
The Effect of Topophysis on the In Vitro Development of Handroanthus guayacan and on Its Metabolism of Meta-Topolin Riboside

Abstract
:1. Introduction
2. Results
2.1. Morphological Growth Parameters
2.2. Hormone Analysis
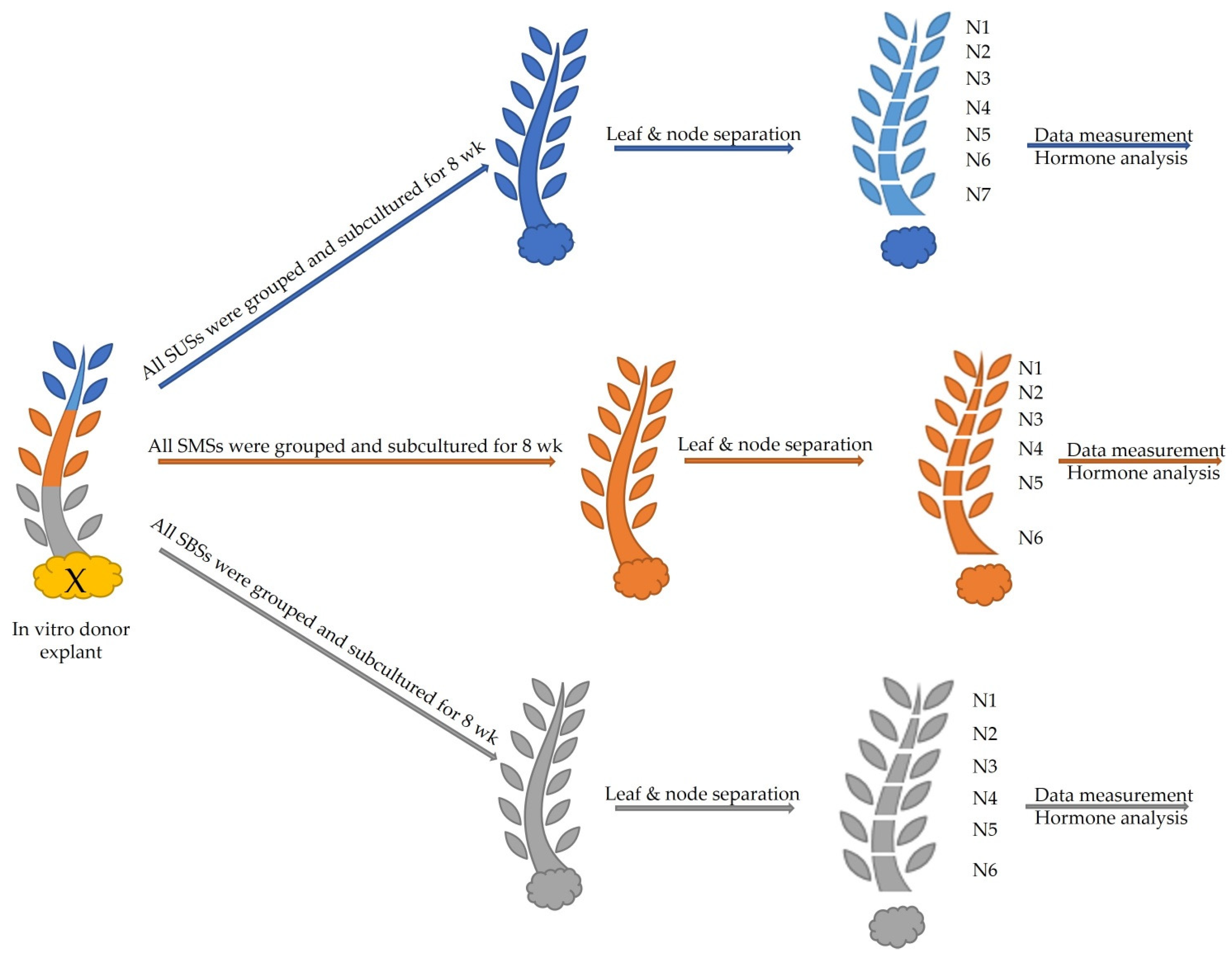
2.2.1. Shoots Derived from the Upper Section (SUSs)
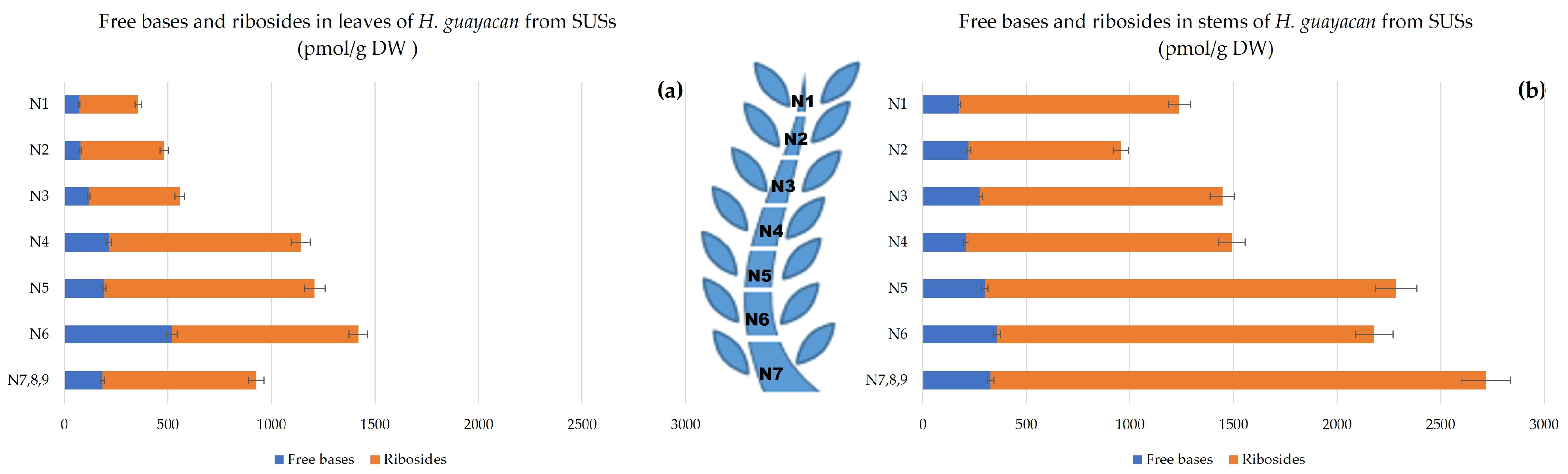
- Aromatic Cytokinin Free Bases and Riboside Isomers;
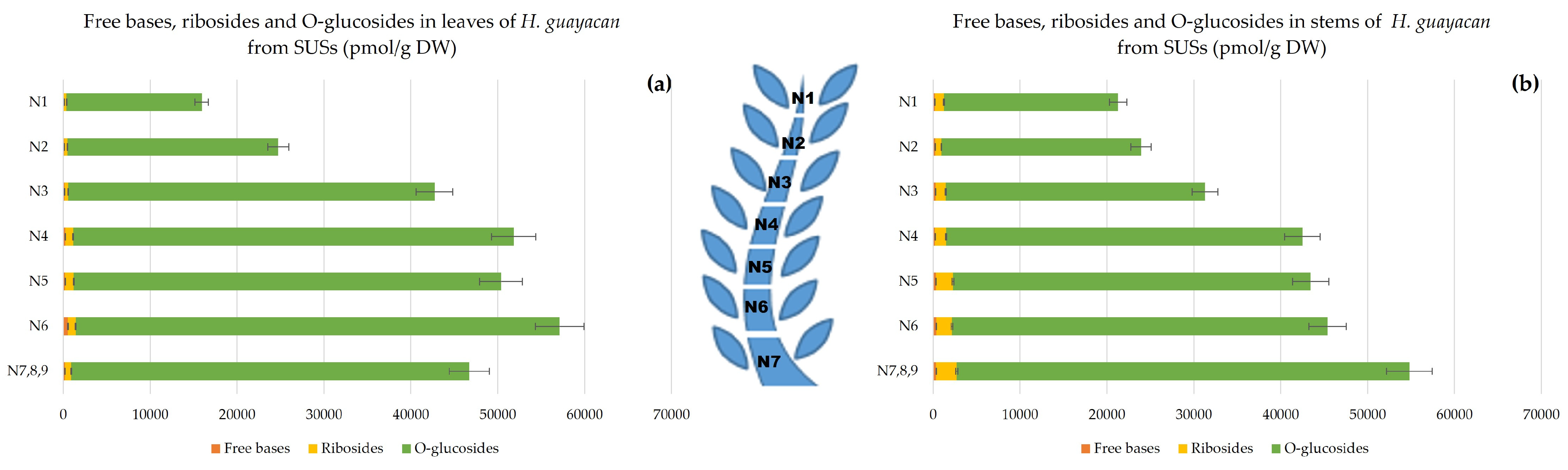
- Topolin-O-Glucosides;
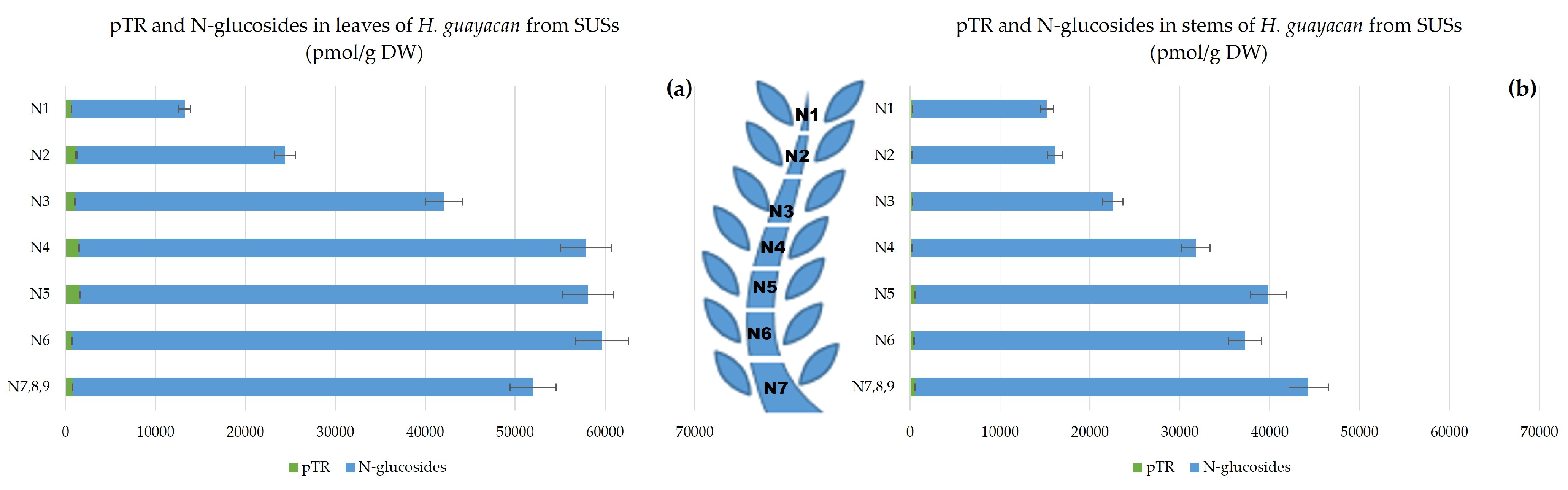
- Aromatic Cytokinin N-Glucosides and pTR;
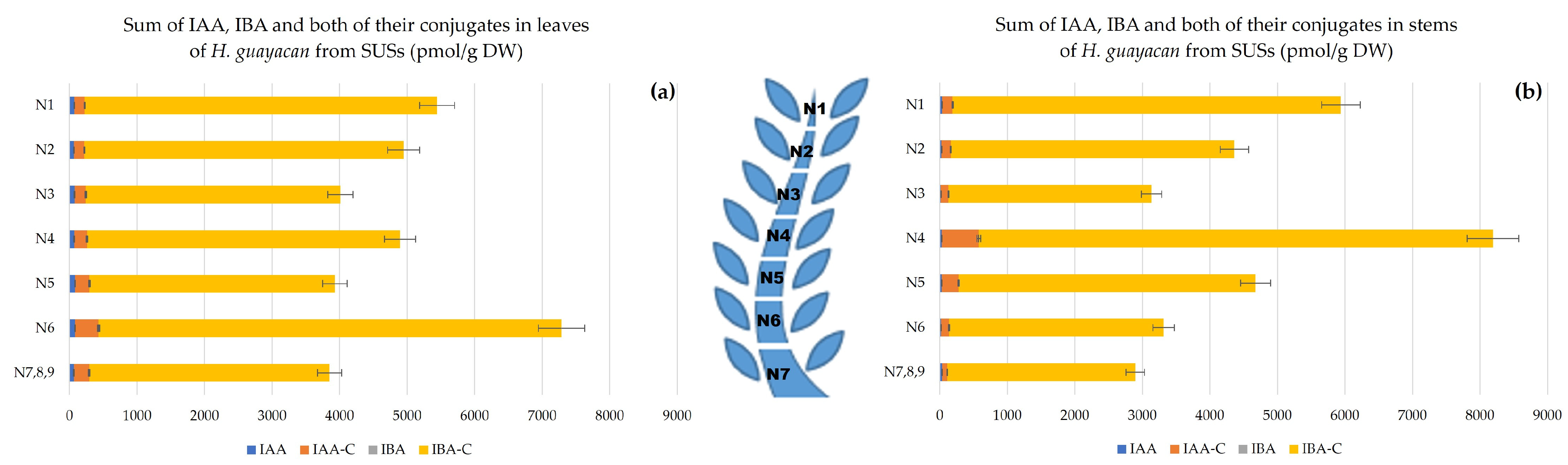
- Auxins.
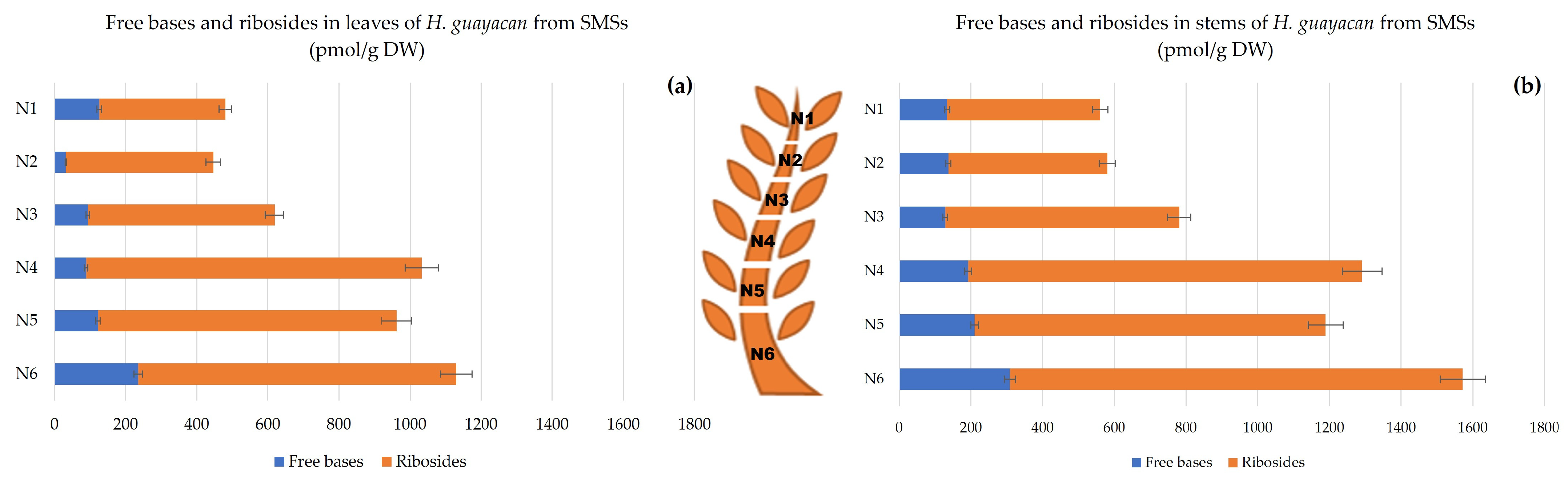
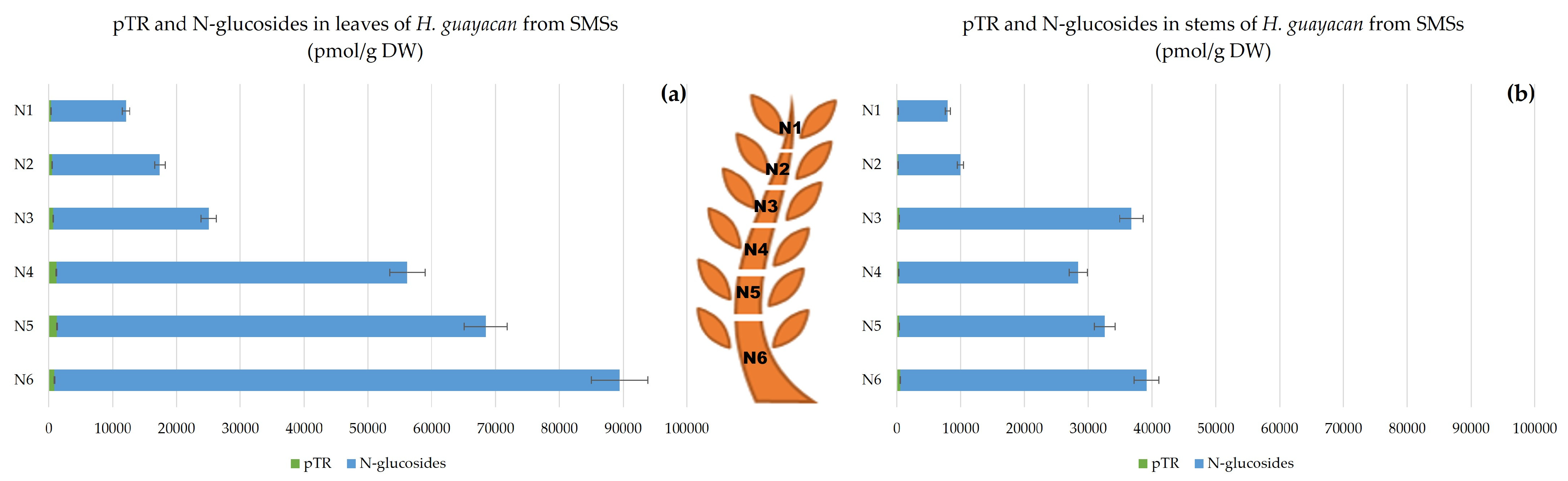
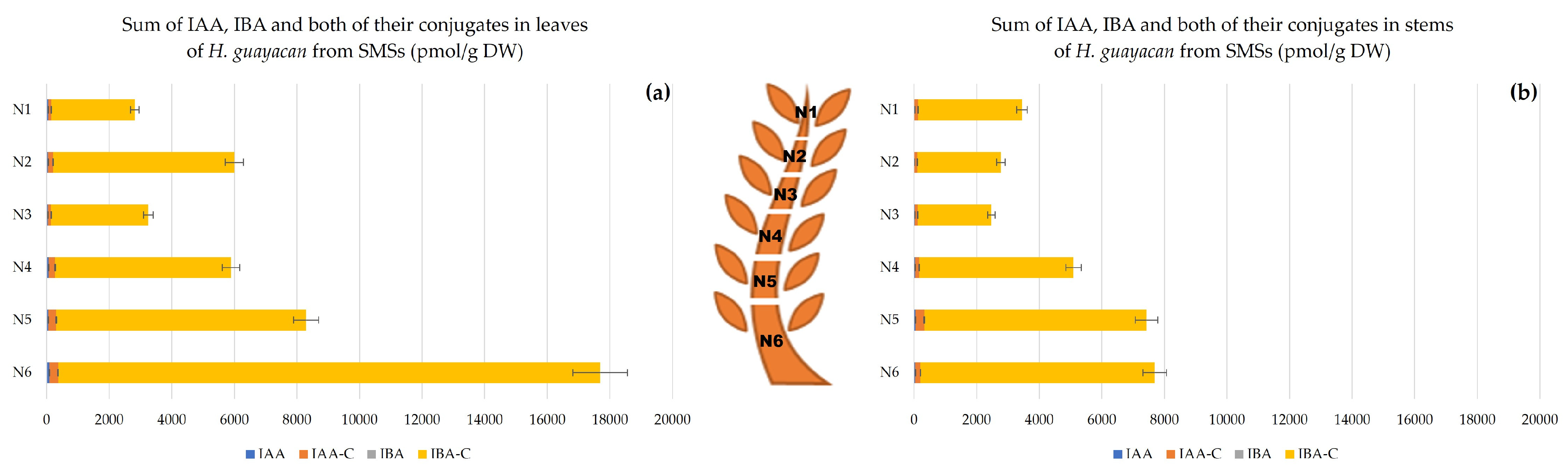
2.2.2. Shoots Derived from the Middle Section (SMSs)
- Aromatic Cytokinin Free Bases and Riboside Isomers;
- Topolin-O-Glucosides;
- Aromatic N-Glucosides and pTR;
- Auxins.
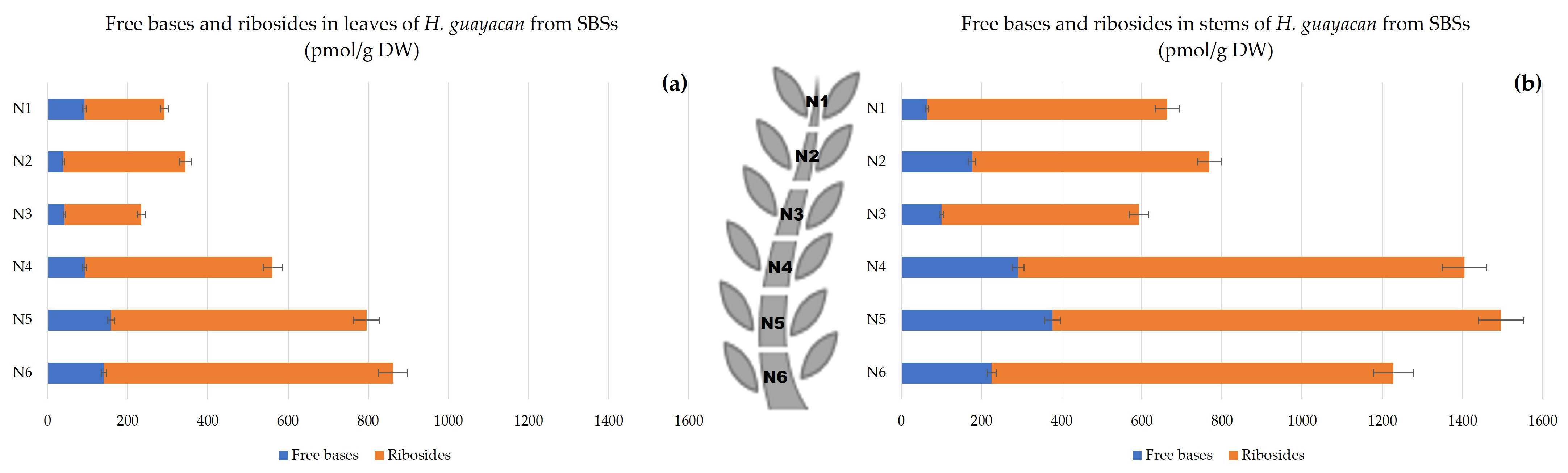
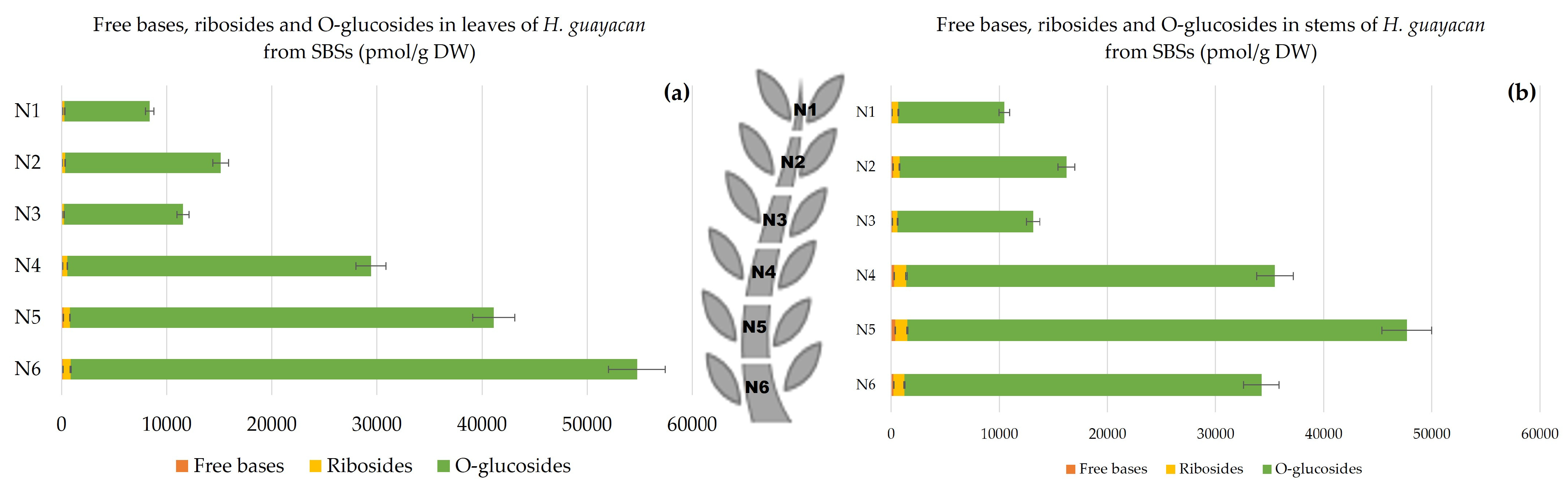
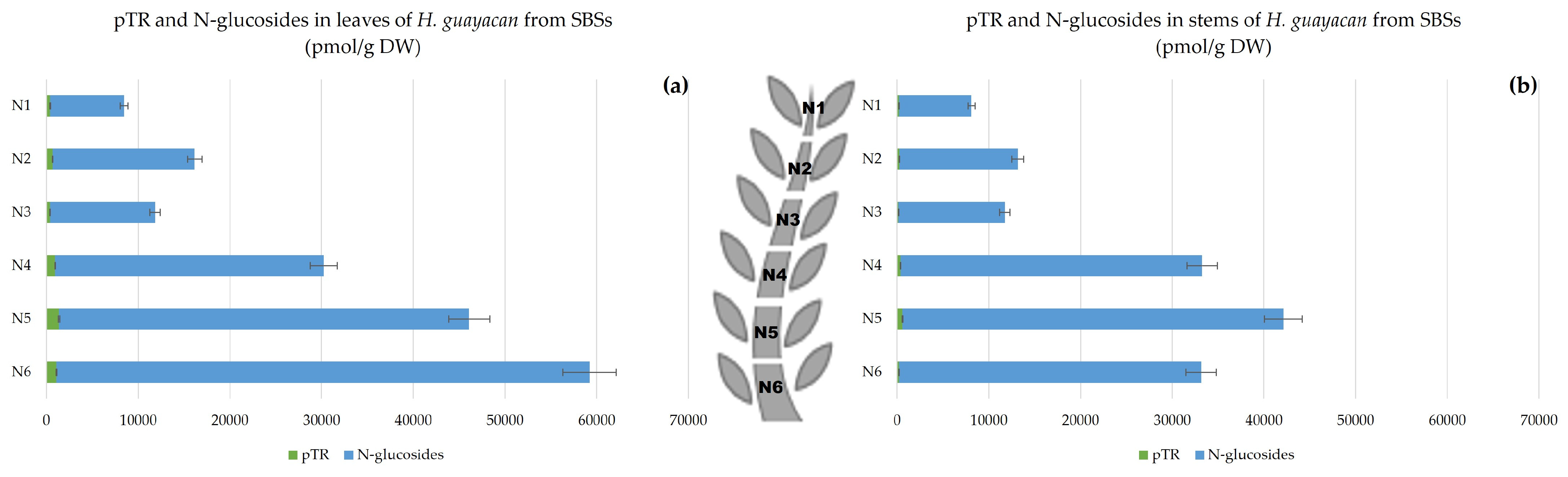
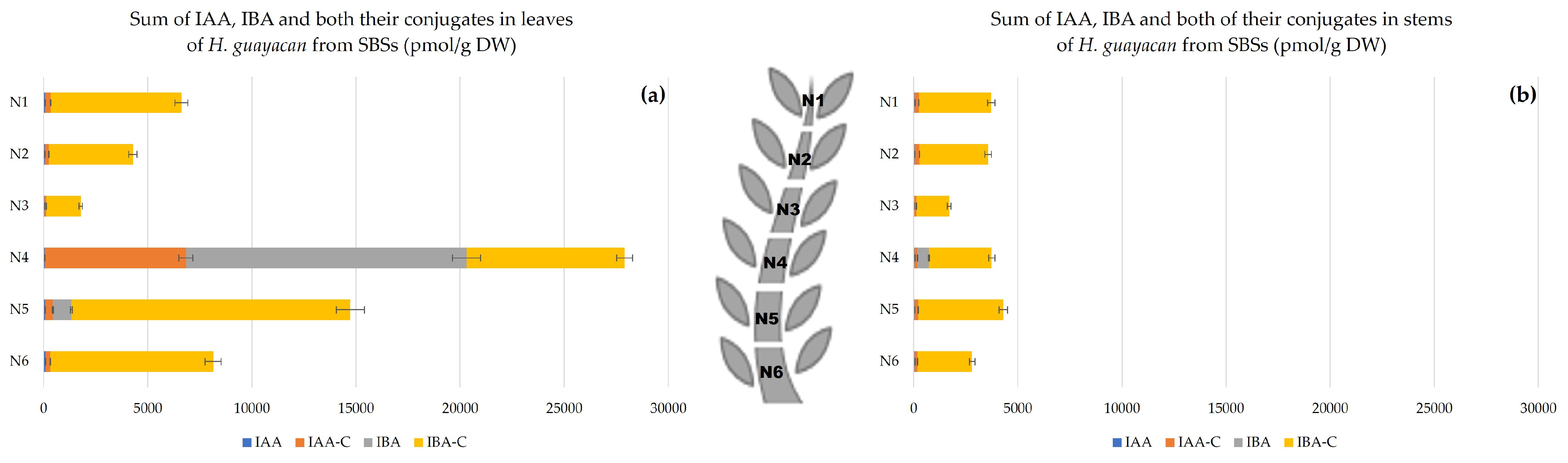
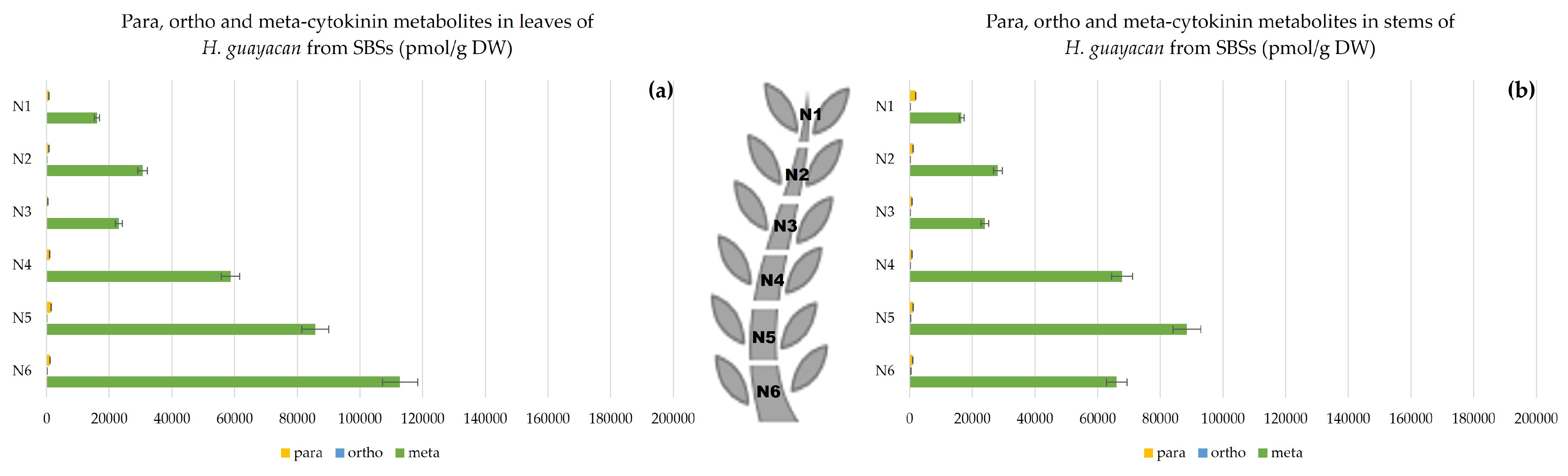
2.2.3. Shoots Originating from the Basal Section (SBSs)
- Aromatic Cytokinin Free Bases and Riboside Isomers;
- Topolin-O-Glucosides;
- Aromatic Cytokinin N-Glucosides and pTR;
- Auxins.
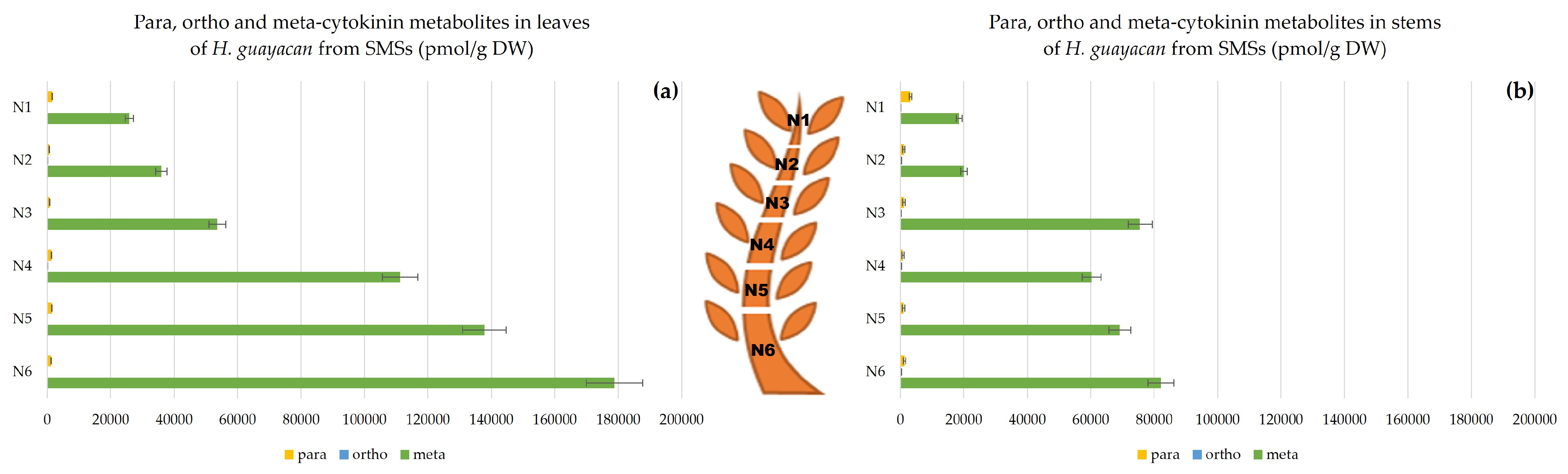
2.3. Meta, Ortho and Para Isomers of CK Metabolites
3. Discussion
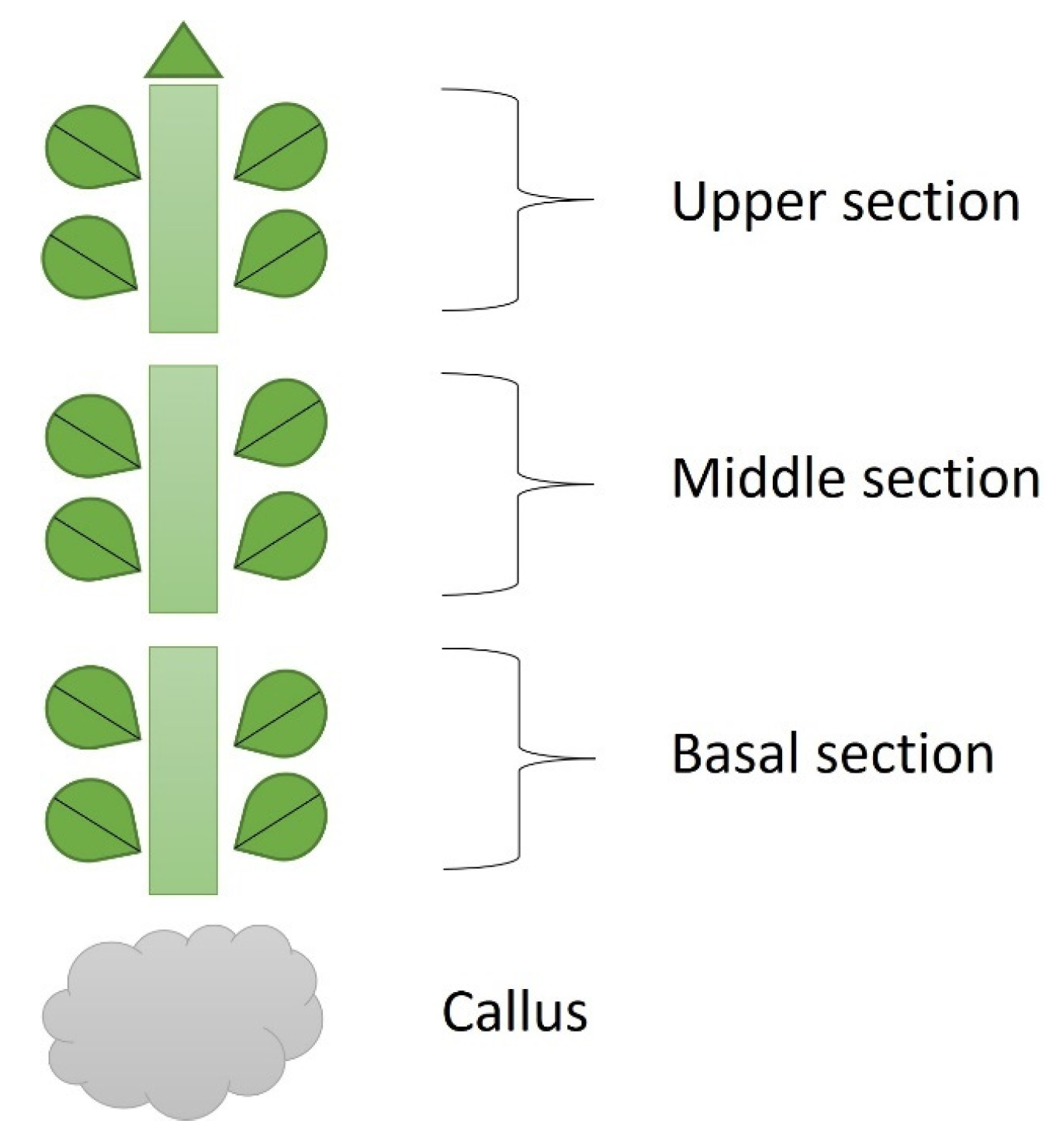
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wendling, I.; Brooks, P.R.; Trueman, S.J. Topophysis in Corymbia torelliana × C. citriodora seedlings: Adventitious rooting capacity, stem anatomy, and auxin and abscisic acid concentrations. New For. 2014, 46, 107–120. [Google Scholar] [CrossRef]
- Osugi, A.; Sakakibara, H. Q&A: How do plants respond to cytokinins and what is their importance? BMC Biol. 2015, 13, 1–10. [Google Scholar] [CrossRef]
- Kieber, J.J.; Schaller, G.E. Cytokinin signaling in plant development. Development 2018, 145, dev149344. [Google Scholar] [CrossRef]
- Mok, D.W.; Mok, M.C. Cytokinin metabolism and action. Annu. Rev. Plant Biol. 2001, 52, 89–118. [Google Scholar] [CrossRef]
- Strnad, M. The aromatic cytokinins. Physiol. Plant. 1997, 101, 674–688. [Google Scholar] [CrossRef]
- Lomin, S.N.; Krivosheev, D.M.; Steklov, M.Y.; Arkhipov, D.V.; Osolodkin, D.I.; Schmülling, T.; Romanov, G.A. Plant membrane assays with cytokinin receptors underpin the unique role of free cytokinin bases as biologically active ligands. J. Exp. Bot. 2015, 66, 1851–1863. [Google Scholar] [CrossRef] [PubMed]
- Hošek, P.; Hoyerová, K.; Kiran, N.S.; Dobrev, P.I.; Zahajská, L.; Filepová, R.; Kamínek, M. Distinct metabolism of N-glucosides of isopentenyladenine and trans-zeatin determines cytokinin metabolic spectrum in Arabidopsis. New Phytol. 2020, 225, 2423–2438. [Google Scholar] [CrossRef]
- Bajguz, A.; Piotrowska, A. Conjugates of auxin and cytokinin. Phytochemistry 2009, 70, 957–969. [Google Scholar] [CrossRef]
- Hou, B.; Lim, E.-K.; Higgins, G.S.; Bowles, D.J. N-Glucosylation of Cytokinins by Glycosyltransferases of Arabidopsis thaliana. J. Biol. Chem. 2004, 279, 47822–47832. [Google Scholar] [CrossRef] [PubMed]
- Bairu, M.W.; Stirk, W.A.; Dolezal, K.; Van Staden, J. Optimizing the micropropagation protocol for the endangered Aloe polyphylla: Can meta-topolin and its derivatives serve as replacement for benzyladenine and zeatin? Plant Cell Tissue Organ Cult. (PCTOC) 2007, 90, 15–23. [Google Scholar] [CrossRef]
- Werbrouck, S.P.O. Merits and drawbacks of new aromatic cytokinins in plant tissue culture. In Proceedings of the IV International Symposium on Acclimatization and Establishment of Micropropagated Plants, Bangalore, India, 8 December 2008; Volume 865, pp. 103–107. [Google Scholar]
- Smulders, M.J.M.; de Klerk, G.J. Epigenetics in plant tissue culture. Plant Growth Regul. 2011, 63, 137–146. [Google Scholar] [CrossRef]
- Magyar-Tábori, K.; Dobránszki, J.; da Silva, J.A.T.; Bulley, S.M.; Hudák, I. The role of cytokinins in shoot organogenesis in apple. Plant Cell Tissue Organ Cult. (PCTOC) 2010, 101, 251–267. [Google Scholar] [CrossRef]
- Gentile, A.; Jàquez Gutiérrez, M.; Martinez, J.; Frattarelli, A.; Nota, P.; Caboni, E. Effect of meta-Topolin on micro-propagation and adventitious shoot regeneration in Prunus rootstocks. Plant Cell Tissue Organ Cult. (PCTOC) 2014, 118, 373–381. [Google Scholar] [CrossRef]
- Nisler, J. Cytokinin Properties of Meta-Topolin and Related Compounds. Meta-Topolin A Growth Regul. Plant Biotechnol. Agric. 2021, 23–30. [Google Scholar] [CrossRef]
- Burley, J.T.; Kellner, J.R.; Hubbell, S.P.; Faircloth, B.C. Genome assemblies for two Neotropical trees: Jacaranda copaia and Handroanthus guayacan. G3 Genes Genomes Genetics 2021, 11, jkab010. [Google Scholar] [CrossRef] [PubMed]
- Grose, S.O.; Olmstead, R.G. Taxonomic revisions in the polyphyletic genus Tabebuia s. I. (Bignoniaceae). Syst. Bot. 2007, 32, 660–670. [Google Scholar] [CrossRef]
- Martín-Seijo, M.; Kaal, J.; Tórne, C.M.; Torné, J.M. Wood in pre-Columbian funerary Rituals: A case study from El Caño (Panama, AD 880–1020). Environ. Archaeol. 2021, 26, 406–422. [Google Scholar] [CrossRef]
- Available online: www.iucnredlist.org (accessed on 21 May 2020).
- Augspurger, C.K.; Wilkinson, H.T. Host Specificity of Pathogenic Pythium Species: Implications for Tree Species Diversity. Biotropica 2007, 39, 702–708. [Google Scholar] [CrossRef]
- Manners, G.D.; Jurd, L.; Wong, R.; Palmer, K. Constituents of Tabebuia guayacan: The structure of guayacanin. Tetrahedron 1975, 31, 3019–3024. [Google Scholar] [CrossRef]
- Harwood, J.; Tribaldos, W.; Lobo, S.; Wcislo, W. Wood Identification of The Altarpiece and a Sculpture of The Ca-thedral Basilica Santa María La Antigua. Eur. J. Sci. Theol. 2021, 17, 137–149. [Google Scholar]
- Keller, R. Bignoniaceae to Buxaceae. In A Field Guide to Tropical Plant Families; Springer International Publishing: Cham, Switzerland, 2023; pp. 173–184. [Google Scholar]
- Power, A.B.; Dodd, R.S.; Libby, W.J. Cyclophysis and topophysis in coast redwood stecklings. Silvae Genetica 1988, 37, 8–14. [Google Scholar]
- Dransfield, J.; Halle, F.; Oldeman, R.A.A.; Tomlinson, P.B. Tropical Trees and Forests. An Architectural Analysis. J. Ecol. 1980, 68, 340. [Google Scholar] [CrossRef]
- Peer, K.R.; Greenwood, M.S. Maturation, topophysis and other factors in relation to rooting in Larix. Tree Physiol. 2001, 21, 267–272. [Google Scholar] [CrossRef]
- Hung, C.D.; Trueman, S.J. Topophysic effects differ between node and organogenic cultures of the eucalypt Corymbia torelliana × C. citriodora. Plant Cell Tissue Organ Cult. (PCTOC) 2011, 104, 69–77. [Google Scholar] [CrossRef]
- Pitekelabou, R.; Aidam, A.V.; Kokou, K. In vitro micropropagation of Nauclea diderrichii (de wild &t. Durand) Merrill: Effect of nodes position on plantlets growth and rooting. Eur. Sci. J. 2015, 11. Available online: https://core.ac.uk/download/pdf/236415441.pdf (accessed on 31 July 2015).
- Zalewska, M.; Miler, N.; Wenda-Piesik, A. Effect of in vitro topophysis on the growth, development, and rooting of chrysanthemum explants (Chrysanthemum grandiflorum/Ramat./Kitam). J. Hortic. Sci. Biotechnol. 2010, 85, 362–366. [Google Scholar] [CrossRef]
- Holub, J.; Hanuš, J.; Hanke, D.E.; Strnad, M. Biological activity of cytokinins derived from ortho-and me-ta-hydroxybenzyladenine. Plant Growth Regul. 1998, 26, 109–115. [Google Scholar] [CrossRef]
- Aremu, A.O.; Plačková, L.; Bairu, M.W.; Novák, O.; Szüčová, L.; Doležal, K.; Finnie, J.F.; Van Staden, J. Endogenous cytokinin profiles of tissue-cultured and acclimatized ‘Williams’ bananas subjected to different aromatic cytokinin treatments. Plant Sci. 2014, 214, 88–98. [Google Scholar] [CrossRef]
- Uzelac, B.; Janošević, D.; Simonović, A.; Motyka, V.; Dobrev, P.I.; Budimir, S. Characterization of natural leaf senescence in tobacco (Nicotiana tabacum) plants grown in vitro. Protoplasma 2016, 253, 259–275. [Google Scholar] [CrossRef]
- Hoyerová, K.; Hošek, P. New Insights into the Metabolism and Role of Cytokinin N-Glucosides in Plants. Front. Plant Sci. 2020, 11, 541514. [Google Scholar] [CrossRef]
- Doležal, K.; Popa, I.; Hauserová, E.; Spíchal, L.; Chakrabarty, K.; Novák, O.; Kryštof, V.; Voller, J.; Holub, J.; Strnad, M. Preparation, biological activity and endogenous occurrence of N6-benzyladenosines. Bioorganic Med. Chem. 2007, 15, 3737–3747. [Google Scholar] [CrossRef]
- Abdouli, D.; Plačková, L.; Doležal, K.; Bettaieb, T.; Werbrouck, S.P. Topolin cytokinins enhanced shoot proliferation, reduced hyperhydricity and altered cytokinin metabolism in Pistacia vera L. seedling explants. Plant Sci. 2022, 322, 111360. [Google Scholar] [CrossRef] [PubMed]
- Spena, A.; Prinsen, E.; Fladung, M.; Schulze, S.C.; Van Onckelen, H. The indoleacetic acid-lysine synthetase gene of Pseudomonas syringae subsp. savastanoi induces developmental alterations in transgenic tobacco and potato plants. Mol. Genet. Genom. 1991, 227, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Reemmer, J.; Murphy, A. Intercellular transport of auxin. In Auxin and Its Role in Plant Development; Springer: Vienna, Austria, 2014; pp. 75–100. [Google Scholar]
- Werbrouck, S.P.O.; Strnad, M.; Van Onckelen, H.A.; Debergh, P.C. Meta-topolin, an alternative to benzyladenine in tissue culture? Physiol. Plant. 2008, 98, 291–297. [Google Scholar] [CrossRef]
- Pokorná, E.; Hluska, T.; Galuszka, P.; Hallmark, H.T.; Dobrev, P.I.; Drábková, L.Z.; Filipi, T.; Holubová, K.; Plíhal, O.; Rashotte, A.M.; et al. Cytokinin N-glucosides: Occurrence, Metabolism and Biological Activities in Plants. Biomolecules 2020, 11, 24. [Google Scholar] [CrossRef]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Shoots | Length of Shoots (cm) | Number of Nodes/Shoot | Number of Nodes/Plant | Weight of Callus (g) | Length of Internodes (cm) | |
|---|---|---|---|---|---|---|
| Upper section | 1.00 ± 0.0 a | 2.64 ± 1.1 a | 5.35 ± 1.5 c | 5.35 ± 1.5 a | 0.38 ± 0.2 a | 0.51 ± 0.2 a |
| Middle section | 3.80 ± 1.3 c | 3.97 ± 1.1 b | 4.88 ± 1.5 b | 18.86 ± 6.5 c | 0.36 ± 0.2 a | 0.84 ± 0.2 c |
| Basal section | 2.53 ± 0.8 b | 2.67 ± 1.2 a | 3.94 ± 1.6 a | 10.15 ± 4.5 b | 0.51 ± 0.3 b | 0.72 ± 0.3 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grira, M.; Prinsen, E.; Werbrouck, S.P.O. The Effect of Topophysis on the In Vitro Development of Handroanthus guayacan and on Its Metabolism of Meta-Topolin Riboside. Plants 2023, 12, 3577. https://doi.org/10.3390/plants12203577
Grira M, Prinsen E, Werbrouck SPO. The Effect of Topophysis on the In Vitro Development of Handroanthus guayacan and on Its Metabolism of Meta-Topolin Riboside. Plants. 2023; 12(20):3577. https://doi.org/10.3390/plants12203577
Chicago/Turabian StyleGrira, Maroua, Els Prinsen, and Stefaan P. O. Werbrouck. 2023. "The Effect of Topophysis on the In Vitro Development of Handroanthus guayacan and on Its Metabolism of Meta-Topolin Riboside" Plants 12, no. 20: 3577. https://doi.org/10.3390/plants12203577