
Influence of the Growing Region on the Phytochemical Composition and Antioxidant Properties of North American Cranberry Fruit (Vaccinium macrocarpon Aiton)

Abstract
:1. Introduction
2. Results and Discussion
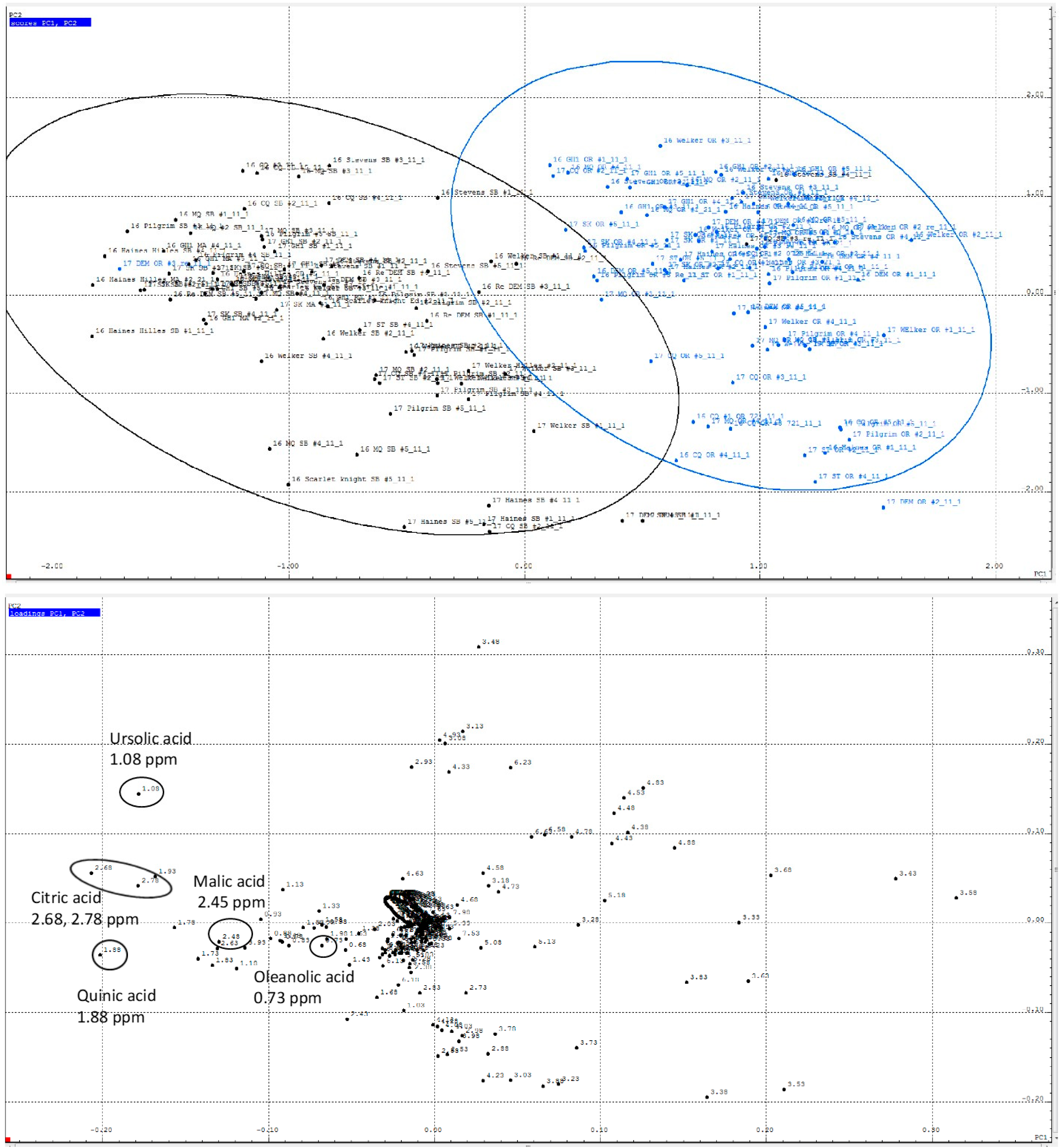
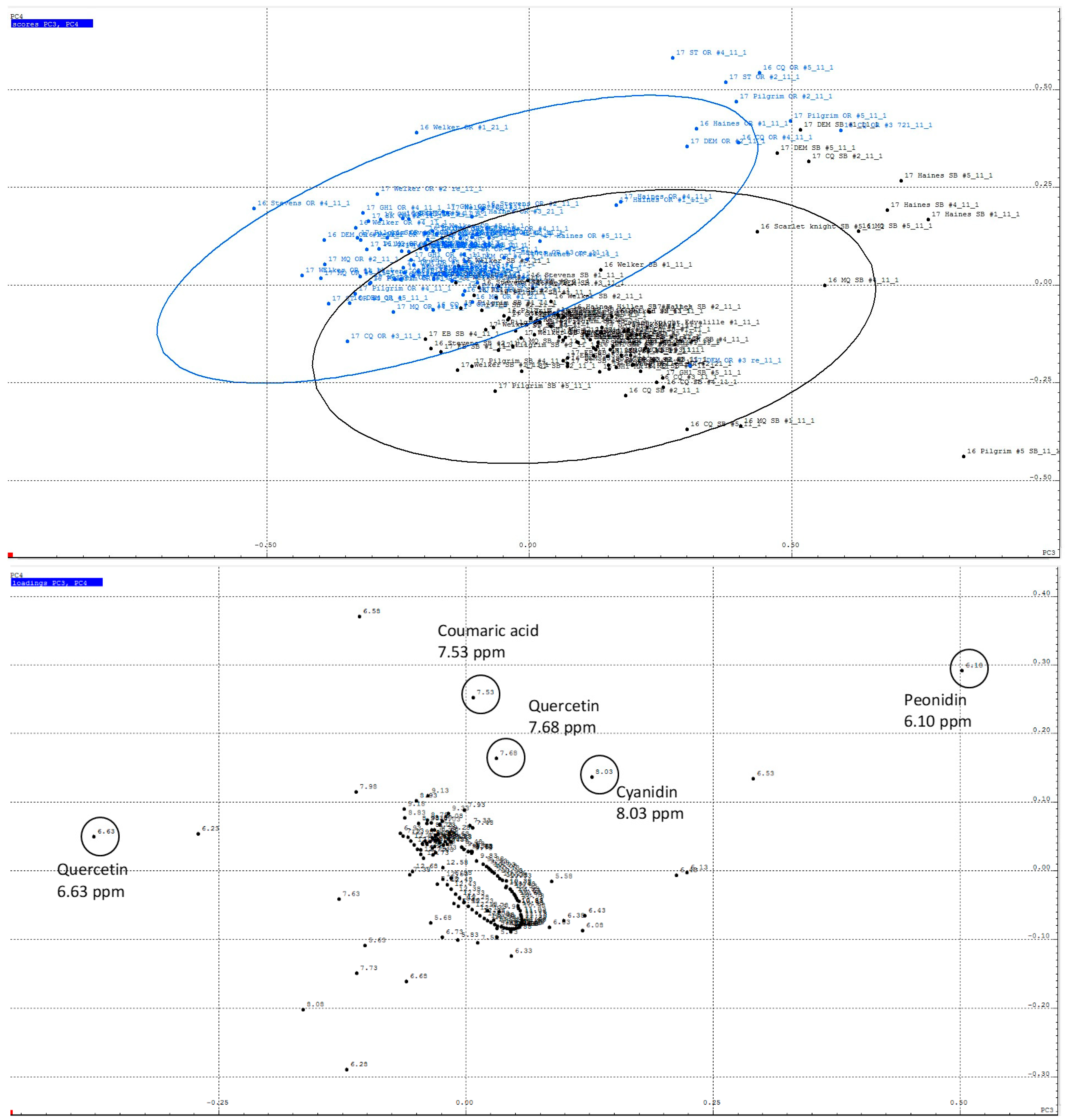
2.1. Non-Targeted PCA of Cranberry 1H NMR Reveals Regional Differences
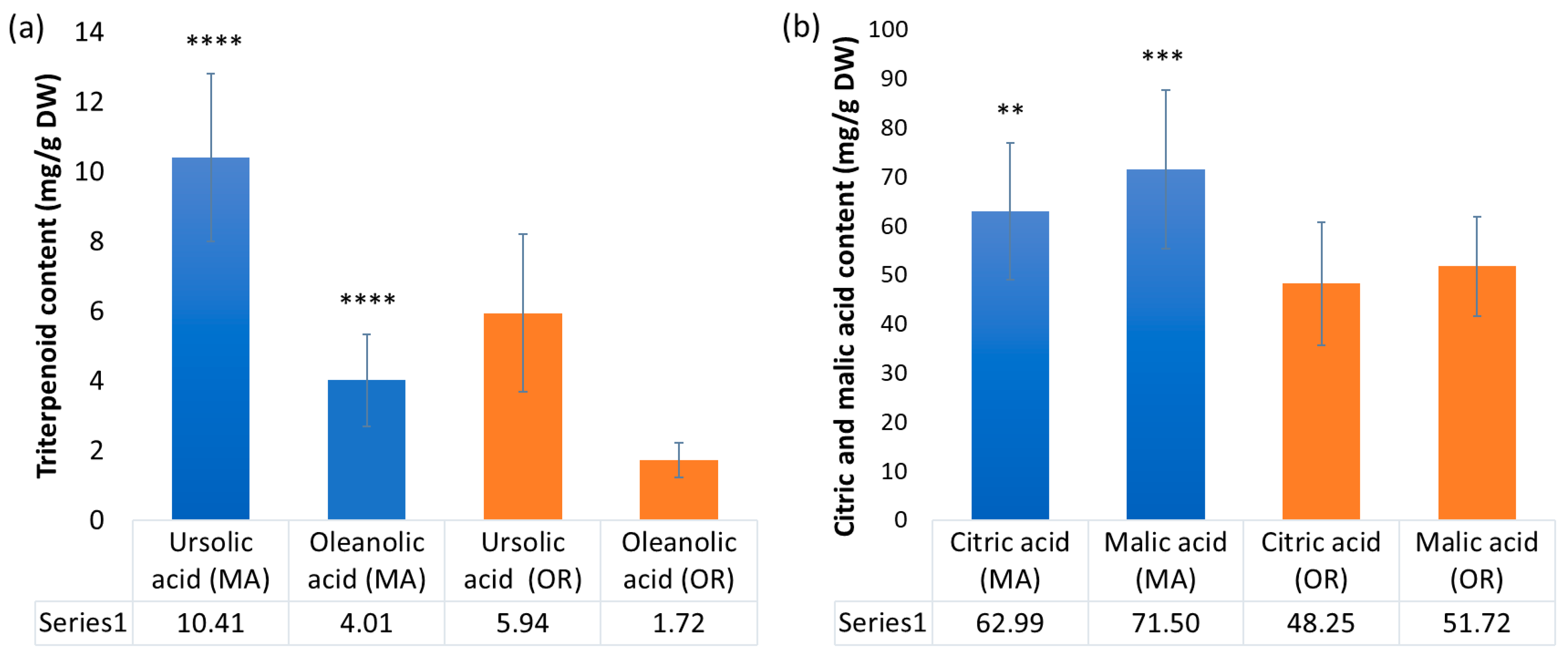
2.2. Triterpenoids and Organic Acids Content
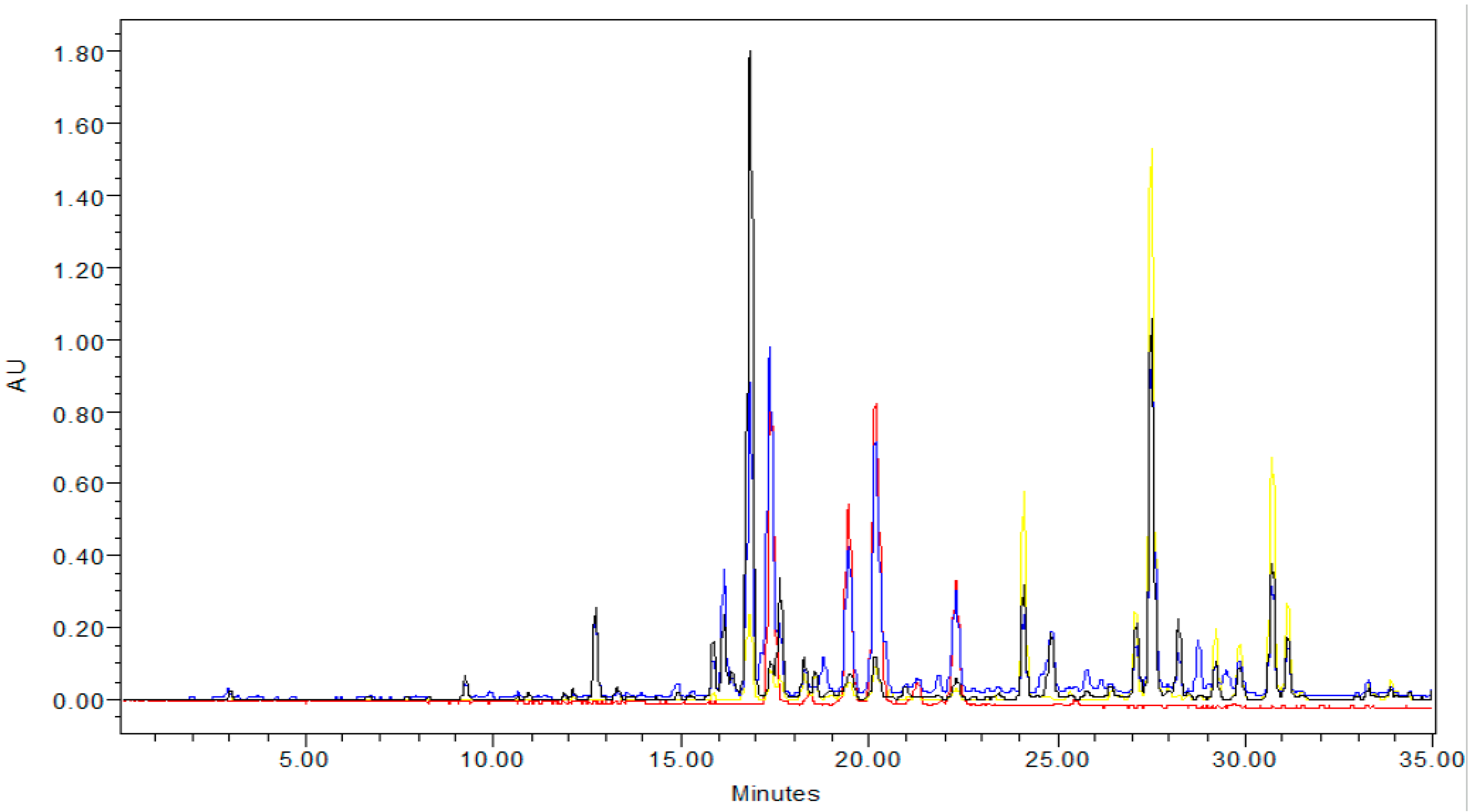
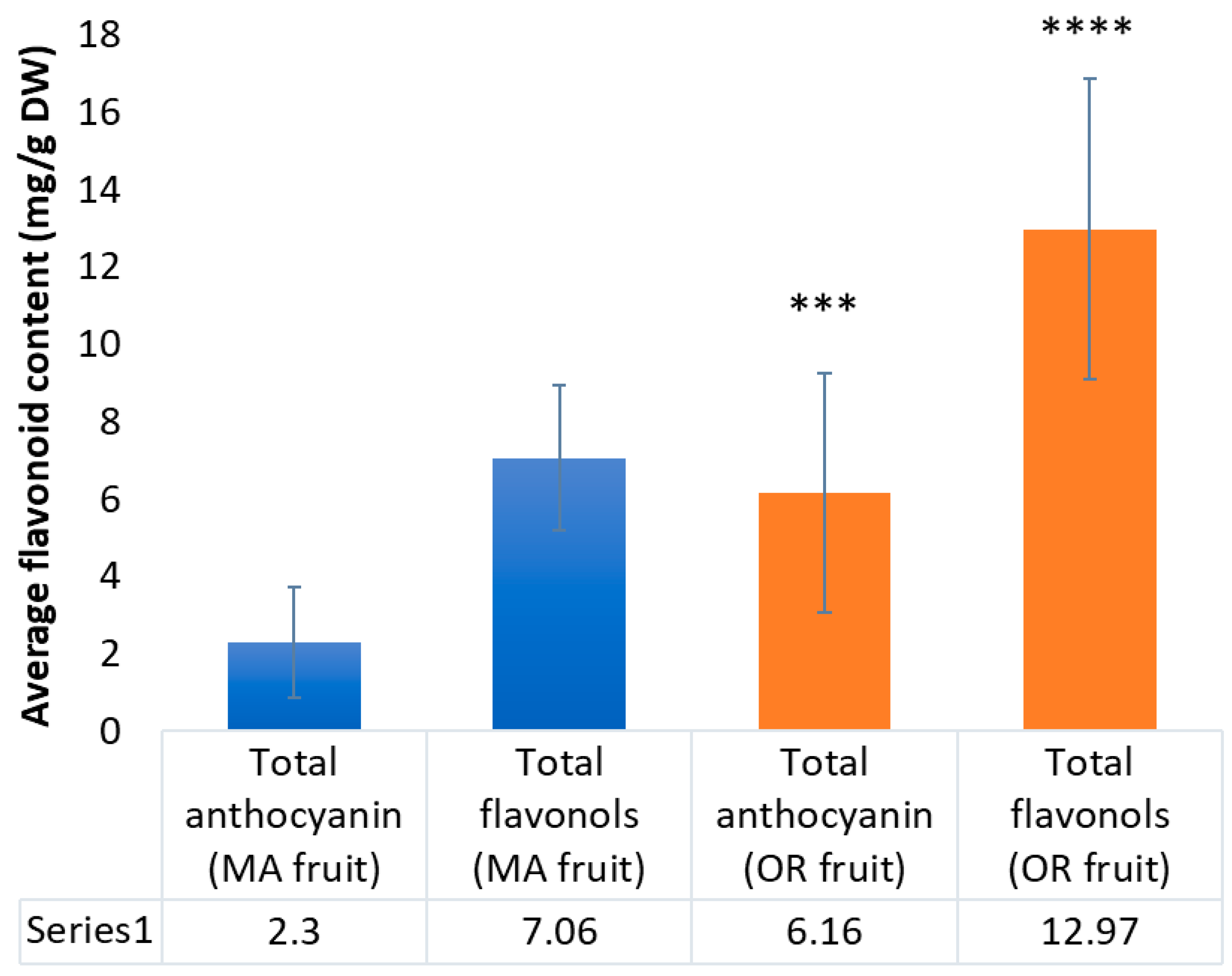
2.3. Content of Anthocyanins and Flavonols
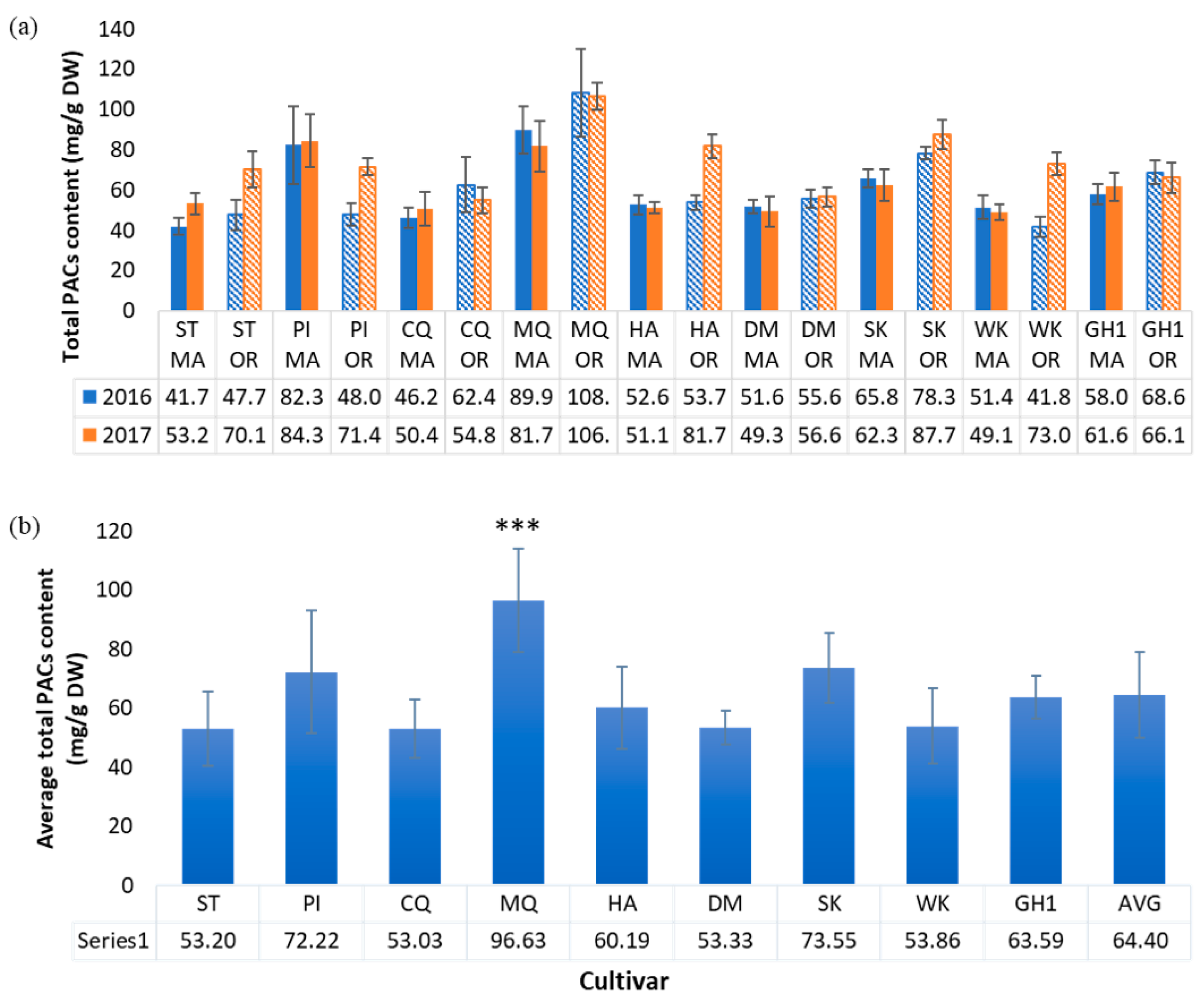
2.4. Proanthocyanidin Content
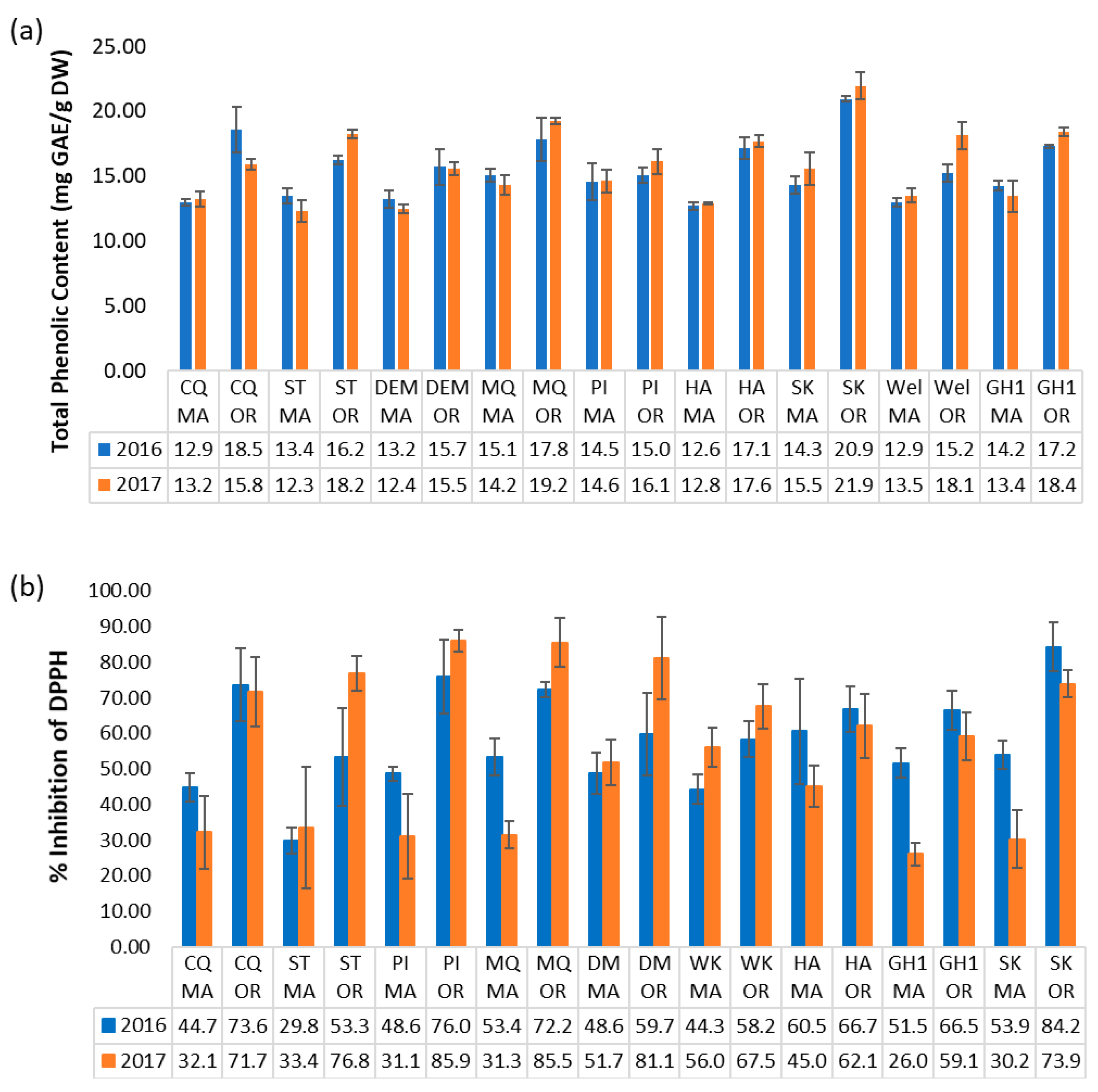
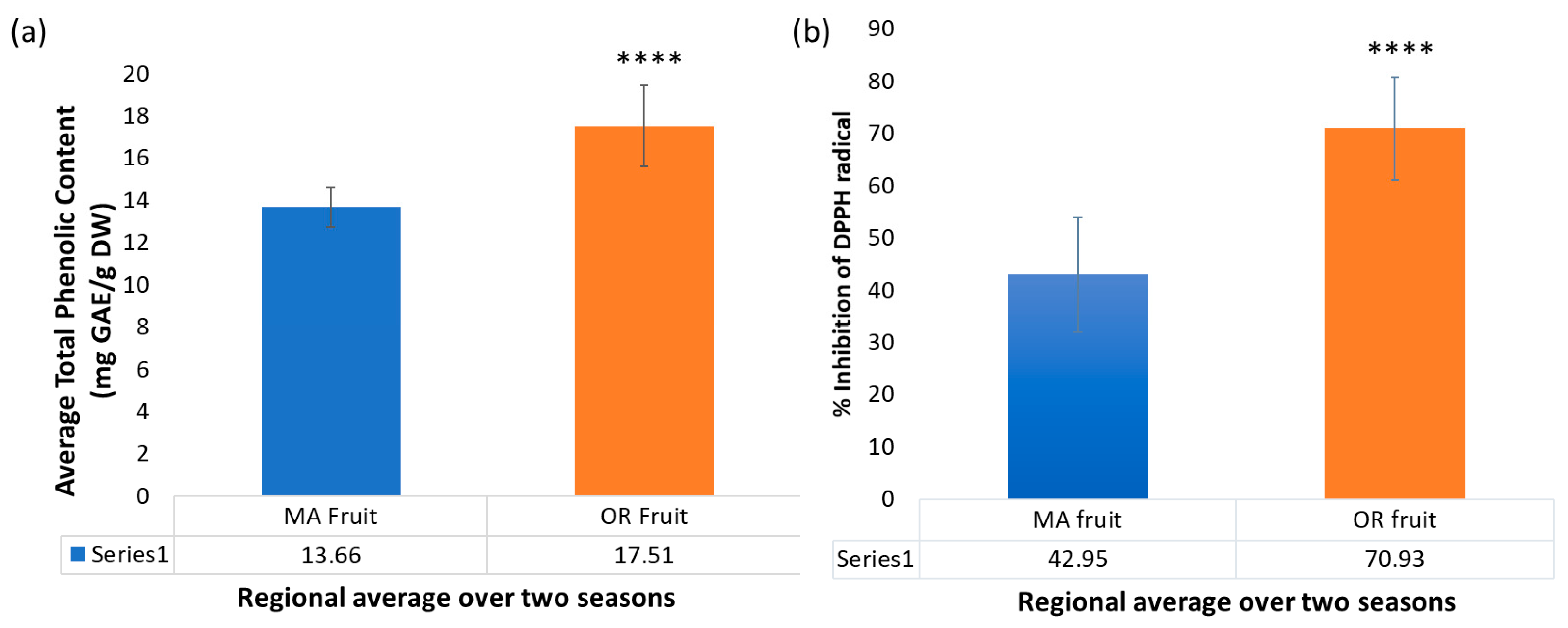
2.5. Trends in Total Phenolic Content
2.6. Correlation between Antioxidant Activity and Metabolite Content
3. Materials and Methods
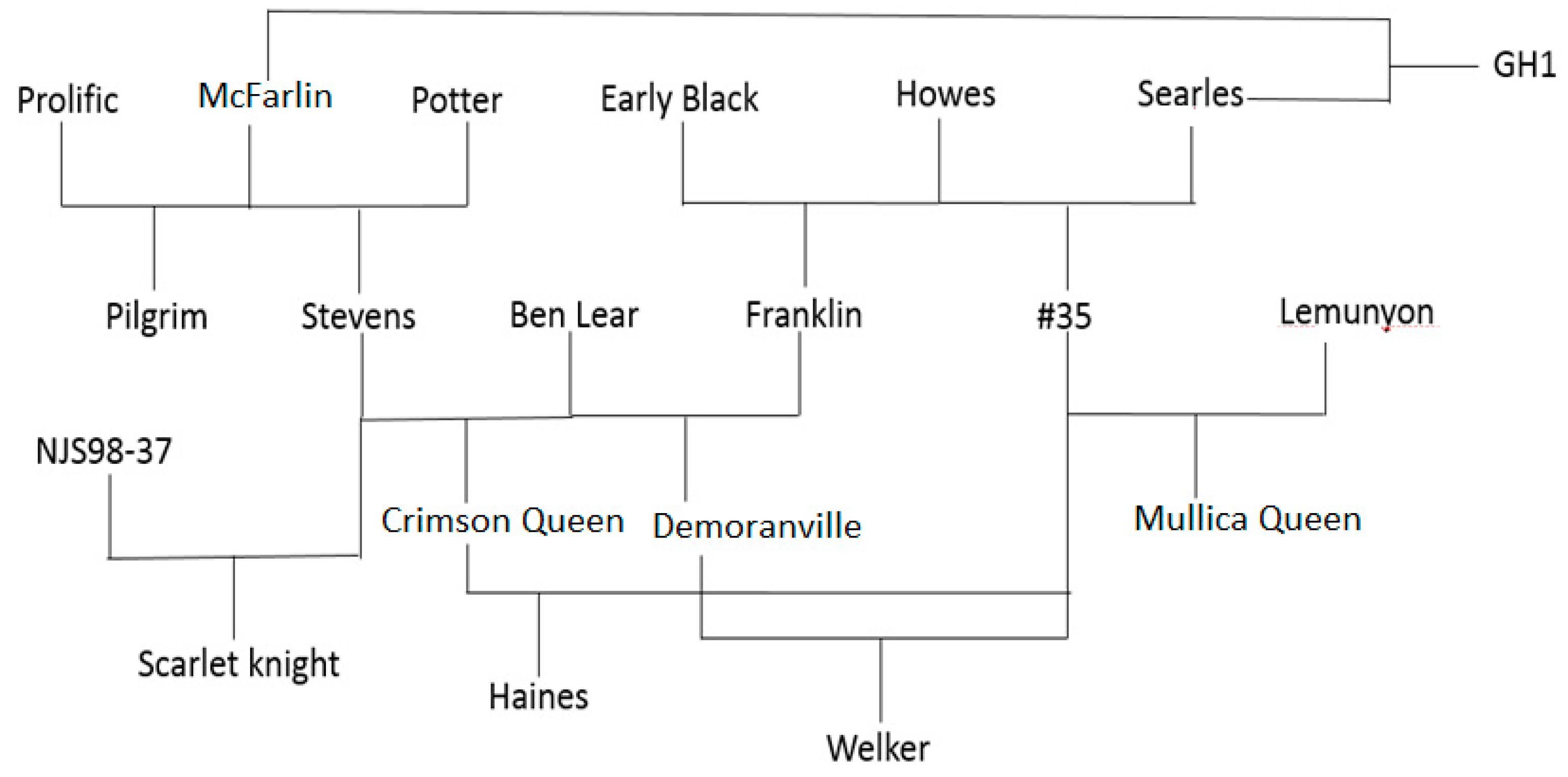
3.1. Plant Materials
3.2. Chemicals
3.3. Sample Extraction and Preparation
3.4. Metabolomic Fingerprinting and Determination of Triterpenoids and Organic Acids Using Proton (1H) qNMR
3.5. Determination of Flavonols and Anthocyanins by HPLC-DAD
3.6. Determination of Total Proanthocyanidin Content by DMAC Method
3.7. Determination of Total Phenolic Content by Folin–Ciocalteu Assay
3.8. Determination of Antioxidant Activity by DPPH Free-Radical Scavenging Assay
3.9. Chemometric and Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fresh Cranberry Production and Top Producing Countries. Available online: https://www.tridge.com/intelligences/cranberry/production (accessed on 15 September 2023).
- Cranberry Production in Top-Producing States to Increase Modestly in 2021. Available online: http://www.ers.usda.gov/data-products/chart-gallery/gallery/chart-detail/?chartId=102649 (accessed on 28 June 2022).
- Blumberg, J.B.; Camesano, T.A.; Cassidy, A.; Kris-Etherton, P.; Howell, A.; Manach, C.; Ostertag, L.M.; Sies, H.; Skulas-Ray, A.; Vita, J.A. Cranberries and Their Bioactive Constituents in Human Health. Adv. Nutr. 2013, 4, 618–632. [Google Scholar] [CrossRef] [PubMed]
- McKay, D.L.; Blumberg, J.B. Cranberries (Vaccinium macrocarpon) and Cardiovascular Disease Risk Factors. Nutr. Rev. 2007, 65, 490–502. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Han, Y.; Gu, M.; Song, M.; Wu, X.; Li, Z.; Li, F.; Goulette, T.; Xiao, H. Dietary Cranberry Suppressed Colonic Inflammation and Alleviated Gut Microbiota Dysbiosis in Dextran Sodium Sulfate-Treated Mice. Food Funct. 2019, 10, 6331–6341. [Google Scholar] [CrossRef]
- Seeram, N.P.; Adams, L.S.; Hardy, M.L.; Heber, D. Total Cranberry Extract versus Its Phytochemical Constituents: Antiproliferative and Synergistic Effects against Human Tumor Cell Lines. J. Agric. Food Chem. 2004, 52, 2512–2517. [Google Scholar] [CrossRef] [PubMed]
- Kresty, L.A.; Clarke, J.; Ezell, K.; Exum, A.; Howell, A.B.; Guettouche, T. MicroRNA Alterations in Barrett’s Esophagus, Esophageal Adenocarcinoma, and Esophageal Adenocarcinoma Cell Lines Following Cranberry Extract Treatment: Insights for Chemoprevention. J. Carcinog. 2011, 10, 34. [Google Scholar] [CrossRef]
- Weh, K.M.; Clarke, J.; Kresty, L.A. Cranberries and Cancer: An Update of Preclinical Studies Evaluating the Cancer Inhibitory Potential of Cranberry and Cranberry Derived Constituents. Antioxidants 2016, 5, 27. [Google Scholar] [CrossRef]
- Narayansingh, R.; Hurta, R.A. Cranberry Extract and Quercetin Modulate the Expression of Cyclooxygenase-2 (COX-2) and IκBα in Human Colon Cancer Cells. J. Sci. Food Agric. 2009, 89, 542–547. [Google Scholar] [CrossRef]
- Côté, J.; Caillet, S.; Doyon, G.; Sylvain, J.-F.; Lacroix, M. Bioactive Compounds in Cranberries and Their Biological Properties. Crit. Rev. Food Sci. Nutr. 2010, 50, 666–679. [Google Scholar] [CrossRef]
- Ma, H.; Johnson, S.L.; Liu, W.; DaSilva, N.A.; Meschwitz, S.; Dain, J.A.; Seeram, N.P. Evaluation of Polyphenol Anthocyanin-Enriched Extracts of Blackberry, Black Raspberry, Blueberry, Cranberry, Red Raspberry, and Strawberry for Free Radical Scavenging, Reactive Carbonyl Species Trapping, Anti-Glycation, Anti-β-Amyloid Aggregation, and Microglial Neuroprotective Effects. Int. J. Mol. Sci. 2018, 19, 461. [Google Scholar] [CrossRef]
- Wu, X.; Xue, L.; Tata, A.; Song, M.; Neto, C.C.; Xiao, H. Bioactive Components of Polyphenol-Rich and Non-Polyphenol-Rich Cranberry Fruit Extracts and Their Chemopreventive Effects on Colitis-Associated Colon Cancer. J. Agric. Food Chem. 2020, 68, 6845–6853. [Google Scholar] [CrossRef]
- Wang, C.; Gao, Y.; Zhang, Z.; Chen, C.; Chi, Q.; Xu, K.; Yang, L. Ursolic Acid Protects Chondrocytes, Exhibits Anti-Inflammatory Properties via Regulation of the NF-ΚB/NLRP3 Inflammasome Pathway and Ameliorates Osteoarthritis. Biomed. Pharmacother. 2020, 130, 110568. [Google Scholar] [CrossRef] [PubMed]
- Mlala, S.; Oyedeji, A.O.; Gondwe, M.; Oyedeji, O.O. Ursolic Acid and Its Derivatives as Bioactive Agents. Molecules 2019, 24, 2751. [Google Scholar] [CrossRef]
- Wang, X.-T.; Gong, Y.; Zhou, B.; Yang, J.-J.; Cheng, Y.; Zhao, J.-G.; Qi, M.-Y. Ursolic Acid Ameliorates Oxidative Stress, Inflammation and Fibrosis in Diabetic Cardiomyopathy Rats. Biomed. Pharmacother. 2018, 97, 1461–1467. [Google Scholar] [CrossRef] [PubMed]
- Kondo, M.; MacKinnon, S.L.; Craft, C.C.; Matchett, M.D.; Hurta, R.A.R.; Neto, C.C. Ursolic Acid and Its Esters: Occurrence in Cranberries and Other Vaccinium Fruit and Effects on Matrix Metalloproteinase Activity in DU145 Prostate Tumor Cells: Anti-Tumor Activity and Content of Ursolic Acid from Vaccinium Fruit. J. Sci. Food Agric. 2011, 91, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Cranberry Marketing Committee. Cranberry Varieties. Available online: https://reports.uscranberries.com/TheCranberryStory/Varieties/ (accessed on 19 January 2023).
- USDA/NASS QuickStats Ad-Hoc Query Tool. Available online: https://quickstats.nass.usda.gov/results/4CD92CD5-FC42-347C-ABFD-3487DC6D149D (accessed on 28 June 2022).
- Brown, P.N.; Murch, S.J.; Shipley, P. Phytochemical Diversity of Cranberry (Vaccinium macrocarpon Aiton) Cultivars by Anthocyanin Determination and Metabolomic Profiling with Chemometric Analysis. J. Agric. Food Chem. 2012, 60, 261–271. [Google Scholar] [CrossRef]
- Wang, Y.; Johnson-Cicalese, J.; Singh, A.P.; Vorsa, N. Characterization and Quantification of Flavonoids and Organic Acids over Fruit Development in American Cranberry (Vaccinium macrocarpon) Cultivars Using HPLC and APCI-MS/MS. Plant Sci. Int. J. Exp. Plant Biol. 2017, 262, 91–102. [Google Scholar] [CrossRef]
- Lu, Y.; Pekerti, B.N.; Toh, Z.S.; Broom, F.; Savage, G.; Liu, S.Q.; Huang, D. Physico-Chemical Parameters and Proanthocyanidin Profiles of Cranberries Cultivated in New Zealand. J. Food Compos. Anal. 2017, 63, 1–7. [Google Scholar] [CrossRef]
- Georgescu, C.; Frum, A.; Virchea, L.-I.; Sumacheva, A.; Shamtsyan, M.; Gligor, F.-G.; Olah, N.K.; Mathe, E.; Mironescu, M. Geographic Variability of Berry Phytochemicals with Antioxidant and Antimicrobial Properties. Molecules 2022, 27, 4986. [Google Scholar] [CrossRef]
- Murphy, B.T.; MacKinnon, S.L.; Yan, X.; Hammond, G.B.; Vaisberg, A.J.; Neto, C.C. Identification of Triterpene Hydroxycinnamates with in Vitro Antitumor Activity from Whole Cranberry Fruit (Vaccinium macrocarpon). J. Agric. Food Chem. 2003, 51, 3541–3545. [Google Scholar] [CrossRef]
- Yuk, J.; McIntyre, K.L.; Fischer, C.; Hicks, J.; Colson, K.L.; Lui, E.; Brown, D.; Arnason, J.T. Distinguishing Ontario Ginseng Landraces and Ginseng Species Using NMR-Based Metabolomics. Anal. Bioanal. Chem. 2013, 405, 4499–4509. [Google Scholar] [CrossRef]
- Markus, M.A.; Ferrier, J.; Luchsinger, S.M.; Yuk, J.; Cuerrier, A.; Balick, M.J.; Hicks, J.M.; Killday, K.B.; Kirby, C.W.; Berrue, F.; et al. Distinguishing Vaccinium species by Chemical Fingerprinting Based on NMR Spectra, Validated with Spectra Collected in Different Laboratories. Planta Med. 2014, 80, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Turbitt, J.R.; Colson, K.L.; Killday, K.B.; Milstead, A.; Neto, C.C. Application of 1 H-NMR-Based Metabolomics to the Analysis of Cranberry (Vaccinium macrocarpon) Supplements. Phytochem. Anal. PCA 2020, 31, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Vorsa, N.; Harrington, P.d.B.; Chen, P. Nontargeted Metabolomic Study on Variation of Phenolics in Different Cranberry Cultivars Using UPLC-IM—HRMS. J. Agric. Food Chem. 2018, 66, 12206–12216. [Google Scholar] [CrossRef] [PubMed]
- Vvedenskaya, I.O.; Rosen, R.T.; Guido, J.E.; Russell, D.J.; Mills, K.A.; Vorsa, N. Characterization of Flavonols in Cranberry (Vaccinium macrocarpon) Powder. J. Agric. Food Chem. 2004, 52, 188–195. [Google Scholar] [CrossRef]
- Chen, H.; Zuo, Y.; Deng, Y. Separation and Determination of Flavonoids and Other Phenolic Compounds in Cranberry Juice by High-Performance Liquid Chromatography. J. Chromatogr. A 2001, 913, 387–395. [Google Scholar] [CrossRef]
- Simmler, C.; Napolitano, J.G.; McAlpine, J.B.; Chen, S.-N.; Pauli, G.F. Universal Quantitative NMR Analysis of Complex Natural Samples. Curr. Opin. Biotechnol. 2014, 25, 51–59. [Google Scholar] [CrossRef]
- Hicks, J.M.; Muhammad, A.; Ferrier, J.; Saleem, A.; Cuerrier, A.; Arnason, J.T.; Colson, K.L. Quantification of Chlorogenic Acid and Hyperoside Directly from Crude Blueberry (Vaccinium angustifolium) Leaf Extract by NMR Spectroscopy Analysis: Single-Laboratory Validation. J. AOAC Int. 2012, 95, 1406–1411. [Google Scholar] [CrossRef]
- Huang, Y.; Li, X.; Peng, X.; Adegoke, A.T.; Chen, J.; Su, H.; Hu, G.; Wei, G.; Qiu, M. NMR-Based Structural Classification, Identification, and Quantification of Triterpenoids from Edible Mushroom Ganoderma Resinaceum. J. Agric. Food Chem. 2020, 68, 2816–2825. [Google Scholar] [CrossRef]
- Fajardo, D.; Morales, J.; Zhu, H.; Steffan, S.; Harbut, R.; Bassil, N.; Hummer, K.; Polashock, J.; Vorsa, N.; Zalapa, J. Discrimination of American Cranberry Cultivars and Assessment of Clonal Heterogeneity Using Microsatellite Markers. Plant Mol. Biol. Report. 2012, 31, 264–271. [Google Scholar] [CrossRef]
- Sedbare, R.; Raudone, L.; Zvikas, V.; Viskelis, J.; Liaudanskas, M.; Janulis, V. Development and Validation of the UPLC-DAD Methodology for the Detection of Triterpenoids and Phytosterols in Fruit Samples of Vaccinium macrocarpon Aiton and Vaccinium oxycoccos L. Molecules 2022, 27, 4403. [Google Scholar] [CrossRef]
- Šedbarė, R.; Siliņa, D.; Janulis, V. Evaluation of the Phytochemical Composition of Phenolic and Triterpene Compounds in Fruit of Large Cranberries (Vaccinium macrocarpon Aiton) Grown in Latvia. Plants 2022, 11, 2725. [Google Scholar] [CrossRef]
- Oszmiański, J.; Kolniak-Ostek, J.; Lachowicz, S.; Gorzelany, J.; Matłok, N. Phytochemical Compounds and Antioxidant Activity in Different Cultivars of Cranberry (Vaccinium macrocarpon L.). J. Food Sci. 2017, 82, 2569–2575. [Google Scholar] [CrossRef] [PubMed]
- Viškelis, J.; Uselis, N.; Liaudanskas, M.; Lanauskas, J.; Bielicki, P.; Univer, T.; Lepsis, J.; Kviklys, D. Location Effects across Northeastern Europe on Bioactive Compounds in Apple Fruit. Agric. Food Sci. 2019, 28, 93–100. [Google Scholar] [CrossRef]
- Brown, P.N.; Shipley, P.R. Determination of Anthocyanins in Cranberry Fruit and Cranberry Fruit Products by High-Performance Liquid Chromatography with Ultraviolet Detection: Single-Laboratory Validation. J. AOAC Int. 2011, 94, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Vvedenskaya, I.; Vorsa, N. Flavonoid Composition over Fruit Development and Maturation in American Cranberry, Vaccinium macrocarpon Ait. Plant Sci. 2004, 167, 1043–1054. [Google Scholar] [CrossRef]
- Cranberry Weather Summaries Archive. Available online: https://ag.umass.edu/cranberry/weather-summaries-archive (accessed on 5 July 2022).
- Bureau of Reclamation. AgriMet Columbia-Pacific Northwest Region. Available online: https://www.usbr.gov/pn/agrimet/webarcread.html (accessed on 5 July 2022).
- Carpenter, J.L.; Caruso, F.L.; Tata, A.; Vorsa, N.; Neto, C.C. Variation in Proanthocyanidin Content and Composition among Commonly Grown North American Cranberry Cultivars (Vaccinium macrocarpon): Proanthocyanidin Content of Cranberries. J. Sci. Food Agric. 2014, 94, 2738–2745. [Google Scholar] [CrossRef]
- Borowska, E.; Mazur, B.; Gadzal, R.; Kopciuch, V.; Buszewski, B. Polyphenol, Anthocyanin and Resveratrol Mass Fractions and Antioxidant Properties of Cranberry Cultivars. Food Technol. Biotechnol. 2009, 47, 56–61. [Google Scholar]
- Narwojsz, A.; Tańska, M.; Mazur, B.; Borowska, E.J. Fruit Physical Features, Phenolic Compounds Profile and Inhibition Activities of Cranberry Cultivars (Vaccinium macrocarpon) Compared to Wild-Grown Cranberry (Vaccinium oxycoccus). Plant Foods Hum. Nutr. 2019, 74, 300–306. [Google Scholar] [CrossRef]
- Wang, S.Y.; Stretch, A.W. Antioxidant Capacity in Cranberry Is Influenced by Cultivar and Storage Temperature. J. Agric. Food Chem. 2001, 49, 969–974. [Google Scholar] [CrossRef]
- Xue, L.; Liu, C.; Ma, H.; Seeram, N.P.; Neto, C.C. Anti-Inflammatory Activities of Cranberry Fruit Extracts in Human THP-1 Monocytes Are Influenced by Their Phytochemical Composition. ACS Food Sci. Technol. 2022, 2, 75–83. [Google Scholar] [CrossRef]
- Gudoityte, E.; Arandarcikaite, O.; Mazeikiene, I.; Bendokas, V.; Liobikas, J. Ursolic and Oleanolic Acids: Plant Metabolites with Neuroprotective Potential. Int. J. Mol. Sci. 2021, 22, 4599. [Google Scholar] [CrossRef]
- Dreier, L.; Wider, G. Concentration Measurements by PULCON Using X-Filtered or 2D NMR Spectra. Magn. Reson. Chem. MRC 2006, 44 (Suppl. S1), S206–S212. [Google Scholar] [CrossRef]
- Feliciano, R.P.; Shea, M.P.; Shanmuganayagam, D.; Krueger, C.G.; Howell, A.B.; Reed, J.D. Comparison of Isolated Cranberry (Vaccinium macrocarpon Ait.) Proanthocyanidins to Catechin and Procyanidins A2 and B2 for Use as Standards in the 4-(Dimethylamino)Cinnamaldehyde Assay. J. Agric. Food Chem. 2012, 60, 4578–4585. [Google Scholar] [CrossRef]
- Prior, R.L.; Fan, E.; Ji, H.; Howell, A.; Nio, C.; Payne, M.J.; Reed, J. Multi-Laboratory Validation of a Standard Method for Quantifying Proanthocyanidins in Cranberry Powders. J. Sci. Food Agric. 2010, 90, 1473–1478. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of Total Phenolic Content and Other Oxidation Substrates in Plant Tissues Using Folin-Ciocalteu Reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Liu, J.; Zhao, J.; Dai, Z.; Lin, R.; Wang, G.; Ma, S. A Pair of New Antioxidant Phenolic Acid Stereoisomers Isolated from Danshen Injection (Lyophilized Powder). Molecules 2014, 19, 1786–1794. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar/Region | Growing Season | Ursolic Acid | Oleanolic Acid | Citric Acid | Malic Acid |
|---|---|---|---|---|---|
| ST/MA | 2016 | 9.63 ± 2.39 | 4.23 ± 0.55 | 57.61 ± 9.00 | 43.78 ± 10.25 |
| 2017 | 10.44 ± 3.28 | 3.83 ± 1.54 | 81.67 ± 10.91 | 54.98 ± 7.17 | |
| ST/OR | 2016 | 7.09 ± 1.05 | 2.11 ± 0.68 | 38.69 ± 3.69 | 40.36 ± 8.91 |
| 2017 | 7.01 ± 1.53 | 2.23 ± 1.18 | 47.86 ± 9.20 | 48.54 ± 7.73 | |
| MQ/MA | 2016 | 9.02 ± 2.50 | 4.24 ± 1.18 | 50.89 ± 4.96 | 78.37 ± 12.07 |
| 2017 | 9.61 ± 2.85 | 3.01 ± 0.90 | 67.31 ± 10.91 | 78.63 ± 9.16 | |
| MQ/OR | 2016 | 6.25 ± 1.55 | 1.54 ± 0.28 | 47.81 ± 15.67 | 60.56 ± 7.72 |
| 2017 | 12.32 ± 5.28 | 2.76 ± 1.58 | 34.97 ± 8.95 | 54.59 ± 8.29 | |
| DM/MA | 2016 | 9.90 ± 2.57 | 3.59 ± 2.59 | 44.11 ± 9.49 | 75.68 ± 10.75 |
| 2017 | 8.62 ± 2.66 | 4.27 ± 1.09 | 59.27 ± 12.83 | 79.92 ± 8.63 | |
| DM/OR | 2016 | 3.18 ± 1.25 | 1.27 ± 0.06 | 45.86 ± 12.79 | 52.21 ± 6.24 |
| 2017 | 6.68 ± 2.39 | 2.05 ± 0.36 | 53.85 ± 13.96 | 59.36 ± 5.81 | |
| PI/MA | 2016 | 12.83 ± 1.78 | 5.80 ± 0.68 | 52.40 ± 9.48 | 51.43 ± 14.32 |
| 2017 | 8.94 ± 2.35 | 2.49 ± 0.93 | 76.49 ± 18.91 | 66.00 ± 12.48 | |
| PI/OR | 2016 | 4.31 ± 1.39 | 1.32 ± 0.99 | 52.41 ± 6.64 | 52.08 ± 8.70 |
| 2017 | 3.09 ± 1.72 | 1.09 ± 0.34 | 63.30 ± 13.73 | 51.46 ± 7.87 | |
| CQ/MA | 2016 | 9.18 ± 2.34 | 3.84 ± 1.22 | 64.30 ± 4.60 | 86.43 ± 10.69 |
| 2017 | 9.50 ± 3.92 | 3.72 ± 1.70 | 84.59 ± 21.09 | 81.98 ± 10.72 | |
| CQ/OR | 2016 | 2.72 ± 1.18 | 2.25 ± 0.65 | 25.95 ± 6.22 | 39.72 ± 4.46 |
| 2017 | 5.60 ± 0.67 | 1.75 ± 0.60 | 58.71 ± 23.22 | 73.67 ± 12.36 | |
| HA/MA | 2016 | 15.39 ± 4.31 | 7.60 ± 2.48 | 57.59 ± 8.39 | 88.27 ± 29.78 |
| 2017 | 7.34 ± 1.20 | 2.60 ± 0.88 | 40.42 ± 15.04 | 104.78 ± 15.76 | |
| HA/OR | 2016 | 5.92 ± 1.84 | 2.30 ± 0.81 | 64.37 ± 17.62 | 56.59 ± 12.12 |
| 2017 | 6.10 ± 2.61 | 1.66 ± 0.89 | 30.97 ± 13.32 | 63.23 ± 8.55 | |
| SK/MA | 2016 | 10.00 ± 2.27 | 3.39 ± 1.38 | 75.26 ± 16.41 | 58.19 ± 12.07 |
| 2017 | 9.46 ± 3.41 | 3.25 ± 1.38 | 80.65 ± 14.47 | 57.94 ± 6.52 | |
| SK/OR | 2016 | 6.01 ± 2.38 | 1.32 ± 0.60 | 61.52 ± 13.71 | 38.94 ± 6.82 |
| 2017 | 8.20 ± 3.73 | 1.66 ± 0.57 | 51.80 ± 7.94 | 40.84 ± 6.99 | |
| WK/MA | 2016 | 16.80 ± 9.95 | 5.99 ± 2.82 | 44.02 ± 10.32 | 76.37 ± 11.90 |
| 2017 | 10.76 ± 2.87 | 3.05 ± 0.98 | 53.35 ± 2.87 | 88.29 ± 0.98 | |
| WK/OR | 2016 | 4.40 ± 3.31 | 1.61 ± 1.11 | 34.46 ± 9.18 | 51.17 ± 9.33 |
| 2017 | 4.72 ± 0.89 | 1.10 ± 0.27 | 33.50 ± 5.02 | 65.45 ± 11.28 | |
| GH1/MA | 2016 | 11.46 ± 4.75 | 4.04 ± 1.38 | 68.13 ± 9.44 | 58.19 ± 1.38 |
| 2017 | 8.45 ± 1.90 | 2.46 ± 1.16 | 75.84 ± 19.68 | 57.83 ± 9.26 | |
| GH1/OR | 2016 | 5.16 ± 3.26 | 1.07 ± 0.32 | 65.68 ± 8.16 | 41.27 ± 3.94 |
| 2017 | 8.20 ± 3.97 | 1.86 ± 0.82 | 56.71 ± 8.73 | 40.97 ± 5.21 |
| Cultivar/Region | Year | Cyanidin Glycosides | Peonidin Glycosides | Total Anthocyanin | Quercetin-3-Galactoside | Total Flavonols | Total Phenolics a | Total PACs |
|---|---|---|---|---|---|---|---|---|
| ST/MA | 2016 | 0.87 ± 0.16 | 2.01 ± 0.38 | 2.88 ± 0.54 | 2.78 ± 0.67 | 6.88 ± 1.46 | 13.46 ± 0.59 | 41.71 ± 4.20 |
| 2017 | 0.20 ± 0.13 | 0.42 ± 0.25 | 0.62 ± 0.24 | 2.58 ± 0.35 | 8.17 ± 1.07 | 12.32 ± 0.84 | 53.20 ± 5.15 | |
| ST/OR | 2016 | 1.22 ± 0.28 | 4.29 ± 1.20 | 5.50 ± 1.48 | 3.77 ± 0.76 | 10.44 ± 2.94 | 16.25 ± 0.33 | 47.71 ± 7.67 |
| 2017 | 0.55 ± 0.22 | 1.65 ± 0.67 | 2.20 ± 1.02 | 4.07 ± 0.90 | 13.37 ± 1.89 | 18.22 ± 0.35 | 70.17 ± 9.11 | |
| MQ/MA | 2016 | 0.68 ± 0.11 | 1.57 ± 0.29 | 2.25 ± 0.39 | 3.75 ± 0.64 | 8.92 ± 1.65 | 15.10 ± 0.49 | 89.94 ± 11.68 |
| 2017 | 0.09 ± 0.02 | 0.20 ± 0.03 | 0.29 ± 0.05 | 1.64 ± 0.37 | 5.42 ± 1.28 | 14.29 ± 0.74 | 81.77 ± 12.68 | |
| MQ/OR | 2016 | 2.15 ± 0.59 | 7.13 ± 1.33 | 9.28 ± 1.90 | 5.00 ± 2.30 | 15.99 ± 5.71 | 17.83 ± 1.65 | 108.22 ± 22.1 |
| 2017 | 1.05 ± 0.18 | 3.31 ± 0.30 | 4.36 ± 0.36 | 4.03 ± 0.77 | 13.01 ± 2.78 | 19.23 ± 0.24 | 106.58 ± 6.85 | |
| DM/MA | 2016 | 0.79 ± 0.22 | 1.76 ± 0.50 | 2.55 ± 0.71 | 2.77 ± 0.65 | 7.86 ± 2.72 | 13.22 ± 0.69 | 51.69 ± 3.29 |
| 2017 | 0.79 ± 0.42 | 1.59 ± 1.22 | 2.68 ± 1.61 | 1.95 ± 0.25 | 7.20 ± 0.79 | 12.48 ± 0.33 | 49.32 ± 9.54 | |
| DM/OR | 2016 | 1.88 ± 0.89 | 7.33 ± 3.02 | 9.41 ± 3.90 | 4.11 ± 1.99 | 14.76 ± 7.38 | 15.72 ± 1.36 | 55.69 ± 4.37 |
| 2017 | 1.39 ± 0.37 | 4.97 ± 1.44 | 6.36 ± 1.79 | 1.69 ± 0.29 | 7.12 ± 0.44 | 15.55 ± 0.53 | 56.64 ± 4.6 | |
| PI/MA | 2016 | 0.95 ± 0.11 | 1.81 ± 0.17 | 2.75 ± 0.26 | 2.59 ± 0.51 | 7.04 ± 0.91 | 14.57 ± 1.39 | 82.39 ± 19.19 |
| 2017 | 0.10 ± 0.03 | 0.16 ± 0.05 | 0.25 ± 0.06 | 2.09 ± 0.31 | 7.04 ± 0.66 | 14.64 ± 0.87 | 84.38 ± 13.16 | |
| PI/OR | 2016 | 1.76 ± 0.52 | 4.20 ± 1.03 | 5.96 ± 1.56 | 4.54 ± 1.83 | 14.13 ± 5.44 | 15.07 ± 0.57 | 48.07 ± 5.59 |
| 2017 | 0.94 ± 0.16 | 2.34 ± 0.30 | 3.28 ± 0.45 | 3.48 ± 0.39 | 12.15 ± 1.59 | 16.12 ± 0.96 | 71.40 ± 4.26 | |
| CQ/MA | 2016 | 0.69 ± 0.14 | 2.03 ± 0.32 | 2.72 ± 0.46 | 3.16 ± 0.72 | 7.55 ± 1.78 | 12.95 ± 0.24 | 46.21 ± 4.93 |
| 2017 | 0.52 ± 0.15 | 1.44 ± 0.59 | 1.97 ± 0.73 | 2.09 ± 0.58 | 7.63 ± 1.54 | 13.23 ± 0.62 | 50.49 ± 8.28 | |
| CQ/OR | 2016 | 1.72 ± 0.36 | 5.76 ± 0.81 | 7.48 ± 0.86 | 6.62 ± 2.94 | 18.63 ± 5.84 | 18.58 ± 1.75 | 62.46 ± 13.68 |
| 2017 | 0.85 ± 0.31 | 3.59 ± 1.82 | 4.44 ± 1.65 | 2.92 ± 1.25 | 10.11 ± 2.22 | 15.86 ± 0.42 | 54.84 ± 6.64 | |
| HA/MA | 2016 | 1.15 ± 0.25 | 2.87 ± 0.40 | 4.02 ± 0.65 | 3.56 ± 0.82 | 10.19 ± 3.25 | 12.69 ± 0.29 | 52.63 ± 4.82 |
| 2017 | 0.51 ± 0.09 | 1.33 ± 0.35 | 1.84 ± 0.40 | 1.10 ± 0.26 | 4.29 ± 0.79 | 12.89 ± 0.07 | 51.11 ± 2.97 | |
| HA/OR | 2016 | 2.31 ± 0.10 | 6.76 ± 2.27 | 9.45 ± 3.11 | 6.00 ± 1.25 | 17.03 ± 3.33 | 17.15 ± 0.81 | 53.76 ± 3.78 |
| 2017 | 0.55 ± 0.33 | 1.51 ± 0.95 | 2.05 ± 1.27 | 2.97 ± 0.65 | 10.55 ± 1.63 | 17.69 ± 0.47 | 81.77 ± 5.97 | |
| SK/MA | 2016 | 1.81 ± 0.46 | 4.28 ± 1.08 | 6.10 ± 1.51 | 4.50 ± 0.96 | 11.37 ± 3.86 | 14.33 ± 0.67 | 65.86 ± 4.59 |
| 2017 | 0.71 ± 0.39 | 1.53 ± 0.90 | 2.24 ± 1.28 | 1.77 ± 0.50 | 5.69 ± 1.78 | 15.58 ± 1.27 | 62.32 ± 7.82 | |
| SK/OR | 2016 | 3.60 ± 0.72 | 10.93 ± 2.07 | 14.53 ± 2.72 | 7.62 ± 2.22 | 23.30 ± 8.25 | 20.94 ± 0.19 | 78.30 ± 3.03 |
| 2017 | 2.03 ± 0.36 | 5.42 ± 0.98 | 7.45 ± 1.33 | 4.09 ± 0.74 | 11.72 ± 1.98 | 21.95 ± 1.01 | 87.71 ± 7.27 | |
| WK/MA | 2016 | 0.88 ± 0.28 | 1.68 ± 0.42 | 2.56 ± 0.66 | 1.90 ± 0.52 | 5.65 ± 1.56 | 12.98 ± 0.31 | 51.48 ± 5.73 |
| 2017 | 0.85 ± 0.11 | 2.71 ± 0.61 | 3.57 ± 0.67 | 1.68 ± 0.32 | 5.25 ± 1.63 | 13.51 ± 0.56 | 49.11 ± 3.83 | |
| WK/OR | 2016 | 1.19 ± 0.38 | 4.37 ± 1.53 | 4.98 ± 1.69 | 2.13 ± 0.07 | 9.36 ± 1.57 | 15.22 ± 0.66 | 41.83 ± 5.14 |
| 2017 | 0.85 ± 0.19 | 2.25 ± 0.62 | 3.11 ± 0.81 | 2.64 ± 0.91 | 10.18 ± 2.77 | 18.12 ± 1.05 | 73.02 ± 5.42 | |
| GH1/MA | 2016 | 0.47 ± 0.13 | 0.95 ± 0.41 | 1.37 ± 0.57 | 2.31 ± 0.50 | 6.45 ± 1.25 | 14.27 ± 0.41 | 58.00 ± 5.07 |
| 2017 | 0.24 ± 0.06 | 0.49 ± 0.15 | 0.73 ± 0.22 | 1.30 ± 0.52 | 4.41 ± 1.41 | 13.44 ± 1.24 | 61.66 ± 7.07 | |
| GH1/OR | 2016 | 1.17 ± 0.20 | 4.26 ± 1.90 | 5.42 ± 2.05 | 3.26 ± 0.93 | 11.25 ± 1.90 | 17.29 ± 0.11 | 68.61 ± 5.92 |
| 2017 | 1.33 ± 0.24 | 4.29 ± 0.92 | 5.62 ± 1.15 | 3.59 ± 0.82 | 10.35 ± 2.05 | 18.40 ± 0.34 | 66.10 ± 7.45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xue, L.; Otieno, M.; Colson, K.; Neto, C. Influence of the Growing Region on the Phytochemical Composition and Antioxidant Properties of North American Cranberry Fruit (Vaccinium macrocarpon Aiton). Plants 2023, 12, 3595. https://doi.org/10.3390/plants12203595
Xue L, Otieno M, Colson K, Neto C. Influence of the Growing Region on the Phytochemical Composition and Antioxidant Properties of North American Cranberry Fruit (Vaccinium macrocarpon Aiton). Plants. 2023; 12(20):3595. https://doi.org/10.3390/plants12203595
Chicago/Turabian StyleXue, Liang, Maureen Otieno, Kimberly Colson, and Catherine Neto. 2023. "Influence of the Growing Region on the Phytochemical Composition and Antioxidant Properties of North American Cranberry Fruit (Vaccinium macrocarpon Aiton)" Plants 12, no. 20: 3595. https://doi.org/10.3390/plants12203595