

Phlomidis Radix Extract Alleviates Paclitaxel-Induced Neuropathic Pain by Modulating Spinal TRPV1 in Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Phlomidis Radix (P.Radix) Extract Preparation
2.3. Paclitaxel, P.Radix, and Sesamoside Administration
2.4. Behavioral Tests
2.5. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.6. Western Blot
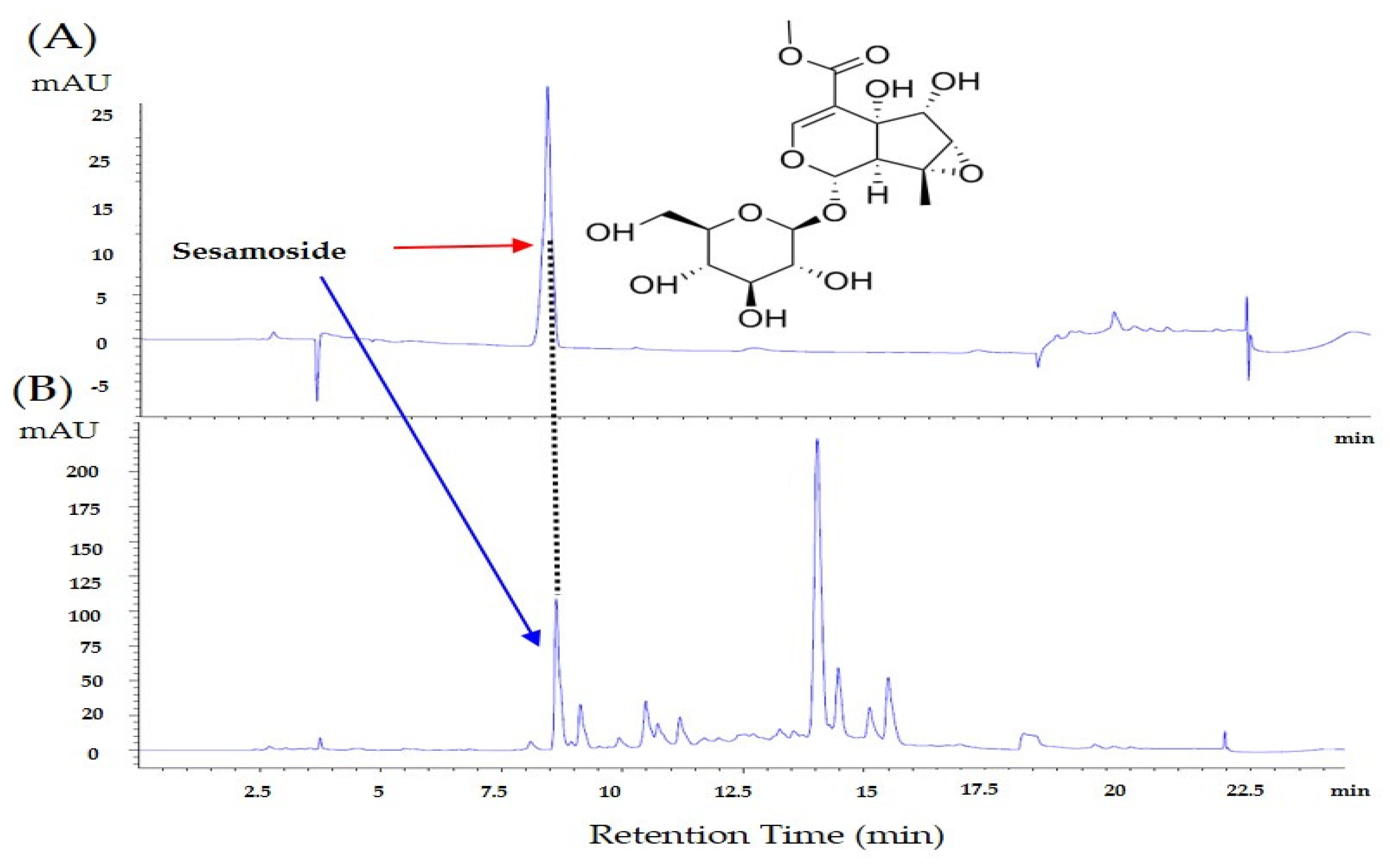
2.7. Identification and Quantification of Sesamoside in P.Radix Extract
3. Results
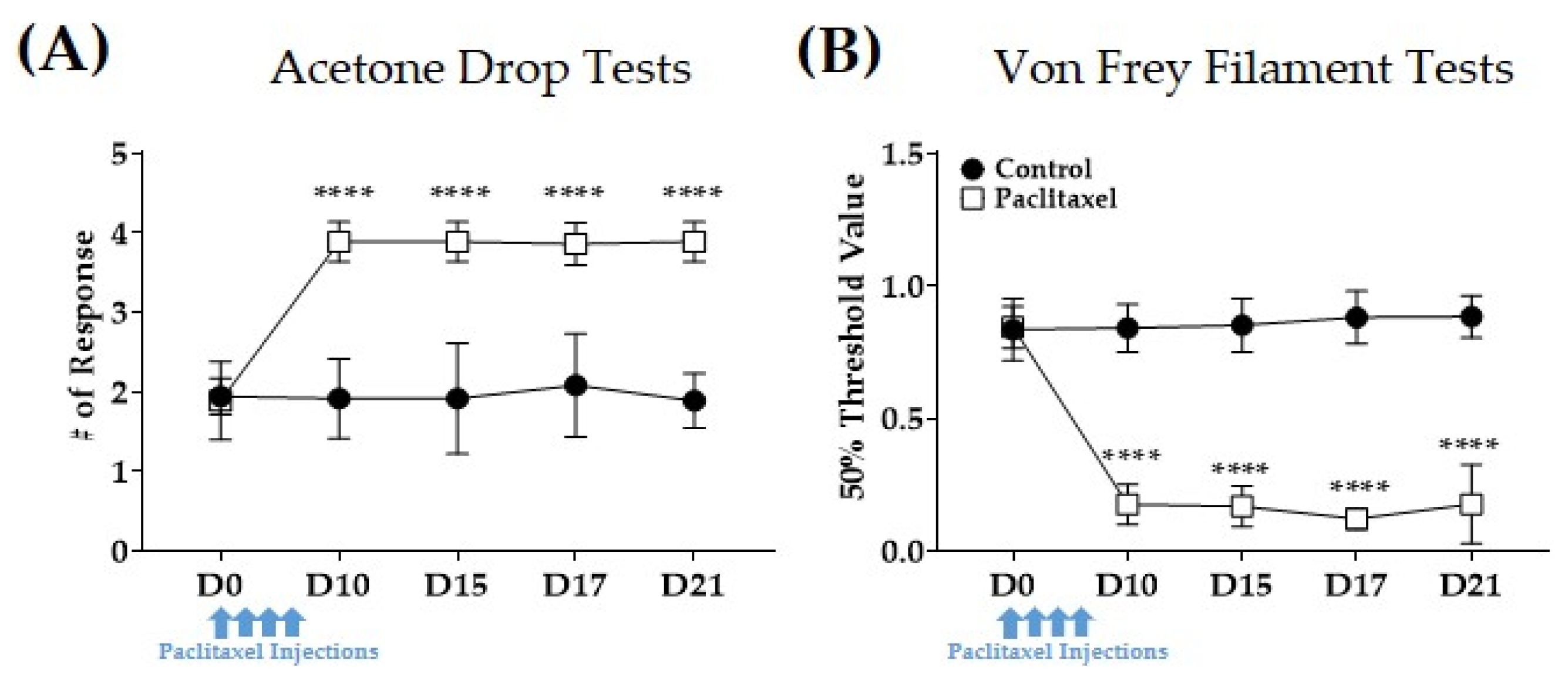
3.1. Multiple Paclitaxel Injections Induce Increased Responses to Cold and Mechanical Stimuli in Mice
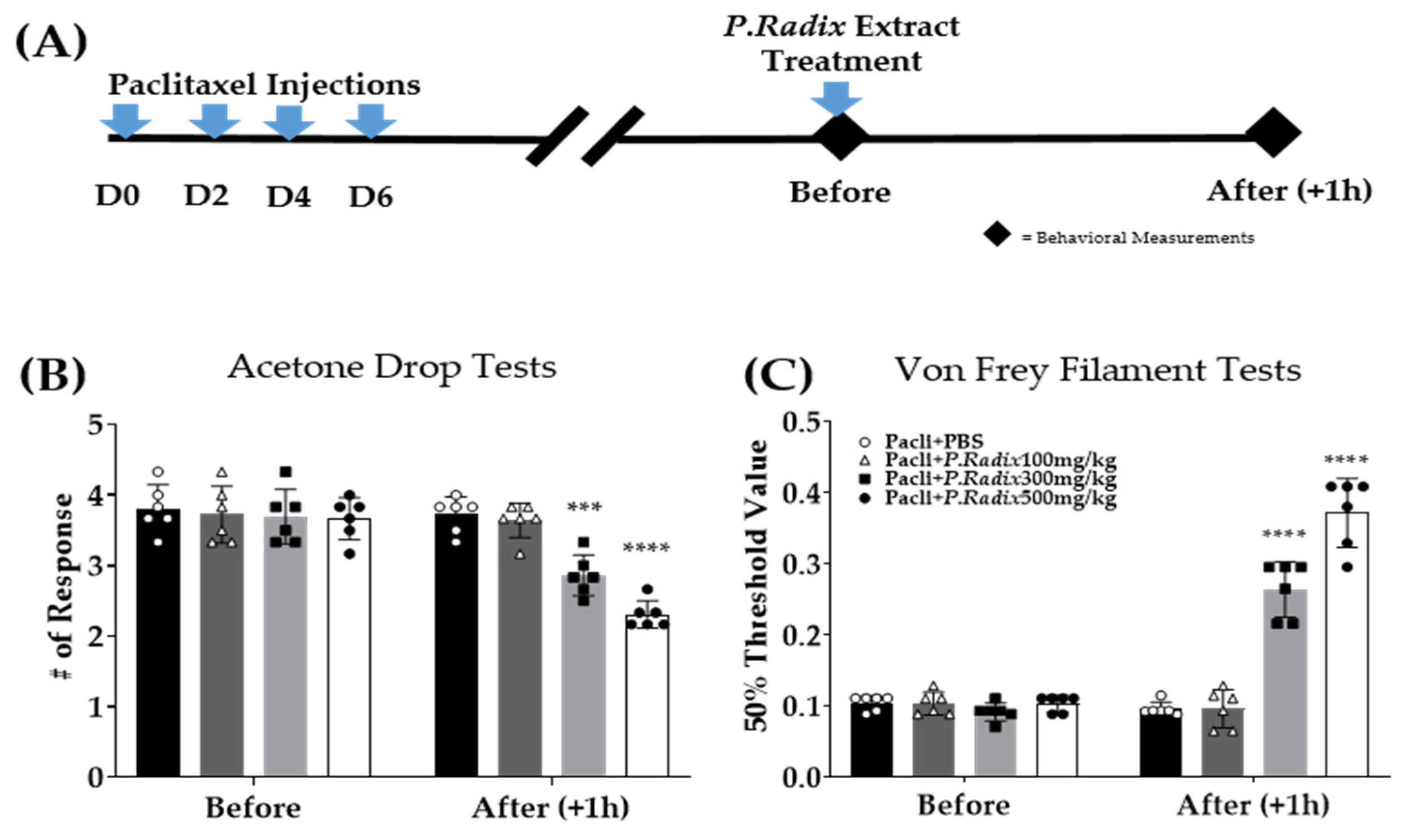
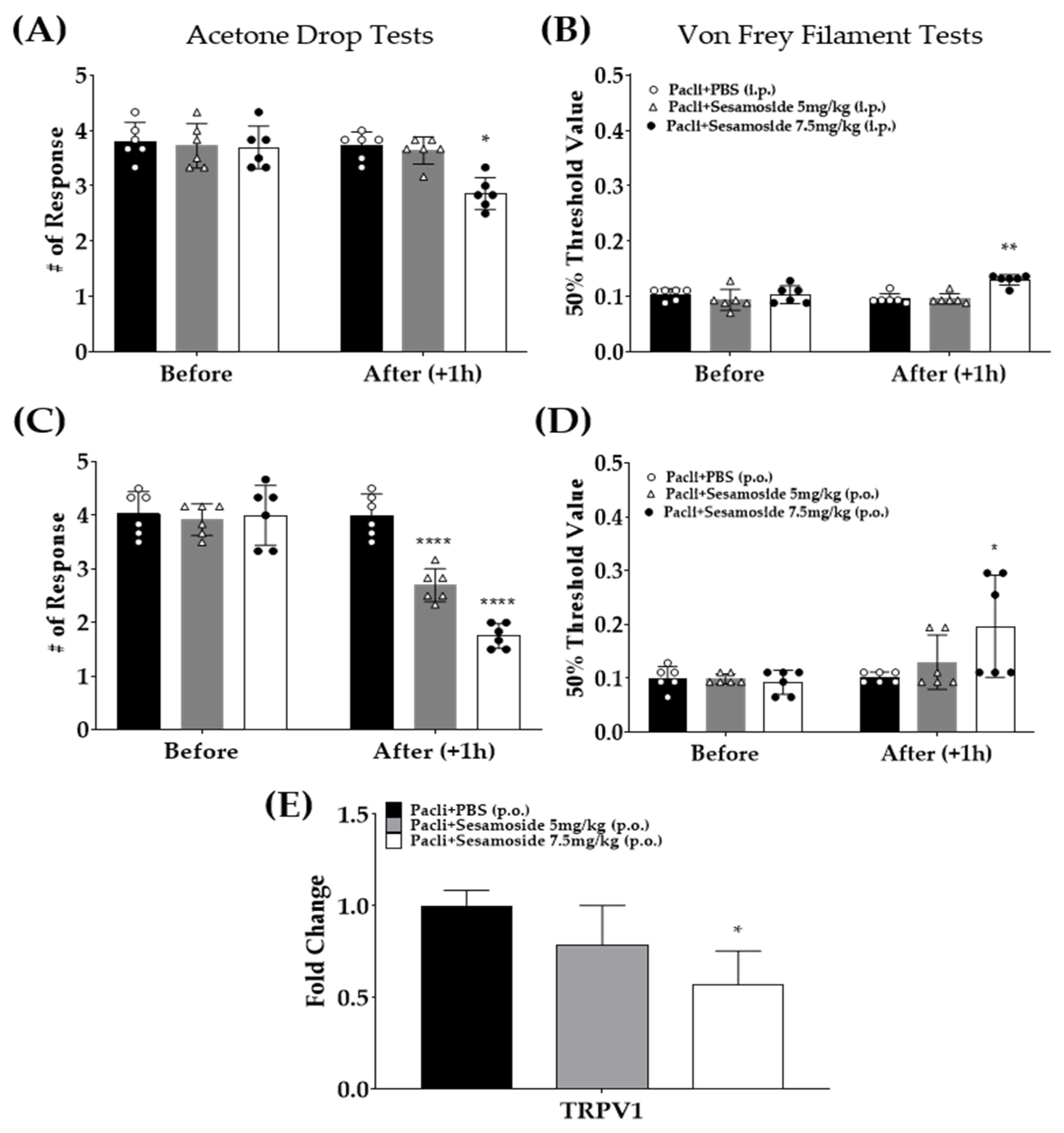
3.2. Analgesic Effect of Single Administration of Phlomidis Radix Extract in Cold and Mechanical Peripheral Neuropathy-Induced by Paclitaxel
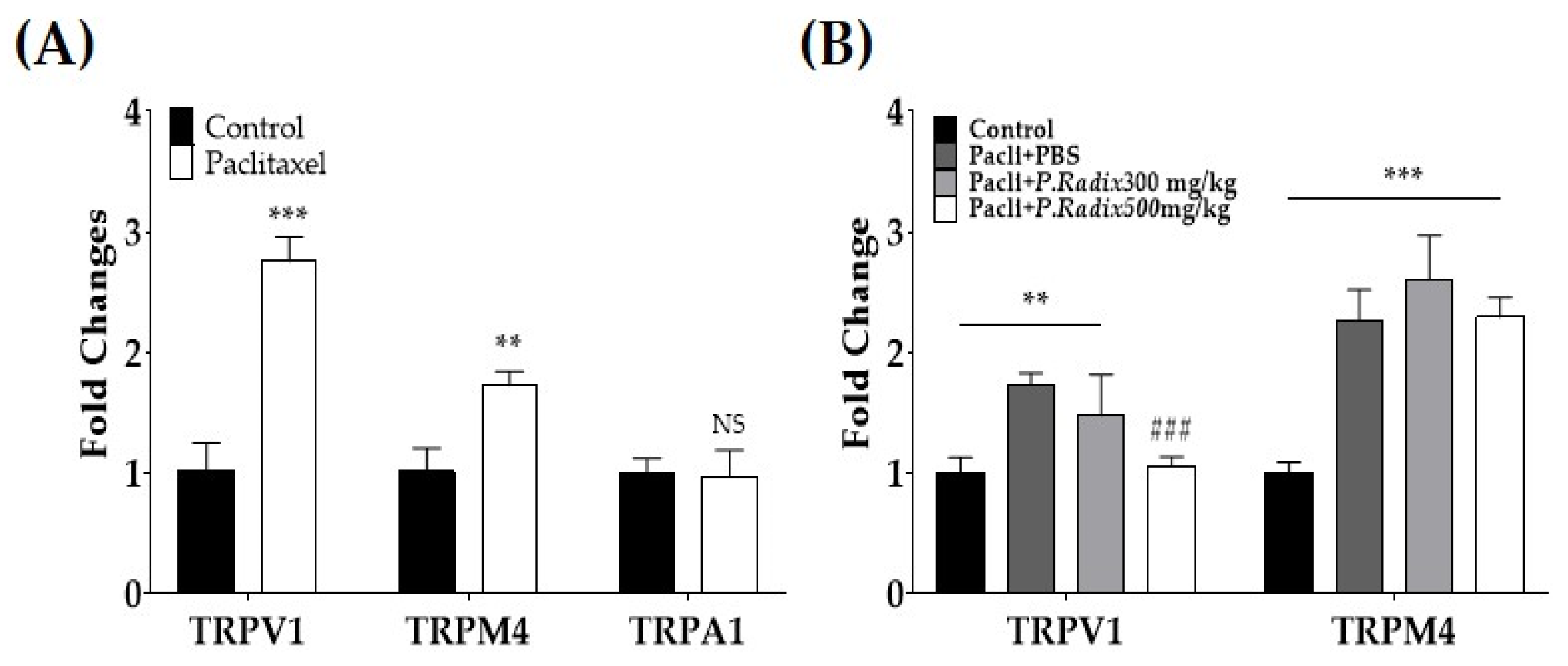
3.3. Gene Expression of TRPV1 and TRPM4, but Not TPRA1, Are Downregulated after Multiple Paclitaxel Injections
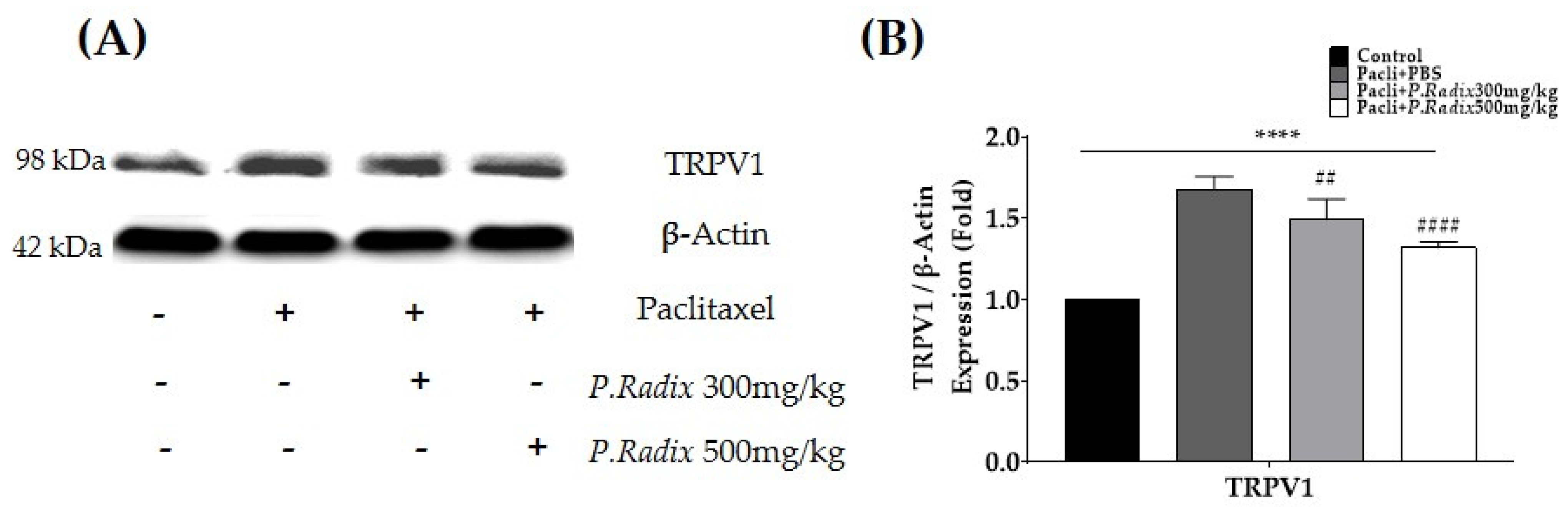
3.4. The Protein Expression of Spinal TRPV1 Is Downregulated after P.Radix Extract Treatment
3.5. Identification and Quantification of Sesamoside in P.Radix Extract by Using High-Performance Liquid Chromatography (HPLC)
3.6. Sesamoside Alleviates Paclitaxel-Induced Cold and Mechanical Hypersensitivity in Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bernabeu, E.; Cagel, M.; Lagomarsino, E.; Moretton, M.; Chiappetta, D.A. Paclitaxel: What has been done and the challenges remain ahead. Int. J. Pharm. 2017, 526, 474–495. [Google Scholar]
- Alves, R.C.; Fernandes, R.P.; Eloy, J.O.; Salgado, H.R.N.; Chorilli, M. Characteristics, properties and analytical methods of paclitaxel: A review. Crit. Rev. Anal. Chem. 2018, 48, 110–118. [Google Scholar] [CrossRef]
- Yan-Hua, Y.; Jia-Wang, M.; Xiao-Li, T. Research progress on the source, production, and anti-cancer mechanisms of paclitaxel. Chin. J. Nat. Med. 2020, 18, 890–897. [Google Scholar]
- Foland, T.B.; Dentler, W.L.; Suprenant, K.A.; Gupta, M.L., Jr.; Himes, R.H. Paclitaxel-induced microtubule stabilization causes mitotic block and apoptotic-like cell death in a paclitaxel-sensitive strain of Saccharomyces cerevisiae. Yeast 2005, 22, 971–978. [Google Scholar] [CrossRef] [PubMed]
- Al-Mahayri, Z.N.; AlAhmad, M.M.; Ali, B.R. Current opinion on the pharmacogenomics of paclitaxel-induced toxicity. Expert Opin. Drug Metab. Toxicol. 2021, 17, 785–801. [Google Scholar] [CrossRef]
- Klein, I.; Lehmann, H.C. Pathomechanisms of paclitaxel-induced peripheral neuropathy. Toxics 2021, 9, 229. [Google Scholar] [CrossRef]
- Goode, D.J.; Whitaker, E.E.; Mecum, N.E. Ovariectomy increases paclitaxel-induced mechanical hypersensitivity and reduces anti-inflammatory CD4+ T cells in the dorsal root ganglion of female mice. J. Neuroimmunol. 2022, 367, 577878. [Google Scholar]
- Montell, C.; Rubin, G.M. Molecular characterization of the Drosophila trp locus: A putative integral membrane protein required for phototransduction. Neuron 1989, 2, 1313–1323. [Google Scholar] [CrossRef] [PubMed]
- Sousa-Valente, J.; Andreou, A.P.; Urban, L.; Nagy, I. Transient receptor potential ion channels in primary sensory neurons as targets for novel analgesics. Br. J. Pharmacol. 2014, 171, 2508–2527. [Google Scholar] [CrossRef]
- Gees, M.; Colsoul, B.; Nilius, B. The role of transient receptor potential cation channels in Ca2+ signaling. Cold Spring Harb. Perspect Biol. 2010, 2, a003962. [Google Scholar] [CrossRef]
- Caterina, M.J.; Leffler, A.; Malmberg, A.B.; Martin, W.J.; Trafton, J.; Petersen-Zeitz, K.R.; Koltzenburg, M.; Basbaum, A.I.; Julius, D. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 2000, 288, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.B.; Gray, J.; Gunthorpe, M.J.; Hatcher, J.P.; Davey, P.T.; Overend, P.; Harries, M.H.; Latcham, J.; Clapham, C.; Atkinson, K.; et al. Vanilloid receptor-1 is essential for inflammatory thermal hyperalgesia. Nature 2000, 405, 183–187. [Google Scholar] [CrossRef]
- Hasan, R.; Zhang, X. Ca(2+) Regulation of TRP Ion Channels. Int. J. Mol. Sci. 2018, 19, 1256. [Google Scholar] [CrossRef]
- Jara-Oseguera, A.; Simon, S.A.; Rosenbaum, T. TRPV1: On the road to pain relief. Curr. Mol. Pharmacol. 2008, 1, 255–269. [Google Scholar] [CrossRef]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef]
- Caterina, M.J.; Julius, D. The vanilloid receptor: A molecular gateway to the pain pathway. Annu. Rev. Neurosci. 2001, 24, 487–517. [Google Scholar] [CrossRef]
- Tominaga, M.; Caterina, M.J.; Malmberg, A.B.; Rosen, T.A.; Gilbert, H.; Skinner, K.; Raumann, B.E.; Basbaum, A.I.; Julius, D. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron 1998, 21, 531–543. [Google Scholar] [CrossRef]
- Tominaga, M.; Tominaga, T. Structure and function of TRPV1. Pflügers Arch. 2005, 451, 143–150. [Google Scholar] [CrossRef]
- Nilius, B.; Voets, T. TRP channels: A TR (I) P through a world of multifunctional cation channels. Pflügers Arch. 2005, 451, 1–10. [Google Scholar] [CrossRef]
- Moran, M.M.; Xu, H.; Clapham, D.E. TRP ion channels in the nervous system. Curr. Opin. Neurobiol. 2004, 14, 362–369. [Google Scholar] [CrossRef]
- Iftinca, M.; Defaye, M.; Altier, C. TRPV1-targeted drugs in development for human pain conditions. Drugs 2021, 81, 7–27. [Google Scholar] [CrossRef]
- Cortright, D.N.; Krause, J.E.; Broom, D.C. TRP channels and pain. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2007, 1772, 978–988. [Google Scholar] [CrossRef]
- Szallasi, A.; Cortright, D.N.; Blum, C.A.; Eid, S.R. The vanilloid receptor TRPV1: 10 years from channel cloning to antagonist proof-of-concept. Nat. Rev. Drug Discov. 2007, 6, 357–372. [Google Scholar] [CrossRef]
- Levine, J.D.; Alessandri-Haber, N. TRP channels: Targets for the relief of pain. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2007, 1772, 989–1003. [Google Scholar] [CrossRef]
- Launay, P.; Fleig, A.; Perraud, A.-L.; Scharenberg, A.M.; Penner, R.; Kinet, J.-P. TRPM4 Is a Ca2+-Activated Nonselective Cation Channel Mediating Cell Membrane Depolarization. Cell 2002, 109, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Talavera, K.; Yasumatsu, K.; Voets, T.; Droogmans, G.; Shigemura, N.; Ninomiya, Y.; Margolskee, R.F.; Nilius, B. Heat activation of TRPM5 underlies thermal sensitivity of sweet taste. Nature 2005, 438, 1022–1025. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Droogmans, G.; Wondergem, R. Transient receptor potential channels in endothelium: Solving the calcium entry puzzle? Endothelium 2003, 10, 5–15. [Google Scholar] [CrossRef]
- McKemy, D.D. How cold is it? TRPM8 and TRPA1 in the molecular logic of cold sensation. Mol. Pain 2005, 1, 16. [Google Scholar] [CrossRef]
- Souza Monteiro de Araujo, D.; Nassini, R.; Geppetti, P.; De Logu, F. TRPA1 as a therapeutic target for nociceptive pain. Expert Opin. Ther. Targets 2020, 24, 997–1008. [Google Scholar] [CrossRef]
- Koivisto, A.; Jalava, N.; Bratty, R.; Pertovaara, A. TRPA1 antagonists for pain relief. Pharmaceuticals 2018, 11, 117. [Google Scholar] [CrossRef] [PubMed]
- Bandell, M.; Story, G.M.; Hwang, S.W.; Viswanath, V.; Eid, S.R.; Petrus, M.J.; Earley, T.J.; Patapoutian, A. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron 2004, 41, 849–857. [Google Scholar] [CrossRef] [PubMed]
- Park, I.; Yang, S.; Kim, W.J.; Noh, P.; Lee, H.O.; Moon, B.C. Authentication of herbal medicines Dipsacus asper and Phlomoides umbrosa using DNA barcodes, chloroplast genome, and sequence characterized amplified region (SCAR) marker. Molecules 2018, 23, 1748. [Google Scholar] [CrossRef]
- Korea Institute of Oriental Medicine. Defining Dictionary for Medicinal Herbs [Korean,‘Hanyak Giwon Sajeon’]. 2016. Available online: http://boncho.kiom.re.kr/codex/ (accessed on 9 October 2023).
- Chun, J.M.; Lee, A.Y.; Nam, J.Y.; Lee, M.Y.; Choe, M.S.; Lim, K.S.; Kim, C.; Kim, J.S. Protective effects of Phlomis umbrosa extract on a monosodium iodoacetate-induced osteoarthritis model and prediction of molecular mechanisms using transcriptomics. Phytomedicine 2021, 81, 153429. [Google Scholar] [CrossRef]
- Shang, X.; Wang, J.; Li, M.; Miao, X.; Pan, H.; Yang, Y.; Wang, Y. Antinociceptive and anti-inflammatory activities of Phlomis umbrosa Turcz extract. Fitoterapia 2011, 82, 716–721. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-J.; Choi, H.-I.; Kim, Y.-C.; You, H.-K.; Shin, H.-S. Effects of dichloromethane fraction of Phlomidis radix on bone formation in human fetal osteoblasts. J. Korean Acad. Periodontol. 2003, 33, 259–269. [Google Scholar] [CrossRef]
- Lee, D.; Kim, Y.S.; Song, J.; Kim, H.S.; Lee, H.J.; Guo, H.; Kim, H. Effects of Phlomis umbrosa Root on Longitudinal Bone Growth Rate in Adolescent Female Rats. Molecules 2016, 21, 461. [Google Scholar] [CrossRef]
- Lee, J.H.; Ji, H.; Ko, S.G.; Kim, W. JI017 Attenuates Oxaliplatin-Induced Cold Allodynia via Spinal TRPV1 and Astrocytes Inhibition in Mice. Int. J. Mol. Sci. 2021, 22, 8811. [Google Scholar] [CrossRef]
- Lee, J.H.; Min, D.; Lee, D.; Kim, W. Zingiber officinale Roscoe Rhizomes Attenuate Oxaliplatin-Induced Neuropathic Pain in Mice. Molecules 2021, 26, 548. [Google Scholar] [CrossRef]
- Dixon, W.J. Efficient analysis of experimental observations. Annu. Rev. Pharmacol. Toxicol. 1980, 20, 441–462. [Google Scholar] [CrossRef]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.; Chung, J.; Yaksh, T. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Park, K.-T.; Kim, S.; Choi, I.; Han, I.-H.; Bae, H.; Kim, W. The involvement of the noradrenergic system in the antinociceptive effect of cucurbitacin D on mice with paclitaxel-induced neuropathic pain. Front. Pharmacol. 2023, 13, 1055264. [Google Scholar] [CrossRef] [PubMed]
- Mielke, S.; Sparreboom, A.; Mross, K. Peripheral neuropathy: A persisting challenge in paclitaxel-based regimes. Eur. J. Cancer 2006, 42, 24–30. [Google Scholar] [CrossRef]
- Cavaletti, G. Chemotherapy-induced peripheral neurotoxicity (CIPN): What we need and what we know. J. Peripher. Nerv. Syst. 2014, 19, 66–76. [Google Scholar] [CrossRef]
- Kim, M.J.; Lee, J.H.; Jang, J.U.; Quan, F.S.; Kim, S.K.; Kim, W. The efficacy of combination treatment of gabapentin and electro-acupuncture on paclitaxel-induced neuropathic pain. Korean J. Physiol. Pharmacol. 2017, 21, 657–666. [Google Scholar] [CrossRef]
- Flatters, S.J.; Bennett, G.J. Studies of peripheral sensory nerves in paclitaxel-induced painful peripheral neuropathy: Evidence for mitochondrial dysfunction. Pain 2006, 122, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S. Chemotherapy-induced peripheral neuropathy and rehabilitation: A review. Semin Oncol. 2021, 48, 193–207. [Google Scholar] [CrossRef]
- Staff, N.P.; Fehrenbacher, J.C.; Caillaud, M.; Damaj, M.I.; Segal, R.A.; Rieger, S. Pathogenesis of paclitaxel-induced peripheral neuropathy: A current review of in vitro and in vivo findings using rodent and human model systems. Exp. Neurol. 2020, 324, 113121. [Google Scholar] [CrossRef]
- Klein, I.; Boenert, J.; Lange, F.; Christensen, B.; Wassermann, M.K.; Wiesen, M.H.; Olschewski, D.N.; Rabenstein, M.; Müller, C.; Lehmann, H.C. Glia from the central and peripheral nervous system are differentially affected by paclitaxel chemotherapy via modulating their neuroinflammatory and neuroregenerative properties. Front. Pharmacol. 2022, 13, 1038285. [Google Scholar] [CrossRef]
- Cata, J.P.; Weng, H.R.; Chen, J.H.; Dougherty, P.M. Altered discharges of spinal wide dynamic range neurons and down-regulation of glutamate transporter expression in rats with paclitaxel-induced hyperalgesia. Neuroscience 2006, 138, 329–338. [Google Scholar] [CrossRef]
- Costa-Pereira, J.T.; Serrão, P.; Martins, I.; Tavares, I. Serotoninergic pain modulation from the rostral ventromedial medulla (RVM) in chemotherapy-induced neuropathy: The role of spinal 5-HT3 receptors. Eur. J. Neurosci. 2020, 51, 1756–1769. [Google Scholar] [CrossRef]
- Yadav, R.; Yan, X.; Maixner, D.W.; Gao, M.; Weng, H.R. Blocking the GABA transporter GAT-1 ameliorates spinal GABAergic disinhibition and neuropathic pain induced by paclitaxel. J. Neurochem. 2015, 133, 857–869. [Google Scholar] [CrossRef]
- Kalynovska, N.; Adamek, P.; Palecek, J. TRPV1 receptors contribute to paclitaxel-induced c-Fos expression in spinal cord dorsal horn neurons. Physiol. Res. 2017, 66, 549. [Google Scholar] [CrossRef]
- Kamata, Y.; Kambe, T.; Chiba, T.; Yamamoto, K.; Kawakami, K.; Abe, K.; Taguchi, K. Paclitaxel induces upregulation of transient receptor potential vanilloid 1 expression in the rat spinal cord. Int. J. Mol. Sci. 2020, 21, 4341. [Google Scholar] [CrossRef]
- Haraguchi, K.; Kawamoto, A.; Isami, K.; Maeda, S.; Kusano, A.; Asakura, K.; Shirakawa, H.; Mori, Y.; Nakagawa, T.; Kaneko, S. TRPM2 contributes to inflammatory and neuropathic pain through the aggravation of pronociceptive inflammatory responses in mice. J. Neurosci. 2012, 32, 3931–3941. [Google Scholar] [CrossRef]
- Miyake, T.; Shirakawa, H.; Kusano, A.; Sakimoto, S.; Konno, M.; Nakagawa, T.; Mori, Y.; Kaneko, S. TRPM2 contributes to LPS/IFNγ-induced production of nitric oxide via the p38/JNK pathway in microglia. Biochem. Biophys Res. Commun. 2014, 444, 212–217. [Google Scholar] [CrossRef]
- Lee, J.K.; Kim, N.J. Recent Advances in the Inhibition of p38 MAPK as a Potential Strategy for the Treatment of Alzheimer’s Disease. Molecules 2017, 22, 1287. [Google Scholar] [CrossRef]
- Bautista, D.; Julius, D. Fire in the hole: Pore dilation of the capsaicin receptor TRPV1. Nat. Neurosci. 2008, 11, 528–529. [Google Scholar] [CrossRef] [PubMed]
- Gaudet, R. TRP channels entering the structural era. J. Physiol. 2008, 586, 3565–3575. [Google Scholar] [CrossRef] [PubMed]
- Samanta, A.; Hughes, T.E.T.; Moiseenkova-Bell, V.Y. Transient Receptor Potential (TRP) Channels. Subcell Biochem. 2018, 87, 141–165. [Google Scholar] [CrossRef] [PubMed]
- Špicarová, D.; Paleček, J. The role of spinal cord vanilloid (TRPV1) receptors in pain modulation. Physiol. Res. 2008, 57, S69–S77. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Adamek, P.; Zhang, H.; Tatsui, C.E.; Rhines, L.D.; Mrozkova, P.; Li, Q.; Kosturakis, A.K.; Cassidy, R.M.; Harrison, D.S. The cancer chemotherapeutic paclitaxel increases human and rodent sensory neuron responses to TRPV1 by activation of TLR4. J. Neurosci. 2015, 35, 13487–13500. [Google Scholar] [CrossRef] [PubMed]
- Vennekens, R.; Nilius, B. Insights into TRPM4 function, regulation and physiological role. Transient Recept. Potential Channels 2007, 179, 269–285. [Google Scholar]
- Benbow, J.H.; Mann, T.; Keeler, C.; Fan, C.; Hodsdon, M.E.; Lolis, E.; DeGray, B.; Ehrlich, B.E. Inhibition of paclitaxel-induced decreases in calcium signaling. J. Biol. Chem. 2012, 287, 37907–37916. [Google Scholar] [CrossRef]
- Corey, D.P.; García-Añoveros, J.; Holt, J.R.; Kwan, K.Y.; Lin, S.Y.; Vollrath, M.A.; Amalfitano, A.; Cheung, E.L.; Derfler, B.H.; Duggan, A.; et al. TRPA1 is a candidate for the mechanosensitive transduction channel of vertebrate hair cells. Nature 2004, 432, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Kwan, K.Y.; Allchorne, A.J.; Vollrath, M.A.; Christensen, A.P.; Zhang, D.S.; Woolf, C.J.; Corey, D.P. TRPA1 contributes to cold, mechanical, and chemical nociception but is not essential for hair-cell transduction. Neuron 2006, 50, 277–289. [Google Scholar] [CrossRef]
- Ta, L.E.; Bieber, A.J.; Carlton, S.M.; Loprinzi, C.L.; Low, P.A.; Windebank, A.J. Transient Receptor Potential Vanilloid 1 is essential for cisplatin-induced heat hyperalgesia in mice. Mol. Pain 2010, 6, 15. [Google Scholar] [CrossRef]
- World Health Organization. Medicinal Plants in the Republic of Korea: Information on 150 Commonly Used Medicinal Plants; World Health Organization, Regional Office for the Western Pacific: Manila, Philippines, 1998.
- Lee, J.E.; Lee, H.; Kim, M.H.; Yang, W.M. Osteogenic effects of Phlomis umbrosa via up-regulation of Runx2 in osteoporosis. Biomed. Rep. 2019, 10, 17–22. [Google Scholar] [CrossRef]
- Fuji, Y.; Uchida, A.; Fukahori, K.; Chino, M.; Ohtsuki, T.; Matsufuji, H. Chemical characterization and biological activity in young sesame leaves (Sesamum indicum L.) and changes in iridoid and polyphenol content at different growth stages. PLoS ONE 2018, 13, e0194449. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conditions | |||
|---|---|---|---|
| Treatment | Sesamoside | ||
| Column | Ymc-Triart C18 Length: 25 cm Diameter: 0.46 cm Particle Size: 5 μm | ||
| Flow rate | 1.0 mL/min | ||
| Injection volume | 10 μL | ||
| UV detection | 234 nm | ||
| Run time | 25 min | ||
| Time (min) | %ACN | 0.1% Phosphoric acid | Flow (mL/min) |
| 0 | 5 | 95 | 1.0 |
| 15 | 40 | 60 | 1.0 |
| 16 | 100 | 0 | 1.0 |
| 19 | 100 | 0 | 1.0 |
| 20 | 5 | 95 | 1.0 |
| 25 | 5 | 95 | 1.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, K.-T.; Ko, S.-G.; Kim, W. Phlomidis Radix Extract Alleviates Paclitaxel-Induced Neuropathic Pain by Modulating Spinal TRPV1 in Mice. Plants 2023, 12, 3819. https://doi.org/10.3390/plants12223819
Park K-T, Ko S-G, Kim W. Phlomidis Radix Extract Alleviates Paclitaxel-Induced Neuropathic Pain by Modulating Spinal TRPV1 in Mice. Plants. 2023; 12(22):3819. https://doi.org/10.3390/plants12223819
Chicago/Turabian StylePark, Keun-Tae, Seong-Gyu Ko, and Woojin Kim. 2023. "Phlomidis Radix Extract Alleviates Paclitaxel-Induced Neuropathic Pain by Modulating Spinal TRPV1 in Mice" Plants 12, no. 22: 3819. https://doi.org/10.3390/plants12223819