1. Introduction
Holm oak (
Quercus ilex L.), an evergreen oak, is distributed from Portugal eastward to Syria, and from Morocco and Algeria north toward France. It is an important forest tree and the main species associated with the
dehesa or
montado, agrosilvopastoral systems of southwestern Europe. Well-adapted to drought, this species is often the dominant type of vegetation in areas of transition between wet and dry climates [
1]. However, longer droughts and the hotter temperatures now being experienced are having negative effects, including severe defoliation, attack by pathogens, and increased mortality [
2,
3], a scenario likely to become even worse as climate change advances.
The main pathogen associated with oak decline is the oomycete
Phytophthora cinnamomi Rands [
4,
5,
6]. This soil-borne microorganism causes root rot, bark cankers, the drying of twigs and branches, and ultimately death. The symptoms are similar to those caused by drought, high temperatures, or rotting of the root system. Nevertheless, the oomycete requires free water in the soil to sporulate and infect the plant. The disease tends to spread in a downward direction due to the movement of the causal agent with water [
7].
Not all holm oaks are, however, equally affected. Some trees are more tolerant to
Phytophthora cinnamomi and drought stresses, a consequence of genetic diversity between provenances and families within provenances [
8,
9]. The selection of those with the appropriate genetic background is necessary for use in traditional tree breeding programs [
10,
11]. Another way to address the increasing mortality problem is the vegetative propagation of tolerant plants to
P. cinnamomi infection [
12]. Micropropagation by somatic embryogenesis allows rejuvenated plants to be obtained with all the genetic potential of the originally selected trees. Furthermore, the clones produced can be used to study the responses of different genotypes to infection, and thus allow the most tolerant to be identified, along with the genes and proteins involved [
13]. The genetic transformation of somatic embryos using a Gnk2-like protein may also allow genotypes with improved anti-oomycete activity to be obtained [
14]. The induction of epigenetic changes on somatic embryos elicits a defense response and provides another opportunity to obtain tolerant holm oak trees [
15,
16]. There is increasing evidence that following exposure to an environmental stimulus, different genotypes acquire some kind of epigenetic memory that determines new behavior when restimulated. Stress exposure in plants leads to many epigenetic changes routed through mechanisms such as chromatin remodeling, DNA methylation, histone modification, and non-coding RNAs. Epigenetic markers are known to be transferred from plants to their offspring [
17,
18]. These epigenetic mechanisms can be used for developing resilient varieties [
19].
Plants have many types of innate immune responses. These are often short-term, but infection may elicit acquired immunity by priming inducible defenses. This type of resistance is referred to as systemic acquired resistance (SAR), and plants are primed more quickly and effectively to adopt a defense response to the next pathogen attack [
20]. Although some priming states disappear within a few days, others can even be transmitted to subsequent generations [
21]. Indeed, transgenerational-induced resistance to
P. cinnamomi occurs naturally in some holm oak trees [
22] via maternally experienced stimuli at the time of seed formation, leading to epigenetic reprogramming [
23,
24]. The large number of genetically identical embryos that can be obtained by somatic embryogenesis [
25] offers the opportunity to expose many initial explants to such priming. If the immune memory thus obtained can be maintained over development, primed seed could be produced for the production of pathogen-tolerant trees [
26,
27].
P. cinnamomi oomycetes secrete elicitins during the colonization process [
28]. These proteins induce immediate defense responses in the host plant via hypersensitivity reactions. Cinnamomin, the elicitin used in this research, is one of the substances that have been proven to cause a defense response in oak plants [
29,
30]. Similar reactions may occur in response to other natural and chemically synthesized compounds, including β-aminobutyric acid (BABA), salicylic acid (SA), methyl-jasmonate (MeJA), and benzothiadiazole (or acibenzolar) (BTH), among others. The response induced depends on the species in question, its genotype, the type of elicitor used, its concentration, and the application time. The external application of these compounds may induce the accumulation of active oxygen species, phytoalexin biosynthesis, the activation of different defense-related enzymes, the production of phenolic compounds, or the appearance of pathogenesis-related proteins, some of which possess antimicrobial properties [
31]. Primed plants do not lose their constitutive degree of defense but may gain greater resistance via the above treatment [
18].
Different in vitro selection techniques for improving plant resistance are available for use with woody species [
32,
33,
34,
35]. Confronting host plant tissue with a fungal pathogen in dual culture has been used to assess the resistance of plant genotypes [
36,
37,
38]. When the tissues of selected lines inhibit the growth of a mycelium, the explanation may lie in their production of externally exported resistance compounds. Dual culture assays have confirmed the chemotactic attraction of
P. cinnamomi mycelia toward holm oak somatic embryos to be dependent on the latter’s genotype [
15,
16]. The same has also been observed in vitro with shoots and leaves [
13] and in roots during the infection process in vivo [
39]. The present work describes somatic embryo treatments by chemical elicitors or filtrates of
P. cinnamomi oomycete to elicit a defense response that is maintained after embryo germination, increasing the likelihood of producing disease-tolerant trees.
3. Discussion
When somatic embryos are forming by recurrent or cleavage embryogenesis under controlled conditions, they can enter a state suitable for introducing epigenetic markers. Kvaalen and Johnsen [
40] first showed this in forest trees when they induced the formation of different epitypes of
Picea abies by culturing embryogenic lines at different temperatures. Epigenetic changes in somatic embryos have been verified in other plant species, such as
Pinus radiate and
Pinus pinaster [
41,
42]. In the present study, the epitypes of holm oak could have been produced by the elicitation of somatic embryogenic lines. This was verified when somatic embryos of the same genotype behaved differently in dual culture with
P. cinnamomi mycelium, depending on the elicitation treatment to which they had been subjected. Elicitation with 50 µM or 100 µM of BTH, BABA, MeJA, SA, or filtrate extracts at 10% or 30% (FILT10 or FILT30) for 3 or 60 days made the embryos show differential growth of the mycelium (DGM values) and the progression of necrosis with respect to untreated embryos. The genotype, elicitor, and elicitation time had an important effect on these results. A similar study with the same species could not find significant differences among elicitation treatments with filtered extracts and p-aminobenzoic acid (PABA), MeJA, and BTH at 5, 10, 25, and 50 μM [
15]. However, in that report, only one holm oak genotype was used, and our results showed a high dependence on the genotype (
p < 0.001) for DGM values.
Several elicitor-binding sites have been identified in cell plasma membranes and have investigated the general mechanism of action of elicitors [
43]. Elicitors act as signal compounds at low concentrations, and it is important to know the application time for effective elicitation without possible toxicity or plant production alteration. No significant differences were seen in terms of biomass multiplication when using either the 50 or 100 μM concentrations of the chemical elicitors. However, the number of embryos produced was dependent on treatment and genotype. Embryos were necrotic after 60-day elicitation with 50 or 100 μM BTH. The same was previously reported when a concentration of 25 μM was used [
15]. Long elicitation with BTH also prevented the germination of the few non-necrotic embryos produced. Embryo production of the genotype E2 was also particularly affected during the 3-day elicitation process, which may be due to stress caused by the agitation used [
44], although it was not affected by treatment with MeJA. The exogenous application of MeJA improves growth and affects the levels of endogenous hormones, as well as the physiological and biochemical characteristics of stressed plants [
15,
45,
46]. High levels of jasmonic acid (JA) can also stimulate the accumulation of indoleacetic acid and, therefore, the production of somatic embryos [
47].
3.1. Differential Growth Measurements
The stimulation or inhibition of mycelium growth can allow the selection at the embryonic level of genotypes tolerant to infection [
36]. In our dual assays, the mycelium grew toward both the embryos and their derived roots, suggesting it to be chemically attracted toward them. The DGM value was dependent on the genotype. It has been described that the constitutive levels of total phenols and condensed tannins may act as chemical defenses in holm oak, with variation between provenance and genotypes [
48,
49].
The development of dual cultures involving
P. cinnamomi mycelium and elicited embryos or roots can provide the basis for the evaluation of tolerance of epitypes formed. Elicitation for tolerance was associated with lower DGM values and depended on the genotype and elicitation treatment but not the concentration of elicitor used. The recorded DGM values indicated that compared to the controls, the elicited somatic embryos inhibited the growth of the mycelium. With BABA treatments, significant differences in terms of less DGM were seen in all genotypes. BABA is a nonprotein amino acid that has been previously linked to priming mechanisms against
Phytophthora sp. [
50]. It is also known to induce defense responses in plants after treating their seeds. Certainly, treating tomato seedlings with BABA results in characteristic genome-wide changes in DNA methylation [
51,
52].
In dual culture assays involving the roots of elicited embryos, the growth of the mycelium was also reduced (lower DGM values) with respect to non-elicited controls. The present results confirm that the elicitation persists with embryo-derived roots. The elicitation of embryogenic lines with BABA also inhibited mycelial growth in root dual assays. The response to BABA is a popular model for studying the molecular signaling underpinning priming. This BABA-induced resistance (BABA-IR) is based on the priming of SA-dependent and independent defenses and is reported to provide broad protection against biotrophic and necrotrophic pathogens and abiotic stresses. So, it was confirmed to induce broad-spectrum resistance with epigenetic regulation and the long-term maintenance of priming [
53,
54,
55].
Long treatment with salicylic acid (SA) or filtrate of a cinnamomin elicitin at 10% (FILT10) also reduced the growth of mycelium with roots. SA is a defense hormone required for both local and systemic acquired resistance in plants (SAR). The exogenous application of SA to induce resistance to pathogens through various pathways has been described [
31,
56]. Elicitins are also described as extracellular sterol-binding proteins with specific signaling responses in the host plant, and they have been widely used for in vitro selection of resistant plants [
28,
32,
33,
34]. Root exudates, like phytoanticipins, are produced prior to biotic stress against
Phytophthora cinnamomi [
57]. The inhibition of the mycelium could be justified by secreted compounds.
3.2. Progress of Necrosis
The correlation between the progress of necrosis and linear colonization has been suggested as a measure of tolerance. The root necrosis seen at 24 h and 48 h could easily reflect the degree of tolerance of plant tissues [
58,
59]. In our dual culture assays, long-term elicitation with FILT10 reduced necrosis in embryos at 48 h and in the roots at 24 h and 48 h of all genotypes compared to non-elicited controls. Some studies had already confirmed with quantitative real-time PCR a significant decrease in pathogen colonization of
Quercus sp. roots after 24 h of pre-treatment with α- and β-cinnamomin [
30]. In addition, great progress has been made in fungal elicitor-triggered plant immunity, especially in the signaling pathways of PTI (pathogen-associated molecular pattern-triggered immunity) and ETI (effector-triggered immunity) [
60]. Short-term elicitation with BABA also reduced at 24 h and 48 h the progress of necrosis in embryos and roots. Although large differences were observed with respect to the genotype, the treatments coincided with the DGM inhibition results.
3.3. Tolerant Epitypes of Elicited Holm Oak Somatic Embryos Could Be Revealed by Challenges in Dual Culture with Phytophthora cinnamomi
The inhibition of the mycelium growth or the reduction in the progression of necrosis by tissues of the produced epitypes could reveal their tolerance to
P. cinnamomi [
36,
37,
38]. Tolerance is defined as the ability of the plant to mitigate the negative effects caused by the pathogen, despite an insignificant reduction in the presence and spread of the pathogen [
57]. Somatic embryos from tolerant trees reduced mycelial growth and necrosis in dual culture respect population control. However, roots from tolerant trees attracted or increased mycelial growth, and they had high necrosis. Previously, other authors carried out a similar test with shoots and leaves from somatic embryos of tolerant holm oaks [
13]. Shoots of one tolerant genotype but leaves of another tolerant genotype inhibited the growth of mycelium. Different responses were suggested by different phenolic content. Subsequently, results in dual culture with elicited embryos were compared with those in naturally tolerant trees. The short-term elicitated embryos with SA and FILT10 had the same behavior as tree-tolerant embryos.
Plant cells respond to
Phytophthora cinnamomi infection by multiple defense actions. Within the same species and between genotypes, plant cells can display a slightly different defense response [
57,
61]. In dual cultures with pathogens, plant cells (embryos and roots) could also launch several signals during defense reactions. The first is the recognition of pathogen-associated molecular patterns by membrane receptors, leading to the activation of the innate immune response. Defenses could include antimicrobial compounds (less DGM) such as phenolic compounds, saponins, proteins and glucosinolates, and physical barriers to stop the progress of the pathogen within the host tissue (less necrosis). With roots, exudates may be the first line of defense against pathogens acting as inhibitors or attractant molecules (more DGM). Inhibitors were found in resistant but not moderately resistant avocado rootstocks [
57]. To take roots fast enough was another strategy observed in tolerant genotypes [
61]. If pathogens are recognized, plant cells can also be protected by SAR, with the activation of the hypersensitive response [
62] characterized by local cell death (more necrosis).
Micropropagated clonal plants allow for minimizing differences in the analysis of plant defense [
63]. Although it would be necessary to perform other advanced analyses to interpret the results, priming holm oak embryos for 3 and 60 days with 50 µM BABA or for 60 days with oomycete filtrates at 10% induced some tolerance (in a genotype-dependent fashion) to
P. cinnamomi infection. Dual cultures could be the first simple procedure for testing holm oak tolerance. The inhibition of mycelium growth and necrosis can give useful information to select efficient elicitation treatments to produce disease-tolerant trees.
4. Materials and Methods
4.1. Somatic Embryos
Three holm oak genotypes were micropropagated for embryo elicitation: Q8 from Mora (Toledo, Spain) and E00 and E2 from the
Finca El Encín in Alcalá de Henares (Madrid, Spain). Somatic embryogenesis was induced using teguments from developing ovules isolated from acorns [
25]. For this, immature acorns were collected from adult trees in July, surface-sterilized with sodium hypochlorite (15%) and 1 drop of Tween
®20 for 10 min, and then subjected to three washes in sterile water. At least 100 ovules/genotypes of 5–6 mm in length were excised and cultured in the dark at 23 °C on a proliferation medium without plant growth regulators. This medium contained the macronutrients of the Schenk and Hildebrandt medium [
64], the micronutrients and vitamins of the Murashigue and Skoog medium [
65], 30 g/L sucrose, and 0.6% agar (S1000, B&V, Parma, Italy). The pH was adjusted to 5.75 before agar addition, and the medium was autoclaved at 121 °C for 30 min. Developed zygotic embryos were extracted during the first month of cultivation. The teguments were then cultured on the same proliferation medium with monthly subculturing under a 16 h light photoperiod using a Sylvana GRO-LUX
® lighting system plus a Philips Cool-White system (120 μmol m
−2 s
−1). Induced somatic embryos were obtained in the third month of culture on the surface and cut areas of the teguments. Embryogenic lines were amplified by recurrent embryogenesis with monthly subcultures until elicitation assays could be started. Somatic embryos isolated from each culture were matured for 1 month on the proliferation medium containing 7 g/L of activated charcoal and then moved at 4 °C for 8 weeks in the dark. Embryos germinated on a fresh proliferation medium without plant growth regulators under a light photoperiod of 16 h at 23 °C.
Somatic embryos (without elicitation) were also obtained from selected holm oaks located in a severely affected area in Plasencia, Extremadura, Spain (39°58′01.6″ N–6°05′33.9″ W). Two asymptomatic genotypes were selected as tolerant trees (T), where the disease caused by
P. cinnamomi produced high mortality in the rest [
12] and other two genotypes were used as population controls (P) where
P. cinnamomi was not present.
4.2. Elicitation Media
Somatic embryos were induced using different elicitors: methyl jasmonate (MeJA), salicylic acid (SA), β-aminobutyric acid (BABA), or benzothiadiazole (BTH) at a concentration of either 50 µM or 100 µM and an oomycete culture filtrate—a cinnamomin-inducing liquid medium—in which the strain of
P. cinnamomi was cultured [
29] and diluted to 10% or 30% (
v/
v) (FILT10 and FILT30, respectively).
For the oomycete culture filtrate, a virulent strain of
P. cinnamomi (UEX1), donated by the University of Extremadura (Plasencia, Spain), was used. This had been isolated from a stand of diseased trees [
66] and was maintained on a PDA agar medium (20 g/L Potato Dextrosa Agar and 5 g/L fresh agar) in darkness at 4 °C. Active cultures were obtained by taking 10 × 10 mm agar plugs containing mycelium and placing them in the center of Petri dishes containing a fresh PDA agar medium. These were cultured at 23 °C for one week in the dark. The oomycete filtrate treatments were based on those reported by Ganesan and Jayabalan (2006) [
32]. First, an elicitin secretion medium (ESM) was prepared using 0.5 g/L KH
2PO
4, 0.25 g/L MgSO
4.7H
2O, 1 g/L asparagine, 1 mg/L thiamine, 0.5 g/L yeast extract, and 20 g/L glucose. This was sterilized by filtration through a 22 µm pore-size membranes. Then, 200 mL Erlenmeyer flasks containing 40 mL of ESM medium were inoculated with eight agar plugs (10 × 10 mm surface area) from active
P. cinnamomi cultures on a semi-solid PDA medium. These were cultured under agitation (50 rpm) in darkness at 23 °C for five days before filtering the medium through a Büchner funnel. The filtrate was used the same day or stored at 4 °C (no more than 72 h). FILT10 and FILT30 were prepared by diluting accordingly with a proliferation medium.
4.3. Elicitation Treatments
Somatic embryos of each genotype were elicited via exposure to elicitors for either 60 days (long elicitation) or 3 days (short-term elicitation). Long elicitation involved culturing 0.3 g of embryogenic clumps for 60 days on a semi-solid proliferation medium supplemented with either 50 or 100 μM of each of the chemical elicitors (separately) or with the FILT10 or FILT30 preparations (replicated in quintuplicate). Incubation was then allowed at 23 °C with a photoperiod of 16 h of light. After the first month in culture, all the embryogenic material produced was cultured in new vessels, again using 0.3 g of tissue.
For short-term elicitation, 0.3 g of embryogenic clumps grown on a semi-solid proliferation medium were cultured for 3 days in a proliferation liquid medium supplemented with one of the elicitors. Five 200 mL Erlenmeyer flasks, containing 50 mL of the elicitation medium, were inoculated with embryogenic tissue, and the cultures were grown with agitation at 110 rpm for 3 days at 23 °C under a 16 h photoperiod. All embryogenic material from each flask was then filtered through nylon filters with a 40 µm pore size and transferred to a semi-solid proliferation medium without elicitors, which was then cultured for 60 days with one subculture at 30 days.
The monthly biomass multiplication rate (MR) was calculated as the number of new vessels needed to culture all the tissue produced by each treatment divided by the initial number of vessels (i.e., 5). The number of single somatic embryos produced per vessel (SE) was counted at the end of the 60-day period for each vessel, genotype and elicitor treatment, and means calculated. For both elicitation times, non-treatment (NT) controls were prepared in the same way, but without elicitors. Well-formed embryos were matured for 1 month and then were kept cold (4 °C) for 8 weeks, and they were then germinated according to a previously described protocol. The conversion rate (embryos with stem and root) was determined for each treatment.
4.4. Dual Culture Assays with P. cinnamomi
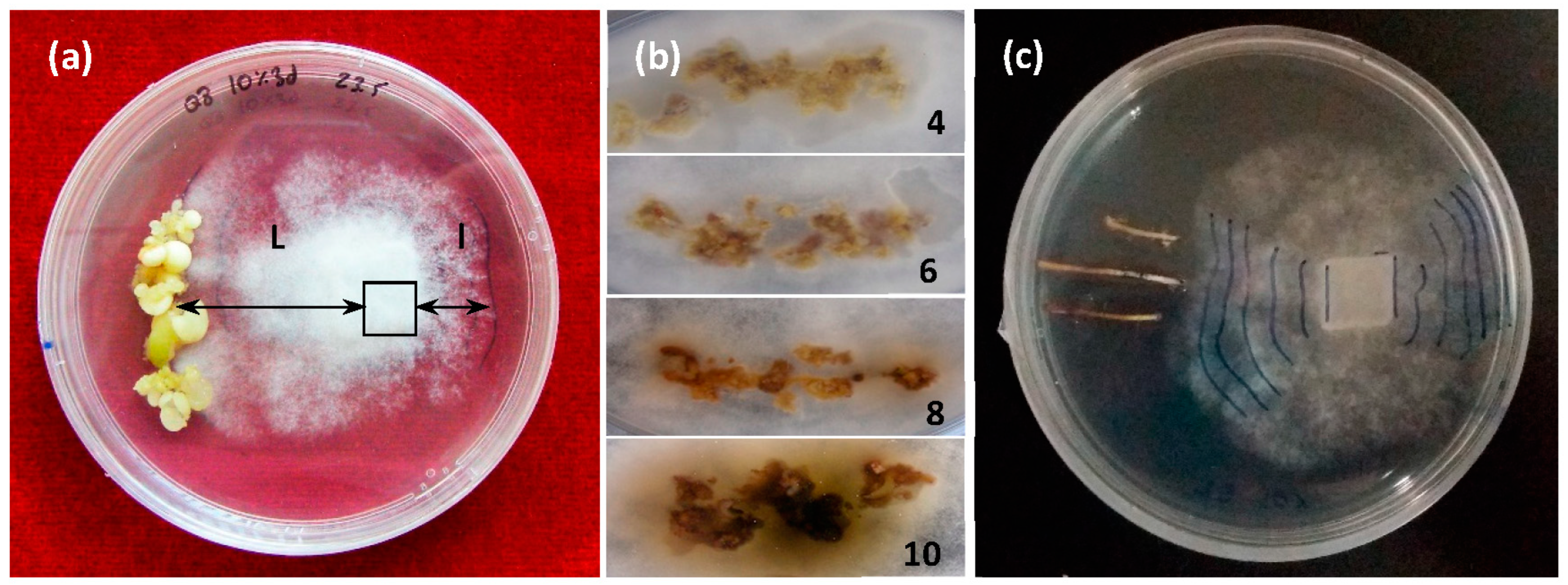
Elicited somatic embryos (about 500 mg) of each genotype and treatment were washed and placed on one side of a 90 mm Petri dish containing a 20 mL proliferation medium. A 10 × 10 mm plug of a PDA agar medium containing actively growing mycelium was placed on the opposite side, a quarter of the dish’s diameter from the respective edge. Assays involving somatic embryos of each genotype that did not undergo elicitation (NT) and cultures with only the mycelium were prepared as the controls. All tests were performed with 10 replicates. Once prepared, the assay plates were incubated at 23 °C in darkness for 10 days. Differential growth of the mycelium (DGM) was determined as the growth of the mycelium toward the somatic embryos (L) minus its growth in the opposite direction (l). Data were collected daily from day 2 to day 10. When the mycelium reached the embryos, the progress of necrosis was measured daily, according to a visual scale (
Figure 3).
Similar assays were performed using three root tips (30 mm long, 2 mm wide) derived from the somatic embryos of the same genotypes (involving all elicitation treatments) that were allowed to germinate. The advance of the mycelium toward the roots was noted daily from day 2 to day 10, and DGM values were determined above. Lesion lengths were measured 24 h and 48 h after the mycelium reached each root (0 h).
4.5. Comparison of Elicited and Tolerant Genotypes When Challenged to Oomycete
Somatic embryos obtained from the naturally tolerant and control trees were grown in dual culture. DGM values on days 3, 4, and 5 with embryos and roots from holm oak somatic embryos were recorded in dual cultures with 20 repetitions per genotype. The progression of necrosis at 24 h and 48 h with embryos and roots was also recorded. Data were expressed as the mean value of 20 repetitions of tolerant genotypes, 20 repetitions of population genotype, and 6 repetitions for each genotype and elicitation treatment.
4.6. Statistics
Data recorded in the different experiments were subjected to analysis of variance using SPSS Statistica v.19 software (IBM Statistics). The normality was evaluated using the Shapiro–Wilk test, confirming the normal distribution of the data. The homogeneity of variance was assessed using the Levene test. According to the ANOVA results, when the F-ratio was significant, post hoc Duncan’s multiple range test analyses were performed to detect significant differences between genotypes and elicitors (p ≤ 0.05).



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




