

Subterranean Clover and Sulla as Valuable and Complementary Sources of Bioactive Compounds for Rainfed Mediterranean Farming Systems
 , , ,
, , ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Antioxidant Capacity and Phenolic Content
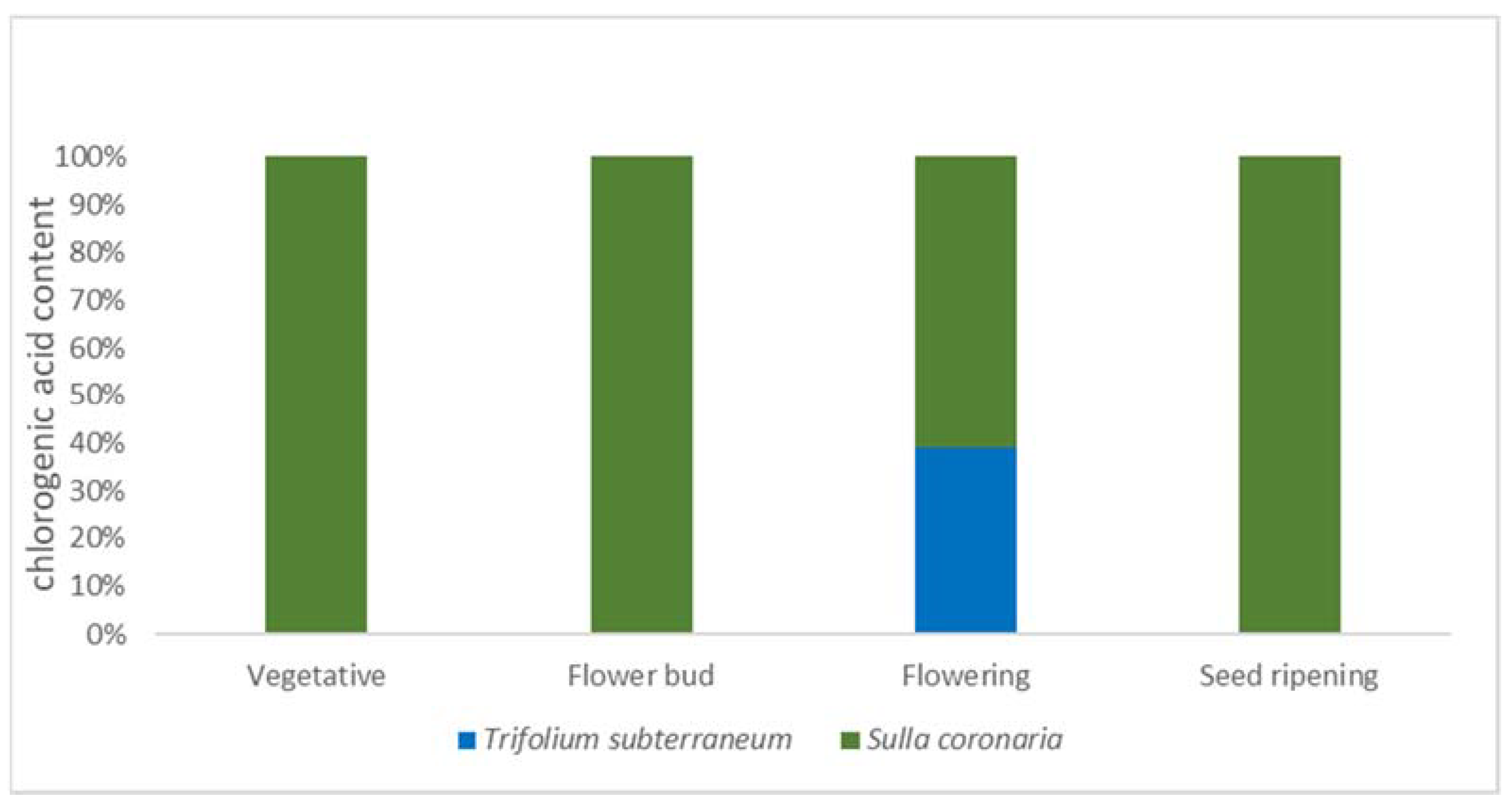
2.2. RP-HPLC Analysis of Phenolic Compounds
- -
- Three isomers of caffeoylquinic acid, i.e., neochlorogenic, chlorogenic, and cryptochlorogenic acids;
- -
- Caffeic acid and a hydroxycinnamic acid derivative (absorption spectrum identical to caffeic acid, quantified as caffeic acid equivalent);
- -
- Five flavanols, i.e., rutin, quercetin 3-galattoside, quercetin 3-glucoside, isorhamnetin 3-rutinoside, and one of its isomers (quantified as isorhamnetin 3-rutinoside equivalent);
- -
- Diosmin, a flavone glycoside of diosmetin.
3. Materials and Methods
3.1. Plant Material and Shoot Sampling
3.2. Antioxidant Capacity and Total Phenolic Content
3.3. RP-HPLC Analysis of Phenolic Compounds
3.4. Data Analyses
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rochon, J.J.; Doyle, C.J.; Greef, J.M.; Hopkins, A.; Molle, G.; Sitzia, M.; Scholefield, D.; Smith, C.J. Grazing legumes in Europe: A review of their status, management, benefits, research needs and future prospects. Grass Forage Sci. 2004, 59, 197–214. [Google Scholar] [CrossRef]
- Sulas, L. The future role of forage legumes in the Mediterranean climatic areas. In Grasslands: Developments Opportunities Perspectives; Reynolds, S.G., Frame, J., Eds.; FAO: Rome, Italy; Science Publishers: Enfield, NH, USA, 2005; pp. 29–54. ISBN 1-57808-359-1. [Google Scholar]
- Stagnari, F.; Maggio, A.; Galieni, A.; Pisante, M. Multiple benefits of legumes for agriculture sustainability: An overview. Chem. Biol. Technol. Agri. 2017, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Cocks, P.S.; Bennett, S.J. Introduction: Role of pasture and forage legumes in Mediterranean farming systems. In Genetic Resources of Mediterranean Pasture and Forage Legumes; Bennett, S.J., Cocks, P.S., Eds.; Kluwer Academic: Dordrecht, The Netherlands, 1999; pp. 9–19. [Google Scholar]
- Sulas, L.; Re, G.A.; Stangoni, A.P.; Ledda, L. Growing cycle of Hedysarum coronarium L. (sulla): Relationship between plant density, stem length, forage yield and phytomass partitioning. Cah. Options Méditerr. 2000, 45, 147–152. [Google Scholar]
- Muresu, R.; Porceddu, A.; Sulas, L.; Squartini, A. Nodule-associated microbiome diversity in wild populations of Sulla coronaria reveals clues on the relative importance of culturable rhizobial symbionts and co-infecting endophytes. Microbiol. Res. 2019, 221, 10–14. [Google Scholar] [CrossRef]
- Katznelson, J.; Morley, F.H.W. A taxonomic revision of sect. Calycomorphum of the genus Trifolium. I. The geocarpic species. Isr. J. Bot. 1965, 14, 112–134. [Google Scholar]
- Nichols, P.G.H.; Collins, W.J.; Barbetti, M.J. Registered Cultivars of Subterranean Clover—Their Characteristics, Origin and Identification, Bulletin No. 4327; Agriculture Western Australia: South Perth, Australia, 1996. Available online: https://researchlibrary.agric.wa.gov.au/cgi/viewcontent.cgi?article=1231&context=bulletins (accessed on 9 November 2022).
- Pecetti, L.; Piano, E. Variation of morphological and adaptive traits in subterranean clover populations from Sardinia (Italy). Genet. Res. Crop Evol. 2002, 49, 189–197. [Google Scholar] [CrossRef]
- Francis, C.M.; Gillespie, D.J. Ecology and distribution of subterranean clover and Medicago species in Sardinia. Aust. Plant Introd. Rev. 1981, 13, 15–25. [Google Scholar]
- Piano, E.; Sardara, M.; Pusceddu, S. Observations on the distribution and ecology of subterranean clover and other annual legumes in Sardinia. Riv. Agron. 1982, 12, 273–283. [Google Scholar]
- Gonzales Lopez, F.; Moreno Cruz, V.; Porqueddu, C.; Sulas, L. Recoleccion de Semillas de Leguminosas Anuales en Cerdena; Technical report; Centro de Investigaciones Científicas y Tecnológicas de Extremadura (CICYTEX): Badajoz, Spain, 1992; pp. 1–33. [Google Scholar]
- Abdi, A.I.; Nichols, P.G.H.; Kaur, P.; Wintle, B.J.; Erskine, W. Morphological diversity within a core collection of subterranean clover (Trifolium subterraneum L.): Lessons in pasture adaptation from the wild. PLoS ONE 2020, 15, e0223699. [Google Scholar] [CrossRef]
- Smetham, M.L. A review of subterranean clover (Trifolium subterraneum L.): Its ecology and use as a pasture legume in Australasia. Adv. Agron. 2003, 79, 303–350. [Google Scholar]
- Nichols, P.G.H.; Foster, K.J.; Piano, E.; Pecetti, L.; Kaur, P.; Ghamkhar, K. Genetic improvement of subterranean clover (Trifolium subterraneum L.). 1. Germplasm, traits and future prospects. Crop Past. Sci. 2013, 64, 312–346. [Google Scholar] [CrossRef]
- Nichols, P.G.H.; Jones, R.A.C.; Ridsdill-Smith, T.J.; Barbetti, M.J. Genetic improvement of subterranean clover (Trifolium subterraneum L.). 2. Breeding for disease and pest resistance. Crop Past. Sci. 2014, 65, 1207–1229. [Google Scholar] [CrossRef] [Green Version]
- Das, S.; Sharangi, A.B.; Egbuna, C.; Jeevanandam, J.; Ezzat, S.M.; Adetunji, C.O.; Onyeike, P.C. Health benefits of isoflavones found exclusively of plants of the fabaceae family. In Functional Foods and Nutraceuticals; Egbuna, C., Dable Tupas, G., Eds.; Springer: Cham, Switzerland, 2020; pp. 473–508. [Google Scholar]
- Křížová, L.; Dadáková, K.; Kašparovská, J.; Kašparovský, T. Isoflavones. Molecules 2019, 24, 1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, N.R. Permanent infertility in ewes exposed to plant oestrogens. Aust. Vet. J. 1990, 67, 197–205. [Google Scholar] [CrossRef]
- Adams, N.R. Detection of the effects of phytoestrogens on sheep and cattle. J. Anim. Sci. 1995, 73, 1509–1515. [Google Scholar] [CrossRef]
- Nichols, P.G.H. An agro-ecological approach to breeding subterranean clover. In Proceedings of the XVII International Grassland Congress, Hamilton, New Zealand, 8–21 February 1993; Baker, M.J., Crush, J.R., Humphries, L.R., Eds.; New Zealand Grassland Association: Palmerston North, New Zealand, 1993; Volume 1, pp. 453–454. [Google Scholar]
- Pace, V.; Contò, G.; Carfì, F.; Chiariotti, A.; Catillo, G. Short-and long-term effects of low estrogenic subterranean clover on ewe reproductive performance. Small Rumin. Res. 2011, 97, 94–100. [Google Scholar] [CrossRef]
- Choi, B.H.; Ohashi, H. Generic criteria and infrageneric system for Hedysarum and related genera (Papilionoideae-Leguminosae). Taxon 2003, 52, 567–576. [Google Scholar] [CrossRef]
- Borreani, G.; Roggero, P.P.; Sulas, L.; Valente, M.E. Quantifying morphological stage to predict the nutritive value in sulla (Hedysarum coronarium L.). Agron. J. 2003, 95, 1608–1617. [Google Scholar] [CrossRef]
- Sulas, L.; Seddaiu, G.; Muresu, R.; Roggero, P.P. Nitrogen fixation of sulla under Mediterranean conditions. Agron. J. 2009, 101, 1470–1478. [Google Scholar] [CrossRef]
- Sulas, L.; Piluzza, G.; Salis, M.; Deligios, P.A.; Ledda, L.; Canu, S. Cropping systems sustainability: Inoculation and fertilisation effect on sulla performances in a new cultivation area. Ital. J. Agron. 2017, 12, 315–324. [Google Scholar] [CrossRef] [Green Version]
- Amato, G.; Giambalvo, D.; Frenda, A.S.; Mazza, F.; Ruisi, P.; Saia, S.; Di Miceli, G. Sulla (Hedysarum coronarium L.) as potential feedstock for biofuel and protein. Bioenerg. Res. 2016, 9, 711–719. [Google Scholar] [CrossRef]
- Ruisi, P.; Siragusa, M.; Di Giorgio, G.; Graziano, D.; Amato, G.; Carimi, F.; Giambalvo, D. Pheno-morphological, agronomic and genetic diversity among natural populations of sulla (Hedysarum coronarium L.) collected in Sicily, Italy. Genet. Resour. Crop Evol. 2011, 58, 245–257. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Ruisi, P.; Di Miceli, G.; Pecetti, L. Morpho-physiological and adaptive variation of Italian germplasm of sulla (Hedysarum coronarium L.). Crop Pasture Sci. 2014, 65, 206–213. [Google Scholar] [CrossRef] [Green Version]
- Bonanno, A.; Di Miceli, G.; Di Grigoli, A.; Frenda, A.S.; Tornambè, G.; Giambalvo, D.; Amato, G. Effects of feeding green forage of sulla (Hedysarum coronarium L.) on lamb growth and carcass and meat quality. Animal 2011, 5, 148–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piluzza, G.; Sulas, L.; Bullitta, S. Tannins in forage plants and their role in animal husbandry and environmental sustainability: A review. Grass Forage Sci. 2014, 69, 32–48. [Google Scholar] [CrossRef]
- Yu, M.; Gouvinhas, I.; Rocha, J.; Barros, A.I. Phytochemical and antioxidant analysis of medicinal and food plants towards bioactive food and pharmaceutical resources. Sci. Rep. 2021, 11, 1–14. [Google Scholar]
- Cheynier, V.; Tomas-Barberan, F.A.; Yoshida, K. Polyphenols: From plants to a variety of food and nonfood uses. J. Agri. Food Chem. 2015, 63, 7589–7594. [Google Scholar] [CrossRef]
- Re, G.A.; Piluzza, G.; Sanna, F.; Molinu, M.G.; Sulas, L. Polyphenolic composition and antioxidant capacity of legume-based swards are affected by light intensity in a Mediterranean agroforestry system. J. Sci. Food Agric. 2019, 99, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Tava, A.; Pecio, Ł.; Lo Scalzo, R.; Stochmal, A.; Pecetti, L. Phenolic content and antioxidant activity in Trifolium germplasm from different environments. Molecules 2019, 24, 298. [Google Scholar] [CrossRef] [Green Version]
- Burlando, B.; Pastorino, G.; Salis, A.; Damonte, G.; Clericuzio, M.; Cornara, L. The bioactivity of Hedysarum coronarium extracts on skin enzymes and cells correlates with phenolic content. Pharm. Biol. 2017, 55, 1984–1991. [Google Scholar] [CrossRef] [Green Version]
- Molle, G.; Decandia, M.; Giovanetti, V.; Cabiddu, A.; Fois, N.; Sitzia, M. Responses to condensed tannins of flowering sulla (Hedysarum coronarium L.) grazed by dairy sheep: Part 1: Effects on feeding behaviour, intake, diet digestibility and performance. Livest. Sci. 2009, 123, 138–146. [Google Scholar] [CrossRef]
- Tibe, O.; Maegher, L.P.; Fraser, K.; Harding, D.R.K. Condensed tannins and flavonoids from the forage legume sulla (Hedysarum coronarium). J. Agric. Food Chem. 2011, 59, 9402–9409. [Google Scholar] [CrossRef] [PubMed]
- Šamec, D.; Karalija, E.; Šola, I.; Vujčić Bok, V.; Salopek-Sondi, B. The role of polyphenols in abiotic stress response: The influence of molecular structure. Plants 2021, 10, 118. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tava, A.; Biazzi, E.; Ronga, D.; Pecetti, L.; Avato, P. Biologically active compounds from forage plants. Phytochem. Rev. 2022, 21, 471–501. [Google Scholar] [CrossRef]
- Wu, F.; Shi, S.; Liu, C.; Zhang, H.; Li, X.; Li, Z. Optimization of the Extraction of Total Phenols from Medicago sativa and its Antioxidant Capacity. Chem. Biodivers. 2022, 19, e202100898. [Google Scholar] [CrossRef]
- Kabtni, S.; Sdouga, D.; Bettaib Rebey, I.; Save, M.; Trifi-Farah, N.; Fauconnier, M.L.; Marghali, S. Influence of climate variation on phenolic composition and antioxidant capacity of Medicago minima populations. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Uyar, Z.; Koz, Ö.; Uyar, E.; Arslan, Ü.; Koyuncu, I.; Nalbantsoy, A. Total phenolic, flavonoid, fatty acid contents and cytotoxic, antioxidant, and antimicrobial activities of Hedysarum aucheri. J. Pharm. Res. Int. 2017, 19, 1–13. [Google Scholar] [CrossRef]
- Bektaş, E.; Kaltalioğlu, K.; Şahin, H.; Türkmen, Z.; Kandemir, A. Analysis of phenolic compounds, antioxidant and antimicrobial properties of some endemic medicinal plants. Int. J. Second. Metab. 2018, 5, 75–86. [Google Scholar] [CrossRef]
- Re, G.A.; Piluzza, G.; Sulas, L.; Franca, A.; Porqueddu, C.; Sanna, F.; Bullitta, S. Condensed tannin accumulation and nitrogen fixation potential of Onobrychis viciifolia Scop. grown in a Mediterranean environment. J. Sci. Food Agric. 2014, 94, 639–645. [Google Scholar] [CrossRef]
- Ventura, M.R.; Bastianelli, D.; Deniz, S.; Saavedra, P.; Rey, L.; Bonnal, L.; Gonzalez-Garcia, E. Phenolic and tannin compounds in subtropical shrubs (Bituminaria bituminosa, Chamaecytisus proliferus, and Adenocarpus foliosus) and the effects on in vitro digestibility. Trop. Anim. Health Prod. 2019, 51, 1757–1761. [Google Scholar] [CrossRef] [PubMed]
- Sulas, L.; Canu, S.; Carroni, A.M.; Re, G.A.; Salis, M.; Piluzza, G. Chemical composition and antioxidant capacity in edible biomass of tagasaste (Chamaecytisus proliferus var. palmensis). Afr. J. Agric. Res. 2016, 11, 4277–4285. [Google Scholar]
- Rice-Evans, C.; Miller, N.; Paganga, G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Hu, W.; Guan, Y.; Feng, K. Biosynthesis of phenolic compounds and antioxidant activity in fresh-cut fruits and vegetables. Front. Microbiol. 2022, 13, 1782. [Google Scholar] [CrossRef]
- Sulas, L.; Re, G.A.; Bullitta, S.; Piluzza, G. Chemical and productive properties of two Sardinian milk thistle (Silybum marianum L. Gaertn.) populations as sources of nutrients and antioxidants: Genet. Resour. Crop Evol. 2016, 63, 315–326. [Google Scholar]
- Dobrinas, S.; Soceanu, A.; Popescu, V.; Carazeanu Popovici, I.; Jitariu, D. Relationship between Total Phenolic Content, Antioxidant Capacity, Fe and Cu Content from Tea Plant Samples at Different Brewing Times. Process 2021, 9, 1311. [Google Scholar] [CrossRef]
- Vergun, O.; Shymanska, O.; Rakhmetov, D.; Grygorieva, O.; Ivanišová, E.; Brindza, J. Parameters of antioxidant activity of Galega officinalis L. and Galega orientalis Lam. (Fabaceae Lindl.) plant raw material. Slovak J. Food Sci. 2020, 14, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Tava, A.; Stochmal, A.; Pecetti, L. Isoflavone content in subterranean clover germplasm from Sardinia. Chem. Biodivers. 2016, 13, 1038–1045. [Google Scholar] [CrossRef] [PubMed]
- Tava, A.; Biazzi, E.; Ronga, D.; Mella, M.; Doria, F.; D’Addabbo, T.; Candido, V.; Avato, P. Chemical identification of specialized metabolites from sulla (Hedysarum coronarium L.) collected in southern Italy. Molecules 2021, 26, 4606. [Google Scholar] [CrossRef]
- Pace, V.; Carbone, K.; Spirito, F.; Terzano, G.M.; Verna, M.; Settineri, D. Phytoestrogen content of subterranean clover related to ewe growth and reproduction. J. Anim. Feed Sci. 2004, 13, 547–550. [Google Scholar] [CrossRef]
- Enkhbat, G.; Nichols, P.G.; Foster, K.J.; Ryan, M.H.; Inukai, Y.; Erskine, W.; Reed, K. Diversity for morphological traits, flowering time and leaf isoflavone content among ecotypes of Trifolium subterraneum L. subsp. yanninicum and their relationships with site of origin. Crop Pasture Sci. 2021, 72, 1022–1033. [Google Scholar]
- Ates, E.; Tenikecier, H.S. Content of isoflavones and macroelements in five clover species (Trifolium spp.) grown on pasture: The influence of a slope aspect. J. Elem. 2022, 27, 379–392. [Google Scholar] [CrossRef]
- Riaz, U.; Kharal, M.A.; Murtaza, G.; Zaman, Q.; Javaid, S.; Malik, H.A.; Abbas, Z. Prospective roles and mechanisms of caffeic acid in counter plant stress: A mini review. Pak. J. Agric. Res. 2019, 32, 8. [Google Scholar] [CrossRef]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; XiaoHui, Z. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Chun, O.K.; Kim, D.O.; Moon, H.Y.; Kang, H.G.; Lee, C.Y. Contribution of individual polyphenolics to total antioxidant capacity of plums. J. Agric. Food Chem. 2003, 51, 7240–7245. [Google Scholar] [CrossRef] [PubMed]
- Hoang, N.T.; Golding, J.B.; Wilkes, M.A. The effect of postharvest 1-MCP treatment and storage atmosphere on ‘Cripps Pink’apple phenolics and antioxidant activity. Food Chem. 2011, 127, 1249–1256. [Google Scholar] [CrossRef] [PubMed]
- Molinu, M.G.; Piluzza, G.; Campesi, G.; Sulas, L.; Re, G.A. Antioxidant sources from leaves of Russian dandelion. Chem. Biodivers. 2019, 16, e1900250. [Google Scholar] [CrossRef]
- Sanna, F.; Piluzza, G.; Campesi, G.; Molinu, M.G.; Re, G.A.; Sulas, L. Antioxidant Contents in a Mediterranean Population of Plantago lanceolata L. Exploited for Quarry Reclamation Interventions. Plants 2022, 11, 791. [Google Scholar] [CrossRef]
- StatPoint Technologies Inc. Statgraphics Centurion XVI (2009) User Manual; StatPoint Technologies Inc.: Warrenton, VA, USA, 2009. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Phenological Stage | TEAC (mmol 100 g−1 DM) | TotP (g GAE kg−1 DM) | NTP (g GAE kg−1 DM) | TP (g GAE kg−1 DM) | TotF (g CE kg−1 DM) | |
|---|---|---|---|---|---|---|
| ABTS | DPPH | |||||
| Vegetative | 11.6 b | 10.6 a | 122.5 a | 64.2 a | 58.3 a | 10.1 a |
| Flower bud | 13.6 a | 9.2 b | 108.5 b | 57.7 a | 50.8 a | 9.5 b |
| Flowering | 7.4 c | 5.2 c | 61.3 c | 34.1 b | 27.2 b | 5.2 c |
| Seed ripening | 6.2 d | 4.4 d | 47.3 d | 23.9 c | 23.4 b | 3.4 d |
| Phenological Stage | TEAC (mmol 100 g−1 DM) | TotP (g GAE kg−1 DM) | NTP (g GAE kg−1 DM) | TP (g GAE kg−1 DM) | TotF (g CE kg−1 DM) | CT (g DE kg−1 DM) | |
|---|---|---|---|---|---|---|---|
| ABTS | DPPH | ||||||
| Vegetative | 20.3 a | 20.4 a | 92.7 a | 28.4 a | 64.3 a | 11.4 a | 20.3 b |
| Flower bud | 18.4 b | 18.0 b | 81.3 b | 22.6 b | 58.7 a | 10.0 b | 25.1 a |
| Flowering | 14.4 c | 14.6 c | 71.6 c | 22.3 b | 49.2 b | 9.1 c | 16.6 c |
| Seed ripening | 10.1 d | 10.0 d | 54.3 d | 20.2 c | 34.1 c | 5.3 d | 9.4 d |
| TotP | NTP | TP | TotF | References | ||
|---|---|---|---|---|---|---|
| Scientific Name | Common Name | |||||
| Trifolium subterraneum | Subterranean clover | 47–122 | 24–64 | 23–58 | 3–10 | Authors |
| Sulla coronaria | Sulla | 54–93 | 20–28 | 34–64 | 5–11 | Authors |
| Trifolium spumosum | Bladder clover | 32–36 | 20–32 | 15–16 | 2–3 | [34] |
| Trifolium alessandrinum | Berseem clover | 27–62 | n.a. | n.a. | n.a. | [41] |
| Trifolium pratense | Red clover | 8–73 | n.a. | n.a. | n.a. | [41] |
| Trifolium repens | White clover | 8–16 | n.a. | n.a. | n.a. | [41] |
| Medicago sativa | Lucerne | 12–16 | n.a. | n.a. | n.a. | [42] |
| Medicago minima | Bur medick | 17 | n.a. | n.a. | 10 | [43] |
| Hedysarum aucheri | Unknown | 16–122 | n.a. | n.a. | 9–66 | [44] |
| Hedysarum cappadocicum | Tatli tirfil | 51 | n.a. | n.a. | n.a. | [45] |
| Onobrychis nitida | Firat korungasi | 102 | n.a. | n.a. | n.a. | [45] |
| Onobrychis viciifolia | Sainfoin | 40–57 | 15–17 | n.a. | n.a. | [46] |
| Bituminaria bituminosa | Tedera | 13 | n.a. | n.a. | n.a. | [47] |
| Chamaecytisus proliferus | Tagasaste | 33–112 | 40–96 | 3–20 | 20–68 | [47,48] |
| Subterranean Clover | |||
|---|---|---|---|
| ABTS | DPPH | TotP | |
| DPPH | 0.7866 *** | ||
| TotP | 0.8226 *** | 0.9470 *** | |
| NTP | 0.7656 *** | 0.9242 ** | 0.9433 *** |
| TP | 0.7786 *** | 0.8506 *** | 0.9328 *** |
| TotF | 0.8744 *** | 0.9520 *** | 0.9800 *** |
| Sulla | |||
|---|---|---|---|
| ABTS | DPPH | TotP | |
| DPPH | 0.9448 *** | ||
| TotP | 0.9372 *** | 0.9326 *** | |
| NTP | 0.6776 ** | 0.7343 ** | 0.7496 ** |
| TP | 0.9282 *** | 0.9053 *** | 0.9821 *** |
| TotF | 0.9116 *** | 0.9110 *** | 0.9466 *** |
| CT | 0.7630 ** | 0.6660 ** | 0.6398 * |
| Phenolic Compounds | *tR (min) | **λmax (nm) | Vegetative | Flower Bud | Flowering | Seed Ripening |
|---|---|---|---|---|---|---|
| Chlorogenic acid | 10.69 | 330 | nd | nd | 1.22 | nd |
| Cryptochlorogenic acid | 11.27 | 330 | nd | nd | tr | nd |
| Hydroxycinnamic acid derivative | 18.10 | 330 | 3.70 b | 4.90 a | 0.45 c | 0.44 c |
| Quercetin 3-glucoside | 21.90 | 350 | 3.49 a | 2.78 b | 1.66 c | 0.87 d |
| Genistin | 22.65 | 280 | 7.66 b | 9.38 a | 4.46 c | 2.13 d |
| Isoflavone I | 28.04 | 280 | 17.38 a | 18.27 a | 7.22 b | 3.60 c |
| Luteolin | 34.05 | 350 | 0.43 a | 0.27 b | tr | nd |
| Sissotrin | 35.16 | 280 | 1.01 a | 0.97 a | 0.97 a | 0.45 b |
| Isoflavone II | 37.36 | 280 | 15.29 a | 12.62 b | 6.37 c | 3.19 d |
| Formononetin | 38.60 | 280 | nd | nd | nd | nd |
| Biochanin A | 41.70 | 280 | 0.85 a | 0.72 b | 0.55 c | 0.41 d |
| Sum | 49.81 | 49.91 | 22.90 | 11.09 |
| Phenolic Compounds | *tR (min) | **λmax (nm) | Vegetative | Flower Bud | Flowering | Seed Ripening |
|---|---|---|---|---|---|---|
| Neochlorogenic acid | 9.45 | 330 | tr | tr | tr | 0.10 |
| Chlorogenic acid | 10.69 | 330 | 3.36 a | 3.43 a | 1.91 b | 0.76 c |
| Cryptochlorogenic acid | 11.27 | 330 | 0.56 a | 0.28 b | 0.25 b | 0.25 b |
| Caffeic acid | 14.33 | 330 | 0.10 b | 0.09 b | 0.03 c | 0.30 a |
| Hydroxycinnamic acid derivative | 15.90 | 330 | 2.47 a | 0.33 c | 0.84 b | 0.35 c |
| Rutin | 19.91 | 350 | 0.38 a | 0.17 b | 0.18 b | 0.05 c |
| Quercetin 3-galattoside | 20.10 | 350 | 1.00 a | 0.67 b | 0.67 b | 0.19 c |
| Quercetin 3-glucoside | 21.00 | 350 | 0.11 a | 0.12 a | 0.12 a | 0.11 a |
| Isorhamnetin 3-rutinoside | 23.19 | 350 | 0.64 b | 1.23 a | 0.43 c | 0.27 d |
| Isorhamnetin 3-rutinoside (isomer) | 23.52 | 350 | 0.17 b | 0.29 a | 0.09 c | 0.03 d |
| Diosmin | 24.06 | 350 | 4.94 a | 3.76 bc | 3.93 b | 3.64 c |
| Sum | 13.73 | 10.37 | 8.45 | 6.05 |
| ABTS | DPPH | TotP | NTP | TP | TotF | |
|---|---|---|---|---|---|---|
| Hydroxycinnamic acid derivative | 0.9700 *** | 0.8071 *** | 0.8293 *** | 0.7614 ** | 0.7966 *** | 0.8624 *** |
| Quercetin 3-glucoside | 0.7486 ** | 0.9136 *** | 0.9612 *** | 0.9028 *** | 0.9007 *** | 0.9546 *** |
| Genistin | 0.9653 *** | 0.7808 *** | 0.8367 *** | 0.8120 *** | 0.7571 ** | 0.9003 *** |
| Isoflavone I | 0.9459 *** | 0.8929 *** | 0.9479 *** | 0.8924 *** | 0.8861 *** | 0.9749 *** |
| Luteolin | 0.7154 ** | 0.9436 *** | 0.9462 *** | 0.8719 *** | 0.9052 *** | 0.9002 *** |
| Sissotrin | 0.4400 * | 0.4232 * | 0.4654 * | 0.4621 * | 0.4098 * | 0.5211 * |
| Isoflavone II | 0.8024 *** | 0.9271 *** | 0.9815 *** | 0.9119 *** | 0.9306 *** | 0.9712 *** |
| Biochanin A | 0.7356 ** | 0.9156 *** | 0.9543 *** | 0.9021 *** | 0.8879 *** | 0.9502 *** |
| ABTS | DPPH | TotP | NTP | TP | TotF | CT | |
|---|---|---|---|---|---|---|---|
| Chlorogenic acid | 0.9489 *** | 0.9136 *** | 0.8544 *** | 0.5105 * | 0.8817 *** | 0.8404 *** | 0.8697 *** |
| Cryptochlorogenic acid | 0.4997 * | 0.5316 * | 0.5605 * | 0.9027 *** | 0.4371 * | 0.4305 * | 0.0920 ns |
| Caffeic acid | 0.4200 * | 0.4373 * | 0.4486 * | 0.1865 ns | 0.4947 * | 0.6311 * | 0.4716 * |
| Hydroxycinnamic acid derivative | 0.4023 * | 0.4400 * | 0.5082 * | 0.8649 *** | 0.3879 * | 0.4272 * | 0.0450 ns |
| Rutin | 0.7321 ** | 0.7754 ** | 0.8148 *** | 0.9413 *** | 0.7125 ** | 0.7677 ** | 0.2869 ns |
| Quercetin 3-galattoside | 0.8459 *** | 0.8600 *** | 0.8988 *** | 0.7645 *** | 0.8561 *** | 0.9177 *** | 0.5207 * |
| Quercetin 3-glucoside | 0.0031 ns | 0.0022 ns | 0.0020 ns | 0.0882 ns | 0.0180 ns | 0.0059 ns | 0.1348 ns |
| Isorhamnetin 3-rutinoside | 0.4669 * | 0.4186 * | 0.3276 * | 0.0495 ns | 0.4099 * | 0.3341 * | 0.8001 *** |
| Isorhamnetin 3-rutinoside isomer | 0.6073 * | 0.5625 * | 0.4746 * | 0.1311 ns | 0.5557 * | 0.4845 * | 0.8675 *** |
| Diosmin | 0.4726 * | 0.5165 * | 0.5698 * | 0.8652 *** | 0.4513 * | 0.5052 * | 0.0990 ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molinu, M.G.; Sulas, L.; Campesi, G.; Re, G.A.; Sanna, F.; Piluzza, G. Subterranean Clover and Sulla as Valuable and Complementary Sources of Bioactive Compounds for Rainfed Mediterranean Farming Systems. Plants 2023, 12, 417. https://doi.org/10.3390/plants12020417
Molinu MG, Sulas L, Campesi G, Re GA, Sanna F, Piluzza G. Subterranean Clover and Sulla as Valuable and Complementary Sources of Bioactive Compounds for Rainfed Mediterranean Farming Systems. Plants. 2023; 12(2):417. https://doi.org/10.3390/plants12020417
Chicago/Turabian StyleMolinu, Maria Giovanna, Leonardo Sulas, Giuseppe Campesi, Giovanni Antonio Re, Federico Sanna, and Giovanna Piluzza. 2023. "Subterranean Clover and Sulla as Valuable and Complementary Sources of Bioactive Compounds for Rainfed Mediterranean Farming Systems" Plants 12, no. 2: 417. https://doi.org/10.3390/plants12020417