1. Introduction
Bottle gourd (
Lagenaria siceraria (Molina) Standl) is an edible, useful, and medicinal vegetable crop belonging to the Cucurbitaceae family, valued for its culinary, therapeutic, and useful properties [
1]. This cross-pollinated vegetable exhibits varying degrees of pollination, ranging from 94% to 99%. The degree of cross-pollination depends on several factors, such as the flowering time, temperature, wind velocity and direction, planting design, insect population, and genotypic nature, ultimately determining the kernel quality [
2,
3,
4]. The reproductive phase in cucurbits, initiated approximately six to seven weeks after planting, is critical in deciding the seed quality due to the source–sink relationship dynamics [
5,
6]. As a result of the source–sink strength relationship, a variation in seed lots can be observed [
7]. Being indeterminate, bottle gourd vines can withstand adverse weather conditions like continuous dry and overcast weather such as seed germination, emergence, seedling establishment, leaf canopy development, flowering, and seed maturation.
Seed quality is paramount in agricultural production and food security, especially during the growing uncertainty caused by climate change [
8,
9,
10] and abiotic factors [
11,
12]. Hybrids, carrying the combined genetic makeup of plants, are pivotal in agricultural biotechnology and crop improvement, with high-quality seeds being in constant demand to ensure productive plantations. Farmers constantly need high-quality sources to ensure efficient and effective plantations; thus, companies must sell high-quality seeds to maintain their competitive position in markets [
13]. Numerous abiotic factors, predominantly high and low temperatures, low light, moisture surpluses [
9,
13,
14], and deficits impose limitations on the growth, development, and yield of different crops [
15,
16]. Cool nights are favorable during the fruiting period, but extreme or low temperatures may result in delayed growth and aborted fruiting sites. Temperature significantly affects leaf expansion, phenology, internode elongation, assimilation partitioning, and biomass production [
17]. The variations in protein and lipid contents were attributed to several environmental factors [
16,
17,
18]. The timing of seed development is crucial, as it directly impacts the accumulation of essential minerals and nutrients necessary for robust seedling growth [
18,
19].
The mineral content of seeds correlates with seedling quality. During seed development, there is a critical need to accumulate minerals and essential nutrients, particularly in the early growth stage [
19,
20]. The minerals collected during seed ontogeny are the primary factors influencing seedling development; growth would be delayed without these components [
20]. To understand the mechanism behind the source–sink relationship and improve the nutritional profiles of bottle gourd crops, it becomes crucial to examine the interactions between macro- and micronutrients during various stages of cross-breeding [
21]. To maintain the crop’s nutritional profile, seeds contain enough metabolic reserves to facilitate seedling establishment during seed development. Carbohydrates, lipids, and proteins interact synergistically to create an optimal environment for seed germination and seedling development. To develop bottle gourd genotypes with improved nutritional and phytochemical compositions, micronutrient and macronutrient profiling of the germplasm is essential [
22]. Before delving into the study of gene activity influencing the inheritance of nutritional and antinutritional traits in both parent and offspring plants, it is imperative to identify the optimal cross-breeding periods for the desired genotypes.
Quality is pivotal in crop production, necessitating specific attributes and functionality during the developmental phase [
4]. The efficiency of hybrid seed production depends on multiple factors, such as selecting an appropriate agro-climate location, a suitable season, improved floral synchronization for enhanced seed setting through suitable crossing periods, supplementary pollination techniques, etc. [
4]. Ref. [
4] found that bottle gourd exhibits bi-hemispheric adaptability and distribution, thriving in tropical and temperate regions. It flourishes in areas with annual rainfall ranging from 400 to 1500 mm, with a preference for moderate soil moisture levels over excessive ones for optimal harvests. According to Grubben and Dento [
23], bottle gourd grows well at temperatures of 25–35 °C. The optimum germination temperature is between 20 and 25 °C. Temperatures below 15 °C and above 35 °C reduce the germination rate [
24]. Agronomic practices that promote the production of more female flowers than male flowers could increase yields; however, Haque et al. [
25] observed fewer seed sets due to the reduction in pollen. Therefore, it is essential to determine the optimum ratio of male and female flowers to optimize the fruit and seed set.
Nevertheless, the effects of varying environmental conditions on bottle gourd seed properties remain inadequately understood, underscoring the importance of studying mineral accumulation during different fruit-setting periods at various temperatures. Thus, the present study aims to gain a deeper understanding and clarify the impact of pollination vulnerability and the outcomes resulting from varying cross-breeding timings on the biochemical composition of bottle gourd.
2. Materials and Methods
2.1. Plant Material and Experimental Setup
The research experiment was conducted at the experimental farm of Seed Science & Technology (29.1416° N, 75.7112° E, with an average elevation of 215 m (705 ft) above mean sea level) during the Kharif (2017) and summer season (2018) in a randomized block design. The soil type of the experimental field is sandy loam. The concentrations of organic matter percent, total nitrogen, available phosphate, and rapidly available phosphate in the uppermost 20–30 cm soil were 0.49%, 182 kg/ha, 18 kg/ha, and 285 kg/ha, respectively. The climate is semi-arid, with freezing winters and hot, dry, desiccating winds during the summer.
The seeds of bottle gourd parental lines, G-2 (Male Line) and Pusa Naveen (Female Line), were procured from the Department of Vegetable Science to produce Hybrid HBGH-35. For the production of hybrid HBGH-35, seeds of the bottle gourd parental lines G-2 (male line) and Pusa Naveen (female line) were obtained from the Department of Vegetable Science. HBGH-35 is the first hybrid developed by the CCS HAU, Hisar, India, using the G2 male line from Gujarat regions, and the female lines used was PUSA Naveen developed by IARI Pusa, both of which are popularly grown in the northern part of India. G2 is an early flowering line, and Pusa Naveen is a high-yielding line. These lines were selected to compare the potential of HBGH-35 concerning parental lines.
Being an indeterminate crop, flowering in bottle gourd continues for around two months. Sowing occurred on 17 July, and flowering began in both parents in the first week of September. There were ten vines per plot with a size of 6 × 3 m, and the vine-to-vine spacing was 60 cm. Every experiment was carried out in three biological replicates and five technical triplicates. All methods followed the protocols based on the relevant guidelines and regulations. A standard recommended package of practices (POP) was followed throughout the experiment.
2.2. Pollination and Emasculation
The flowers (male and female) were emasculated one day before opening, and they were wrapped in a thin layer of cotton. The following day, pollen was collected from the male flower and dusted on the female flower, which was then wrapped with a thin layer of cotton until the fruit formation started.
Monoecious bottle gourds (Lagenaria siceraria) bear male and female flowers. Key inferior ovary variations between male and female flowers were as follows:
The male flower has a long, slender peduncle and one pollen-producing stamen. Male flowers lack or have a primitive ovary. Male bottle-gourd flowers lack or have a primitive ovary. Female flowers have a unique ovary at the base that will produce fruit if fertilized. The bulbous ovary is at the floral center. Well-developed female flowers have ovules in their ovaries. A functioning ovary is essential for female flowers. Bottle gourds and other monoecious plants generate more male flowers than female flowers. This may be an adaptive technique to ensure pollination with enough pollen. Environmental conditions and plant health affect the flower output and ratio. To determine flower sex and optimize pollination, growers watch the flowering pattern. If natural pollination is insufficient, farmers may hand-pollinate to increase the fruit set.
After bagging, we waited for one week and observed female flower ovary growth. The increased size of the ovary confirmed the successful crossing after it was tagged using different color threads, as shown in the picture. The pollinated female flower was tagged for further identification.
The emasculation and pollination work were carried out in both parental lines with the help of needles, scalpers, and forceps. The two glass cups, cotton, muslin cloth, plastic bag, camel brush, and scissors were used to collect pollen from male flowers and dust it on the stigmatic surface of emasculated female flowers for hybrid seed production. Manual emasculation and dusting were continued throughout the crossing period. Subsequently, we pinched off all the floral buds emerging beyond 15 days after the crossing period to ensure better growth and production of crossed fruits (
Figure 1). The harvesting dates of different crossing periods are listed in
Table 1.
At the start of the crossing period in hybrid and parental line seed production, the male flower was chosen in the parental line, bagged with paper bags the previous evening, pinched off the following day, and kept carefully in a glass container. The fruits were harvested from the hybrid and parental lines at maturity when the color turned from green to brown. Seeds were extracted manually by slicing the ripened fruits into small pieces, and seeds were separated by squeezing the fruit pulp. The extracted seeds were cleaned with fresh water and dried to lower the moisture content.
2.3. Soil Profile
The concentrations of organic matter percent, total nitrogen, available phosphate, and rapidly available phosphate in the uppermost 20–30 cm soil were 0.49%, 182 kg/ha, 18 kg/ha, and 285 kg/ha, respectively.
2.4. Agro-Meteorological Data
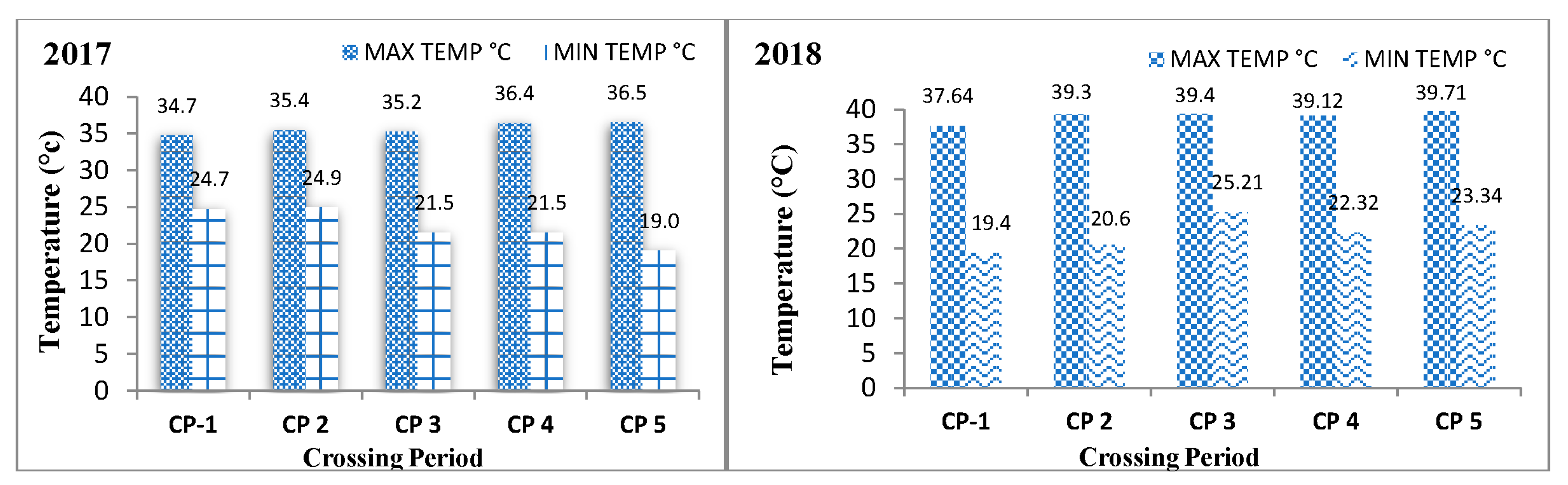
The harvested seeds of five crossing periods of all genotypes were studied for various biochemical parameters under varying weather conditions. Agro-meteorological data were recorded at the agro-meteorological observatory during the seed development period of both seasons and converted to weekly data, presented in
Table 2 and
Figure 2.
2.5. Biochemical Analysis
The seeds were extracted from fruits set in each crossing period manually. After extraction, seeds are dried in the shade to lower the moisture content to 8% and be stored for further biochemical analysis. The moisture content of the seed was measured using the hot oven method. In this method, the pre-weighed aluminum boxes and seed material are placed in an oven maintained at 103 °C. Seeds are dried at this temperature for 1 hr. The relative humidity of the ambient air in the laboratory must be less than 70 percent when the moisture determination is carried out.
The moisture content as a percentage by weight (fresh weight basis) is calculated to one decimal place by using the following formulae:
M1 = Weighing container weight with cover in grams
M2 = Weighing container weight with cover and seeds before drying
M3 = After-drying weight of the weighing container with cover and seeds
Total soluble solids (°Brix) were determined by using a hand refractometer. The seeds’ ash, moisture content, and crude fiber content were determined using the recommended method [
26]. The estimated total nitrogen (N) and crude protein were calculated by multiplying 4.27 by the N concentration [
26]. The lipid content was determined using the Soxhlet method. Total carbohydrate was estimated as the method described by Dubois et al. [
27]. Vitamin C was measured using the titration method of reduction of 2,6-dichlorophenol-indophenol (DCPIP) dye described [
28]. The energy value was calculated by multiplying the mean values of crude protein, lipid content, and total carbohydrate by factors of 4, 9, and 4, respectively.
2.6. Determination of Macro and Micro-Elements
The mature seeds (50) were dried at constant temperature, then ground to fine powder. Random sampling was performed to ensure that the samples represented the entire mature dried bottle gourd seed population. The measurements of macro- and microelements in dried seeds were carried out using energy dispersive X-ray (EDX) analysis and atomic absorption spectrometry (AAS). Three biological and five technical replicates from each sample were used for analysis.
2.7. Energy Dispersive X-ray Analysis (EDX)
Three seeds from each selection were cryo-fractured in liquid nitrogen and split into halves using a scalpel blade. Seeds were mounted on stubs and secured using double-sided insulating carbon tape. Seed mineral composition was determined separately for the embryo, cotyledon, and seed coat regions.
2.8. Atomic Absorption Spectrometry
The contents of the elements were determined in the seed (2 to 3 fruits per vine) materials after incineration in a muffle furnace at 660 °C. The resultant ash content was solubilized on porcelain crucibles using a 10 mL acid mixture (HNO
3 and HCl mixed in a 1:3 ratio) [
29] using an atomic absorption spectrophotometer (Alpha 4 Model). A rapid sequential absorption spectrometer was used to evaluate potassium using flame atomic emission spectroscopy (FAES) (Varian AA280FS). Flame atomic absorption spectroscopy was used to determine the concentrations of Ca, Mg, Zn, Cu, Fe, and Mn (FAAS).
Table 3 summarizes the components evaluated at each wavelength, recovery level, and relative standard deviation (RSD). Using a Varian Alpha UV-VIS spectrometer at 400 nm, the amount of phosphorus was measured (Spectronic Unicam, Berlin, Germany) [
30].
2.9. Measurement of Germination Axis
2.9.1. Standard Germination
One hundred seeds were taken in four replications from each treatment and placed uniformly between two sufficiently moist germination papers. They were then rolled and placed in a germinator, where the temperature and relative humidity were kept at 25.1 °C and 95.1%, respectively. The final counts were made on the 14th day of the germination test for normal seedlings and expressed in percentages as per the ISTA protocol [
31].
The germination percentage was worked out using the following formula:
2.9.2. Biomass Accumulation
The plants were uprooted and thoroughly washed with distilled water to eliminate the soil. To remove surface moisture, a paper towel was used. An electronic balance (Presica 105A) was used to measure fresh weights. Weighed fresh samples were heated in an air oven at 70 °C for 24 h and then dried to a constant weight at 35 °C. The plant dry weight was equal to the shoot and root dry weights. The root-shoot ratio was calculated according to Agren and Ingestad [
32].
2.9.3. Root and Shoot Length
A two-factorial experiment was replicated three times to measure the germinant axis and provide experimental units. Sand particles were removed from seedlings developed through sand soil by uprooting and washing them. The lengths of the roots and shoots were measured using computer-assisted image analysis. A tabloid scanner (Hewlett Packard Scanjet 4c/t; Palo Alto, CA, USA) was used to capture digital photos at a scanning resolution of 250 dpi. The lengths of the roots and shoots were measured using image software analysis.
2.10. Statistical Analysis
The data were presented as mean SD, and (three replicates)-way ANOVA was conducted to check the significance of the main effects (cultivars and temperature seasons) and their interaction on growth indices, followed by a post hoc comparison (Tuckey’s test) at a 5% level (p < 0.05). Statistical analysis was performed using SPSS v25.0 software (SPSS for Windows, Chicago, IL, USA). The correlations between phytochemicals and weather variables were statistically evaluated and indicated by Pearson’s coefficient indexes using two-tailed bivariate correlates analysis; p < 0.05 was considered statistically significant.
4. Discussion
The estimation of the proximate composition in bottle gourd seeds is essential to determine the source–sink relationship. The relatively low moisture content in bottle gourd is advantageous since the high moisture content is associated with increased bacterial action during storage and is a crucial determinant of seed quality [
35]. The results of this study are very similar to those of [
36], which stated that the moisture content is essential in determining the seed quality. The effect of the moisture content during seed production in bottle gourds is a much more complicated process because the seed development occurs within the moist fruits for several weeks. Seeds are often held in a wet state before their final harvest. The ash content represents the index of mineral elements present in the seed. This indicates that the whole seed may have a higher mineral content [
37] and be of good quality. The ash content in whole seeds is generally higher than the 3.7% reported as the ash content of the calabash seed [
37,
38]. Hassan et al. [
37] observed that ash content represents an index of high minerals that improved the overall performance of seed quality. Dietary fiber promotes the food absorption and bowel movement through the small intestine [
39]. Bottle gourd is a rich source of dietary fibers that expand food inside the colon wall, easing the passage of waste and making it an effective anti-constipation agent. The crude fiber content of the bottle gourd seed is comparable with that of calabash (3.2%) and watermelon (2.46%) [
40].
Total soluble solids (TSS) measure the sugar content of seeds and play a significant role during seed production. The seeds harvested in the fourth week of the crossing period with a temperature of 31.7 °C had a higher protein content (8.2–19.1%), while with the temperature rise (40 °C), the protein content decreased (6.2–17.6%) significantly due to the inactivation of the enzymes involved in protein synthesis [
41]. The protein forms the principal component of embryonic development; thus, this could be why the higher protein concentration in Hybrid HBGH-35 (19.1%) at CP-4 represents high-quality seeds [
42]. Similar results were found [
43] in soybean and in groundnuts [
44]. Lipids represent the energy content required during seed germination. A similar trend was observed for the lipid content (
Table 5). All the cultivars showed significant variations (
p < 0.05) among them. However, the values are lower than the 41–46.8% reported in the same Lagenaria species [
45]. Carbohydrates are a rich source of energy [
10,
46]. The results conform well with those observed in bottle gourd [
37], watermelon [
47], oil bean seed [
48], pumpkin seed [
49], and African walnut [
50], which confirmed that seeds possessing higher carbohydrates have a high germination rate and seed emergence. The results are well documented by several researchers [
51,
52,
53,
54,
55], who reported that temperature plays a crucial role in the accumulation of vitamin C. A higher vitamin C accumulation indicates a higher antioxidant potential of the seed [
51].
Results for EDX are semi-qualitative estimations based on conventional analysis and theoretical intensity adjustments [
56]. It has been reported that phytin globoids are prevalent in the periplasm of embryonic cells in legume seeds, and also that P, Mg, and K accumulate in the globoids. Calcium was scarcely found in the embryonic tissues of bottle gourd seeds. Similar observations were previously reported [
57,
58]. In the seed embryonic region, Zn, Cu, and Mn percentages associated with the P percentage suggested a potential relationship with phytate. Iwai et al. [
59] reported that specific distribution patterns of minerals in seeds depend upon the cell type. However, this report was based on a single spot analysis, which involves analyzing areas’ electron beam size. Phytate synthesis in seeds deposits phosphates and cations for a sufficient period during germination [
55]. According to the results of the present study, seeds may be a significant source of nutrients for catering to the requirements of seedling growth [
60]. A high seed P content improves plant establishment and increases the dry matter accumulation. This results from a faster initial root growth, which gives seedlings earlier access to growth-limiting resources (water and mineral elements). According to Grant et al. [
61], seed P reserves sustain a maximal growth of seedlings for several weeks after germination until the plant has three or more leaves and a substantial root system. Indole acetic acid (IAA)-induced cell wall weakening and solute aggregation inside the cell to establish internal osmotic potential are essential for cell expansion [
62]. K+ buildup lowers the water potential and maintains the pH of growing cells. Since a stoichiometric K+ inflow electrochemically counterbalances the IAA-stimulated H+ outflow, the IAA-induced elongation quickly decreases and eventually stops when K+ is absent. A direct correlation between the seed K content and shoot and root elongation may thus appear.
Sodium (Na) is an element associated with potassium concentration, maintaining body fluid homeostasis [
63]. The high amount of potassium is a characteristic feature of most plant foods [
46,
64,
65]. A high potassium content in relation to the sodium content is thought to benefit hypertensive individuals [
66]. Moreover, the recommended dietary allowance (RDA) for Na and K is 200 and 500 mg, respectively [
33], indicating that bottle gourd seeds may be regarded as a rich source of these minerals for the body. Phosphorous (P) and calcium (Ca) are associated with each other and are required for the growth and maintenance of teeth, bones, and muscles [
34,
67,
68]. An adult’s RDA of Ca and P is 810 mg/day [
51]. This confirms that bottle-gourd seeds are good sources of phosphorous [
69,
70,
71]. The amount of Mg in bottle gourd seeds is higher than in groundnut, shea nut, cotton seed, and calabash seed [
67,
72]. Hence, bottle gourd seeds may thus be a rich source of Mg, assuming a total absorption by the body and ignoring the effect of antinutritional factors [
33]. Iron (Fe) is the most abundant microelement in crop seeds. A similar trend was observed for the Fe content, with no significant differences (
p > 0.05). The values in [
64,
65,
73] well support the results. Copper (Cu) is an essential trace element and an integral part of ceruloplasmin (a Cu protein) and cytochrome (an electron carrier). It plays a vital role in energy metabolism [
42,
74]. High temperatures disrupt Fe homeostasis and inactivate enzymes. The values obtained in the present study seem high compared with the pride of Barbados [
66] and calabash seeds [
72]. The temperature was directly influenced by zinc (Zn) and manganese (Mn) accumulation in bottle-gourd cultivars. Manganese plays a structural role in the chloroplast membrane and is a cofactor for DNA, RNA, and fatty acid synthesis [
75], while Zn is required for growth, tissue repair, and the normal functioning of the immune system [
76].
Minerals play a pivotal role in plant nutrition and development [
77]. The utilization of mineral components in agriculture, as advocated by Ayurveda, has been a longstanding practice in various traditional farming systems [
78,
79]. Ayurveda suggests that incorporating mineral-rich preparations into the soil can enrich it with these crucial elements, ensuring that bottle-gourd plants receive adequate mineral nutrition throughout their growth cycle [
80]. Ayurvedic practices involve using herbal formulations combined with mineral components, which acts as a carrier for other nutrients, facilitating their absorption by plants [
81]. This synergistic approach harnesses the benefits of both herbal and mineral components to promote plant growth and health of bottle gourd.
The results of the germination indices reveal interesting insights into the germination performance of different genotypes and crossing periods in bottle gourd (
Lagenaria siceraria) cultivation. This finding aligns with previous studies indicating that environmental factors during seed maturation, such as temperature and moisture, can influence the seed quality and germination rates [
82]. The Kharif season typically provides more favorable conditions for seed development, potentially contributing to the observed difference in germination percentages. Genotypic variation in germination capacity is well-documented in crop plants [
83]. Genetic factors, including seed coat characteristics and embryo vigor, can significantly impact germination rates.
Interestingly, the crossing period also played a crucial role in seed germination. The lowest germination percentage was recorded in seeds produced from crossing period CP1. This result may be attributed to factors such as parental genetic compatibility and seed maturation conditions during that specific crossing period. Subsequent crossing periods, CP2 through CP4, showed a significant increase in the germination percentage, with the maximum germination observed in CP4. This pattern suggests that certain crossing periods may be more conducive to achieving higher germination rates, which could be linked to optimal environmental conditions during seed development [
84].
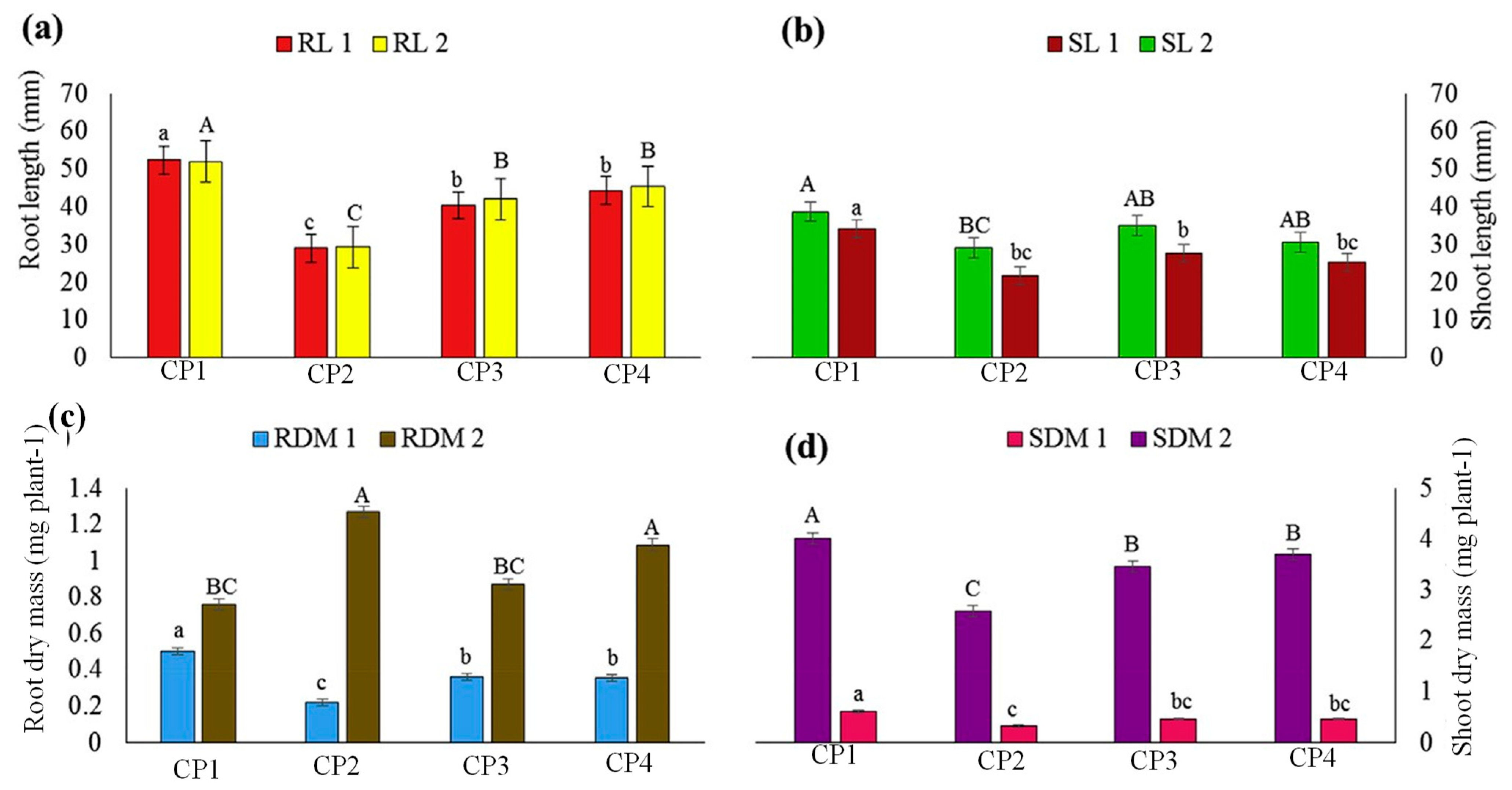
The growth characteristics of hybrid plants derived from the HBGH-35 and G2 parental lines provide additional insights into the impact of the genotype and crossing period on plant development. The significant differences observed in root length, shoot length, root dry mass, and shoot dry mass between HBGH-35 and G2 reaffirm the role of genetic variation in determining plant growth parameters [
85]. At CP4, HBGH-35 exhibited the highest root length, suggesting its potential for developing a robust root system, which can be advantageous for nutrient uptake and overall plant stability [
86]. Similarly, the variation in shoot length and biomass measurements further emphasizes the genotype-specific responses to crossing periods. The highest shoot length recorded for HBGH-35 at CP1 and CP3 suggests its ability to produce longer and potentially more photosynthetically active shoots under certain conditions [
87]. On the other hand, G2 consistently exhibited a lower shoot length and biomass measurements, indicating its comparatively inferior growth characteristics.
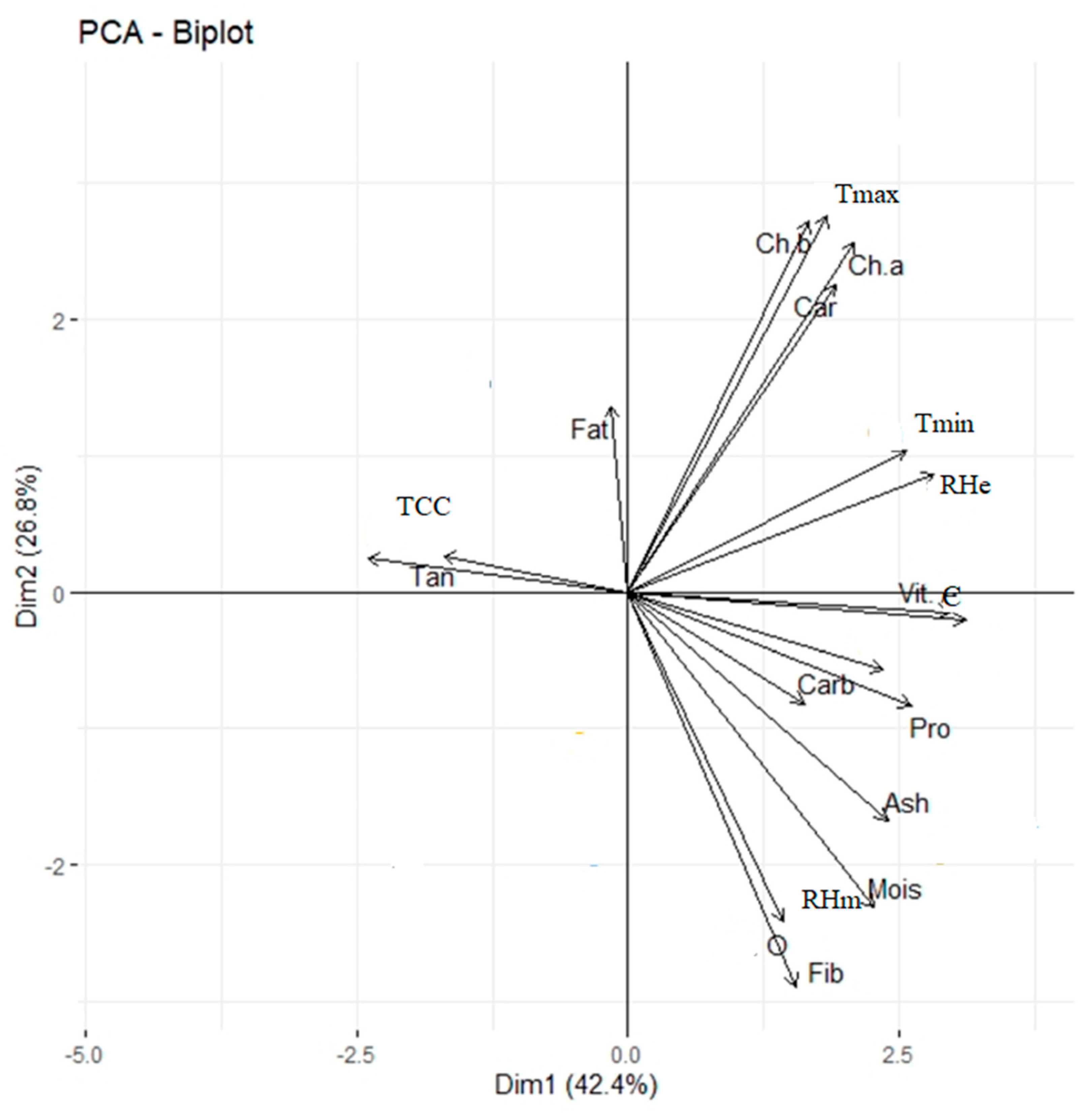
Correlation studies provide a better understanding of the association of different traits that may be useful to breeders in selecting cultivars possessing groups of desired agronomical traits [
88,
89]. Similar results were reported in previous studies [
90,
91,
92,
93]. Introducing high-yield cultivars depends upon the agro-climatic conditions and phenological traits and choosing a small number of essential traits with a positive correlation [
46,
94]. Henceforth, the results showed that all the biochemical components were significantly positively correlated to the minimum temperature and morning and evening relative humidity, suggesting their relationship with temperature and solar radiation.


 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}



