA Silencing of Transposable Elements in Pollen. Cell 2009, 136, 461–472. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.-H.; Fang, Y.-Y.; Duan, C.-G.; Fang, R.-X.; Ding, S.-W.; Guo, H.-S. Genome-wide identification of endogenous RNA-directed DNA methylation loci associated with abundant 21-nucleotide siRNAs in Arabidopsis. Sci. Rep. 2016, 6, 36247. [Google Scholar] [CrossRef] [Green Version]
- Martienssen, R.A. Maintenance of heterochromatin by RNA interference of tandem repeats. Nat. Genet. 2003, 35, 213–214. [Google Scholar] [CrossRef]
- Hardcastle, T.J.; Lewsey, M.G. Mobile small RNAs and their role in regulating cytosine methylation of DNA. RNA Biol. 2016, 13, 1060–1067. [Google Scholar] [CrossRef] [Green Version]
- Cutter, A.R.; Hayes, J.J. A brief review of nucleosome structure. FEBS Lett. 2015, 589, 2914–2922. [Google Scholar] [CrossRef] [Green Version]
- Fyodorov, D.V.; Zhou, B.-R.; Skoultchi, A.I.; Bai, Y. Emerging roles of linker histones in regulating chromatin structure and function. Nat. Rev. Mol. Cell Biol. 2018, 19, 192–206. [Google Scholar] [CrossRef]
- Lei, B.; Capella, M.; Montgomery, S.A.; Borg, M.; Osakabe, A.; Goiser, M.; Muhammad, A.; Braun, S.; Berger, F. A Synthetic Approach to Reconstruct the Evolutionary and Functional Innovations of the Plant Histone Variant H2A.W. Curr. Biol. 2021, 31, 182–191.e185. [Google Scholar] [CrossRef] [PubMed]
- Buschbeck, M.; Hake, S.B. Variants of core histones and their roles in cell fate decisions, development and cancer. Nat. Rev. Mol. Cell Biol. 2017, 18, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Magaña-Acosta, M.; Valadez-Graham, V. Chromatin Remodelers in the 3D Nuclear Compartment. Front. Genet. 2020, 11, 600615. [Google Scholar] [CrossRef] [PubMed]
- Ryan, D.P.; Owen-Hughes, T. Snf2-family proteins: Chromatin remodellers for any occasion. Curr. Opin. Chem. Biol. 2011, 15, 649–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischle, W.; Wang, Y.; Allis, C.D. Histone and chromatin cross-talk. Curr. Opin. Cell Biol. 2003, 15, 172–183. [Google Scholar] [CrossRef] [Green Version]
- Huettel, B.; Kanno, T.; Daxinger, L.; Aufsatz, W.; Matzke, A.J.M.; Matzke, M. Endogenous targets of RNA-directed DNA methylation and Pol IV in Arabidopsis. EMBO J. 2006, 25, 2828–2836. [Google Scholar] [CrossRef] [Green Version]
- Böhmdorfer, G.; Rowley, M.J.; Kuciński, J.; Zhu, Y.; Amies, I.; Wierzbicki, A.T. RNA-directed DNA methylation requires stepwise binding of silencing factors to long non-coding RNA. Plant J. Cell Mol. Biol. 2014, 79, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Wongpalee, S.P.; Liu, S.; Gallego-Bartolomé, J.; Leitner, A.; Aebersold, R.; Liu, W.; Yen, L.; Nohales, M.A.; Kuo, P.H.; Vashisht, A.A.; et al. CryoEM structures of Arabidopsis DDR complexes involved in RNA-directed DNA methylation. Nat. Commun. 2019, 10, 3916. [Google Scholar] [CrossRef] [Green Version]
- Zhong, X.; Hale, C.J.; Law, J.A.; Johnson, L.M.; Feng, S.; Tu, A.; Jacobsen, S.E. DDR complex facilitates global association of RNA polymerase v to promoters and evolutionarily young transposons. Nat. Struct. Mol. Biol. 2012, 19, 870–875. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Lyons, D.B.; Kim, M.Y.; Moore, J.D.; Zilberman, D. DNA Methylation and Histone H1 Jointly Repress Transposable Elements and Aberrant Intragenic Transcripts. Mol. Cell 2020, 77, 310–323.e317. [Google Scholar] [CrossRef]
- Sharifi-Zarchi, A.; Gerovska, D.; Adachi, K.; Totonchi, M.; Pezeshk, H.; Taft, R.J.; Schöler, H.R.; Chitsaz, H.; Sadeghi, M.; Baharvand, H.; et al. DNA methylation regulates discrimination of enhancers from promoters through a H3K4me1-H3K4me3 seesaw mechanism. BMC Genom. 2017, 18, 964. [Google Scholar] [CrossRef] [PubMed]
- Gendrel, A.-V. Dependence of Heterochromatic Histone H3 Methylation Patterns on the Arabidopsis Gene DDM1. Science 2002, 297, 1871–1874. [Google Scholar] [CrossRef] [PubMed]
- Bhadouriya, S.L.; Mehrotra, S.; Basantani, M.K.; Loake, G.J.; Mehrotra, R. Role of Chromatin Architecture in Plant Stress Responses: An Update. Front. Plant Sci. 2021, 11, 603380. [Google Scholar] [CrossRef] [PubMed]
- Talbert, P.B.; Henikoff, S. Remodeller–variant tag team slams transposons. Nat. Cell Biol. 2021, 23, 297–298. [Google Scholar] [CrossRef]
- Tao, Y.; Xi, S.; Shan, J.; Maunakea, A.; Che, A.; Briones, V.; Lee, E.Y.; Geiman, T.; Huang, J.; Stephens, R.; et al. Lsh, chromatin remodeling family member, modulates genome-wide cytosine methylation patterns at nonrepeat sequences. Proc. Natl. Acad. Sci. USA 2011, 108, 5626–5631. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Loci | Induced Defect | Other Genes Involved | Reference |
|---|---|---|---|
| bal | Dwarfing | NBS-LRR-class disease-resistance gene cluster | [29] |
| fwa | Late flowering | SINE (Short interspersed nuclear element) in the FWA promoter region | [24,33] |
| bonsai | Dwarfing Late flowering | BONSAI gene flanked by long interspersed nuclear element (LINE) | [30] |
| superman (sup) | Abnormal flower development | DECREASED DNA METHYLATION 2 (DDM2) | [34,35] |
| agamous (ag) | Abnormal flower development | DECREASED DNA METHYLATION 2 (DDM2) | [34] |
| RdDM | DDM1 | |
|---|---|---|
| In both animals and plants | Unique to plants [12,66]. | In both, with a DDM1 homologue in mammals [11,23,39,67] |
| Heterosis and embryo lethality | RdDM mutants are viable and can reproduce even when inbred with no significant effects on plant heterosis [12,19]. | Yes, DDM1 mutation may possess lethal effects on the embryo, which is severe in double mutants. Inbred ddm1 mutants generally have reduced heterosis [25,37,68]. |
| Participates in PTGS or RNAi pathways | In all plants [12,69]. | Limited participation in all plants [70,71]. |
| sRNA-dependent activity | Uses microRNA precursors, 21–22nt, 24nt small RNAs [12]. | sRNA-independent [72]. |
| de novo DNA methylation | Yes, using 21–22nt siRNAs [12]. | Not involved in de novo DNA methylation [12]. |
| Maintenance DNA methylation | Yes, in all sequence contexts [12]. | Yes, in all sequence contexts [22]. |
| Silenced TEs | Short TEs, the edges of long TEs, euchromatic TEs and TE relics [11]. | The bodies of long TEs, heterochromatic TEs, protein-coding TEs and mobile TEs [11]. |
| Dominance | Euchromatin [11,73]. | Heterochromatin [11,22]. |
| Gene body silencing | Common [12]. | Rare [22]. |
| Pol IV activity | Yes [12]. | No [12]. |
| Argonaute and Dicer activity | Yes, AGO1, AGO4, AGO6 and AGO9. Complete or partial Dicer activity with DCL1, DCL2, DCL3 and DCL4 depending on the small RNA type, RdDM component involved and target loci [12,74,75]. | No [12]. |
| Nucleosome displacement | No, but involved in DNA methylation [12]. | Yes, involved in DNA methylation, histone methylation, nucleosome positioning and other chromatin modifications [11,22,76]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akinmusola, R.Y.; Wilkins, C.-A.; Doughty, J. DDM1-Mediated TE Silencing in Plants. Plants 2023, 12, 437. https://doi.org/10.3390/plants12030437
Akinmusola RY, Wilkins C-A, Doughty J. DDM1-Mediated TE Silencing in Plants. Plants. 2023; 12(3):437. https://doi.org/10.3390/plants12030437
Chicago/Turabian StyleAkinmusola, Ruth Y., Catherine-Axa Wilkins, and James Doughty. 2023. "DDM1-Mediated TE Silencing in Plants" Plants 12, no. 3: 437. https://doi.org/10.3390/plants12030437