
In Vitro α-Amylase and α-Glucosidase Inhibitory Activity of Green Seaweed Halimeda tuna Extract from the Coast of Lhok Bubon, Aceh
 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Inhibitory Activity against α-Amylase
2.2. Inhibitory Activity against α-Glucosidase
2.3. Phytochemical Test
2.4. Identification of Active Compounds Using GC-MS
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Identification
4.2. Extraction of Seaweed
4.3. Liquid–Liquid Partition
4.4. Inhibitory Activity against α-Amylase
4.5. Inhibitory Activity against α-Glucosidase
4.6. Phytochemical Analysis
4.6.1. Flavonoid Test
4.6.2. Steroid and Triterpenoid Test
4.6.3. Tannin Test
4.6.4. Alkaloid Test
4.6.5. Saponin Test
4.6.6. Phenol Hydroquinone Test
4.7. Identification of Active Compounds Using GC-MS
4.8. Data Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dipiro, J.T.; Talbert, R.L.; Yee, G.C. Pharmacotheraphy a Phtophysiologic Approach, 6th ed.; McGraw Hill: New York, NY, USA, 2005; pp. 1333–1367. [Google Scholar]
- Yuniastuti, A.; Susanti, R.; Iswari, R.S. Efek infusa umbi garut (Marantha arundinaceae L.) terhadap kadar glukosa dan insulin plasma tikus yang diinduksi streptozotocyn. Indones. J. Math. Nat. Sci. 2018, 41, 34–39. [Google Scholar] [CrossRef]
- Putri, R.E. Hubungan Pola Konsumsi Makanan Terhadap Terjadinya Diabetes Melitus Tipe 2 di UPTD Puskesmas Makmur Kabupaten Bireuen. Bachelor’s Thesis, Universitas Sumatera Utara, Medan, Indonesia, 2021. [Google Scholar]
- Souza, P.M.; Sales, P.M.; Simeoni, L.A.; Silva, E.C.; Silveira, D.; de Oliveira Magalhães, P. Inhibitory activity of α-amylase and α-glucosidase by plant extracts from the Brazilian cerrado. Planta Med. 2012, 78, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Wahyuntari, B. Penghambatan α-amilase: Jenis, sumber, dan potensi pemanfaatannya dalam kesehatan. J. Teknol. Dan Ind. Pangan 2011, 22, 197–201. [Google Scholar]
- Firdaus, M.; Astawan, M.; Muchtadi, D.; Wresdyani, T.; Waspadji, S.; Setyawati, S.K. Pengaruh ekstrak rumput laut cokelat terhadap fungsi endotelium aorta tikus diabetes melitus. Maj. Farm. Indones. 2010, 21, 151–157. [Google Scholar] [CrossRef]
- Sun, C.; Zhao, C.; Guven, E.C.; Paoli, P.; Simal-Gandara, J.; Ramkumar, K.M.; Wang, S.; Buleu, F.; Pah, A.; Turi, V.; et al. Dietary polyphenols as antidiabetic agents: Advances and opportunities. Food Front. 2020, 1, 18–44. [Google Scholar] [CrossRef] [Green Version]
- Milović, S.; Kundaković, T.; Mačić, V.E.S.N.A.; Stanković, J.A.; Grozdanić, N.A.Đ.A.; Đuričć, I.; Stanković, I.; Stanojković, T. Anti α-glucosidase, antitumour, antioxidative, antimicrobial activity, nutritive and health protective potential of some seaweeds from the Adriatic coast of Montenegro. Farmacia 2017, 65, 731–740. [Google Scholar]
- Abdelrheem, D.A.; Rahman, A.A.; Elsayed, K.N.M.; Ahmed, S.A. GC/MS spectroscopic approach, antimicrobial activity and cytotoxicity of some marine macroalgae from Qusier and Marsa Alam Seashore (Red Sea), Egypt. Egypt. J. Aquat. Biol. Fish. 2020, 24, 125–144. [Google Scholar] [CrossRef]
- Gazali, M.; Nurjanah; Zamani, N.P. The screening of bioactive compound of the green algae Halimeda macroloba (Decaisne, 1841) as an antioxidant agent from Banyak Island Aceh Singkil. IOP Conf. Ser. Earth Environ. Sci. 2019, 348, 012043. [Google Scholar] [CrossRef]
- Gazali, M.; Nurjanah; Zamani, N.P.; Zuriat; Nasution, M.A. A study on a potential bioactive compound in green seaweed Chaetomorpha antennina Kützing (1847) extract as antioxidant from the Gosong Telaga Coast, Aceh Singkil. IOP Conf. Ser. Earth Environ. Sci. 2020, 564, 012058. [Google Scholar] [CrossRef]
- Gazali, M.; Nurjanah; Zamani, N.P.; Nasution, M.A.; Zuriat; Syafitri, R. Screening for antioxidant activity in extracts of the marine macro algae Enteromorpha flexuosa (Wulfen) J. Agardh from South Aceh. IOP Conf. Ser. Earth Environ. Sci. 2021, 763, 012027. [Google Scholar] [CrossRef]
- Diansyah, S.; Kusumawati, I.; Hardinata, F. Inventarisasi jenis-jenis makroalga di Pantai Lhok Bubon Kecamatan Samatiga Kabupaten Aceh Barat. J. Perikan. Trop. 2018, 5, 93–103. [Google Scholar] [CrossRef]
- Gazali, M. Aktivitas inhibitor tirosinase rumput laut Halimeda spp. dari Pesisir Aceh Barat. J. Perikan. Trop. 2018, 5, 149–159. [Google Scholar] [CrossRef]
- Erniati, E.; Syahrial, S.; Imanullah, I.; Erlangga, E.; Nurul‘Akla, C.M.; Shobara, W.; Nasuha, J.; Ritonga, G.H.; Daulay, A.M.; Romansah, H.; et al. Rumput laut yang tumbuh alami di pantai Barat Pulau Simeulue, Aceh Indonesia: Faktor lingkungan dan variasi geografik. J. Perikan. Trop. 2022, 25, 29–38. [Google Scholar] [CrossRef]
- Gazali, M.; Nurjanah; Neviaty, P.Z. Skreening alga hijau Halimeda opuntia (Linnaeus) sebagai antioksidan dari Pesisir Aceh Barat. J. Ilmu Pertan. Indones. 2019, 24, 267–272. [Google Scholar] [CrossRef]
- Jhong, C.H.; Riyaphan, J.; Lin, S.H.; Chia, Y.C.; Weng, C.F. Screening alpha-glucosidase and alpha-amylase inhibitors from natural compounds by molecular docking in silico. Biofactors 2015, 41, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Saranya, M.S.; Arunprasath, A. Evaluation of phytochemical compounds in Corbichonia decumbens (Frossk). Excell by using Gas Chromatography-Mass Spectrometry. J. Appl. Adv. Res 2019, 4, 89–93. [Google Scholar] [CrossRef] [Green Version]
- Madhubala, M.; Santhi, G. Phytochemical and GC-MS analysis on leaves of selected medicinal plants in Boraginaceae family Cordia dichotoma L. Pramana Res. J. 2019, 9, 2249–2276. [Google Scholar]
- Kanthaiah, K.; Velu, R.K. Characterization of the bioactive metabolite from a plant growth promoting rhizobacteria Pseudomonas aeruginosa VRKK1 and exploitation of antibacterial behavior against Xanthomonas campestris a causative agent of bacterial blight disease in cowpea. Arch. Phytopathol. Plant Prot. 2022, 55, 797–814. [Google Scholar] [CrossRef]
- Wuttke, A.; Idevall-Hagren, O.; Tengholm, A. P2Y1 receptor-dependent diacylglycerol signaling microdomains in β cells promote insulin secretion. FASEB J. 2013, 27, 1610–1620. [Google Scholar] [CrossRef]
- Anjali, K.P.; Sangeetha, B.M.; Devi, G.; Raghunathan, R.; Dutta, S. Bioprospecting of seaweeds (Ulva lactuca and Stoechospermum marginatum): The compound characterization and functional applications in medicine-a comparative study. J. Photochem. Photobiol. B Biol. 2019, 200, 111622. [Google Scholar] [CrossRef]
- Conforti, F.; Loizzo, M.R.; Statti, G.A.; Menichini, F. Comparative radical scavenging and antidiabetic activities of methanolic extract and fractions from Achillea ligustica ALL. Biol. Pharm. Bull. 2005, 28, 1791–1794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, M.T.; Ali, E.S.; Uddin, S.J.; Shaw, S.; Islam, M.A.; Ahmed, M.I.; Shill, M.C.; Karmakar, U.K.; Yarla, N.S.; Khan, I.N.; et al. Phytol: A review of biomedical activities. Food Chem. Toxicol. 2018, 121, 82–94. [Google Scholar] [CrossRef]
- Elmazar, M.M.; El-Abhar, H.S.; Schaalan, M.F.; Farag, N.A. Phytol/phytanic acid and insulin resistance: Potential role of phytanic acid proven by docking simulation and modulation of biochemical alterations. PLoS ONE 2013, 8, e45638. [Google Scholar] [CrossRef]
- Amudha, M.; Rani, S. GC-MS analysis of bioactive components of Cordia retusa (Boraginaceae). Hygeia J. Drug Med. 2014, 6, 12–19. [Google Scholar] [CrossRef]
- Zayed, M.Z.; Wu, A.; Sallam, S.M. Comparative phytochemical constituents of Leucaena leucocephala (Lam.) leaves, fruits, stem barks, and wood branches grown in Egypt using GC-MS method coupled with multivariate statistical approaches. BioResources 2019, 14, 996–1013. [Google Scholar] [CrossRef]
- Al-Marzoqi, A.H.; Hameed, I.H.; Idan, S.A. Analysis of bioactive chemical components of two medicinal plants (Coriandrum sativum and Melia azedarach) leaves using gas chromatography-mass spectrometry (GC-MS). Afr. J. Biotechnol. 2015, 14, 2812–2830. [Google Scholar] [CrossRef] [Green Version]
- Signe, J.K.; Aponglen, G.A.; Ajeck, J.M.; Taiwe, G.S. Anticonvulsant activities of friedelan-3-one and n-dotriacontane both isolated from Harungana madagascariensis Lam (Hypericaceae) seeds extracts. J. Med. Plants Res. 2020, 14, 509–517. [Google Scholar]
- Soosairaj, S.; Dons, T. Bio-active compounds analysis and characterization in Ethanolic plant extracts of Justicia tranquebariensis L. (Acanthaceae)-using GC-MS. Int. J. Chemtech Res. 2016, 9, 260–265. [Google Scholar]
- Senarath, W.T.P.S.K.; Pizon, J.R.L.; Nuñeza, O.M.; Uy, M.M. In-vitro alpha-amylase inhibitory activity, antioxidant potential, and GC-MS analysis of crepe ginger (Costus speciosus (J. Koenig.) Sm) leaves. Int. J. Pharma. Sci. Res. 2018, 9, 4741–4749. [Google Scholar]
- Kemit, N.; Widarta, I.W.R.; Nocianitri, K.A. Pengaruh jenis pelarut dan waktu maserasi terhadap kandungan senyawa flavonoid dan aktivitas antioksidan ekstrak daun alpukat (Persea Americana Mill). ITEPA J. Ilmu Dan Teknol. Pangan 2016, 5, 130–141. [Google Scholar]
- Iwai, K. Antidiabetic and antioxidant effect of polyphenols in brown alga Ecklonia stolonifera in genetically diabetic kk-a9y mice. Plant Food Hum. Nutr. 2008, 63, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Basir, A.; Tarman, K.; Desniar, D. Antibacterial and antioxidant activity of green algae Halimeda gracilis from Seribu Island District. JPHPI 2017, 20, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Harbone, J.B. Phytochemical Methods—A Guide to Modern Techniques of Plant Analysis, 3rd ed.; Chapman and Hall: London, UK, 1998; pp. 36–89. [Google Scholar]
- Chin, Y.X.; Lim, P.E.; Maggs, C.A.; Phang, S.M.; Sharifuddin, Y.; Green, B.D. Anti-diabetic potential of selected Malaysian seaweeds. J. Appl. Phycol. 2015, 27, 2137–2148. [Google Scholar] [CrossRef]
- Pacheco, L.V.; Parada, J.; Pérez-Correa, J.R.; Mariotti-Celis, M.S.; Erpel, F.; Zambrano, A.; Palacios, M. Bioactive polyphenols from southern Chile seaweed as inhibitors of enzymes for starch digestion. Mar. Drugs 2020, 18, 353. [Google Scholar] [CrossRef]
- Mohapatra, L.; Bhattamisra, S.K.; Panigrahy, R.C.; Parida, S.K. Evaluation of the antioxidant, hypoglycemic and anti-diabetic activities of some seaweed collected from the East Coast of India. Biomed. Pharmacol. J. 2016, 9, 365–375. [Google Scholar] [CrossRef]
- Gunathilaka, T.L.; Samarakoon, K.W.; Ranasinghe, P.; Peiris, L.D.C. In-vitro antioxidant, hypoglycemic activity, and identification of bioactive compounds in phenol-rich extract from the marine red algae Gracilaria edulis (Gmelin) Silva. Molecules 2019, 24, 3708. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Kumari, S.N.; Bhargavan, D. Evaluation of in vitro antioxidant potential of ethanolic extract from the leaves of Achyranthes aspera. Asian J. Pharm. Clinic. Res. 2012, 5, 146–148. [Google Scholar]
- Reka, P.; Banu, T.; Seethalakshmi, M. Alpha amylase and alpha glucosidase inhibition activity of selected edible seaweeds from south coast area of India. Int. J. Pharm. Pharm. Sci. 2017, 9, 64–68. [Google Scholar] [CrossRef]
- Husni, A.; Pratiwi, T.; Samudra, A.G.; Nugroho, A.E. In vitro antidiabetic activity of Sargassum hystrix and Eucheuma denticulatum from Yogyakarta Beach of Indonesia: Antidiabetic activity of S. hystrix and E. denticulatum. Proc. Pak. Acad. Sci. B Life Environ. Sci. 2018, 55, 1–8. [Google Scholar]
- Nguyen, T.H.; Nguyen, T.L.P.; Tran, T.V.A.; Do, A.D.; Kim, S.M. Antidiabetic and antioxidant activities of red seaweed Laurencia dendroidea. Asian Pac. J. Trop. Biomed. 2019, 9, 501. [Google Scholar] [CrossRef]
- Pirian, K.; Moein, S.; Sohrabipour, J.; Rabiei, R.; Blomster, J. Antidiabetic and antioxidant activities of brown and red macroalgae from the Persian Gulf. J. App. Phycol. 2017, 29, 3151–3159. [Google Scholar] [CrossRef]
- Sanger, G.; Rarung, L.K.; Damongilala, L.J.; Kaseger, B.E.; Montolalu, L.A.D.Y. Phytochemical constituents and antidiabetic activity of edible marine red seaweed (Halymenia durvilae). IOP Conf. Ser. Earth Environ. Sci. 2019, 278, 012069. [Google Scholar] [CrossRef]
- Payghami, N.; Jamili, S.; Rustaiyan, A.; Saeidnia, S.; Nikan, M.; Gohari, A.R. Alpha-amylase inhibitory activity and sterol composition of the marine algae, Sargassum glaucescens. Pharmac. Res. 2015, 7, 314–321. [Google Scholar] [CrossRef] [Green Version]
- Senthilkumar, P.; Sudha, S. Evaluation of alpha-amylase and alphaglucosidase inhibitory properties of selected seaweeds from Gulf of Mannar. Int. Res. J. Pharm. 2012, 3, 128–130. [Google Scholar]
- Surya, S.; Salam, A.D.; Tomy, D.V.; Carla, B.; Kumar, R.A.; Sunil, C. Diabetes mellitus and medical plants-a review. Asian Pac. J. Trop. Dis. 2014, 4, 337–347. [Google Scholar] [CrossRef]
- Adhikari, B. Roles of alkaloids from medicinal plants in the management of diabetes mellitus. J. Chem. 2021, 2021, 2691525. [Google Scholar] [CrossRef]
- Gabay, O.; Sanchez, C.; Salvat, C.; Chevy, F.; Breton, M.; Nourissat, G.; Wolf, C.; Jacques, C.; Berenbaum, F. Stigmasterol: A phytosterol with potential anti-osteoarthritic properties. Osteoarthr. Cartil. 2010, 18, 106–116. [Google Scholar] [CrossRef]
- Lauro, F.V.; Francisco, D.C.; Lenin, H.H.; Elodia, G.C.; Eduardo, P.G.; Marcela, R.N.; Bety, S.A. New steroid derivative with hypoglycemic activity. Int. J. Clin. Exp. Med. 2014, 7, 3983–3991. [Google Scholar]
- Jacques, P.F.; Cassidy, A.; Rogers, G.; Peterson, J.J.; Meigs, J.B.; Dwyer, J.T. Higher dietary flavonol intake is associated with lower incidence of type 2 diabetes. J. Nutr. 2013, 143, 1474–1480. [Google Scholar] [CrossRef] [Green Version]
- Nicolle, E.; Souard, F.; Faure, P.; Boumendjel, A. Flavonoids as promising lead compounds in type 2 diabetes mellitus: Molecules of interest and structure-activity relationship. Curr. Med. Chem. 2011, 18, 2661–2672. [Google Scholar] [CrossRef]
- Oke, J.M.; Hamburger, M.O. Screening of Some Nigerian Medicinal Plants for Antioxidant Activity Using 2, 2, diphenyl-picryl-hydrazyl radical. Afr. J. Biomed. Res. 2002, 5, 77–79. [Google Scholar] [CrossRef]
- Xu, H. Inhibition kinetics of flavonoids on yeast α-glucosidase merged with docking simulation. Prot. Pept. Lett. 2010, 17, 1270–1279. [Google Scholar] [CrossRef]
- Ghani, U. Re-exploring promising α-glucosidase inhibitors for potential development into oral anti-diabetic drugs: Finding needle in the haystack. Europ. J. Med. Chem. 2015, 103, 133–162. [Google Scholar] [CrossRef] [PubMed]
- Su, C.H.; Hsu, C.H.; Ng, L.T. Inhibitory potential of fatty acids on key enzymes related to type 2 diabetes. Biofactors 2013, 39, 415–421. [Google Scholar] [CrossRef] [PubMed]
- El Barky, A.R.; Hussein, S.A.; Alm-Eldeen, A.A.; Hafez, Y.A.; Mohamed, T.M. Anti-diabetic activity of Holothuria thomasi saponin. Biomed. Pharmacother. 2016, 84, 1472–1487. [Google Scholar] [CrossRef] [PubMed]
- Janaki, M.; Santhi, V.; Kannagi, A.N.I.T.A. Bioactive potential of Fusinus nicobaricus from Gulf of Mannar. Int. J. Pharmaaceut. Res. Biosci. 2015, 4, 262–270. [Google Scholar] [CrossRef]
- Rocha, O.P.; Felício, R.D.; Rodrigues, A.H.B.; Ambrósio, D.L.; Cicarelli, R.M.B.; Albuquerque, S.D.; Young, M.C.M.; Yokoya, N.S.; Debonsi, H.M. Chemical profile and biological potential of non-polar fractions from Centroceras clavulatum (C. Agardh) Montagne (Ceramiales, Rhodophyta). Molecules 2011, 16, 7105–7114. [Google Scholar] [CrossRef]
- Song, Y.; Cho, S.K. Phytol induces apoptosis and ROS-mediated protective autophagy in human gastric adenocarcinoma AGS cells. Biochem. Anal. Biochem. 2015, 4, 1. [Google Scholar] [CrossRef]
- Poulose, N.; Sajayan, A.; Ravindran, A.; Chandran, A.; Priyadharshini, G.B.; Selvin, J.; Kiran, G.S. Anti-diabetic potential of a stigmasterol from the seaweed Gelidium spinosum and its application in the formulation of nanoemulsion conjugate for the development of functional biscuits. Front. Nut. 2021, 8, 694362. [Google Scholar] [CrossRef]
- Sundaram, S.M.; Bharathi, T.; Pennarasi, G.; Sabarirajan, P.; Vishalanand, M. Studies on phytochemicals, antibacterial efficacy and antioxidant potency of Capparis sepiaria on enteric pathogens. IJBB 2011, 1, 1–7. [Google Scholar] [CrossRef]
- Payum, T. Phytoconstituents and proximate composition of Clerodendrum colebrookianum walp.: A widely used anti high blood pressure medicinal food plant in Eastern Himalayas. Pharmacog. J. 2020, 12, 1534–1540. [Google Scholar] [CrossRef]
- Matsuda, H.; Daisuke, S.; Masaya, A.; Shinpei, O.; Saitoh, R.; Ryo, O.; Takaaki, S. Effects of Dietary Phytol on glucose uptake and insulin secretion in vitro and in vivo. Food Nutr. Curr. Res. 2018, 1, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Osman, S.M.; Hussein, M.A. Purslane seeds fixed oil as a functional food in treatment of obesity induced by high fat diet in obese diabetic mice. J. Nutr. Food Sci. 2015, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Azizi, W.A.; Ekantari, N.; Husni, A. Inhibitory activity of Sargassum hystrix extract and its methanolic fractions on inhibiting α-glucosidase activity. Indones. J. Pharm. 2019, 30, 35–42. [Google Scholar] [CrossRef]
- Devi, E.T. Isolasi dan identifikasi senyawa flavanoid pada ekstrak daun seledri (Apium graveolens L.) dengan metode refluks. PSEJ 2017, 2, 56–67. [Google Scholar] [CrossRef]
- Widowati, R.; Handayani, S.; Suprihatin, I.L.R. Phytochemicals and antioxidant of methanol extract of Gracilaria salicornia, Halimeda gracilis, Halimeda macroloba, and Hypnea asperi from Tidung island coastal region. Eur. J. Mol. Clin. Med. 2021, 8, 896–907. [Google Scholar]
- Hidayah, E.N. Analisis Metabolomik Padi Hitam (Oryza sativa L.) Setelah Infeksi Xanthomonas Oryzae pv.oryzae. Master’s Thesis, Universitas Gadjah Mada, Yogyakarta, Indonesia, 2017. [Google Scholar]
 ) and acarbose (
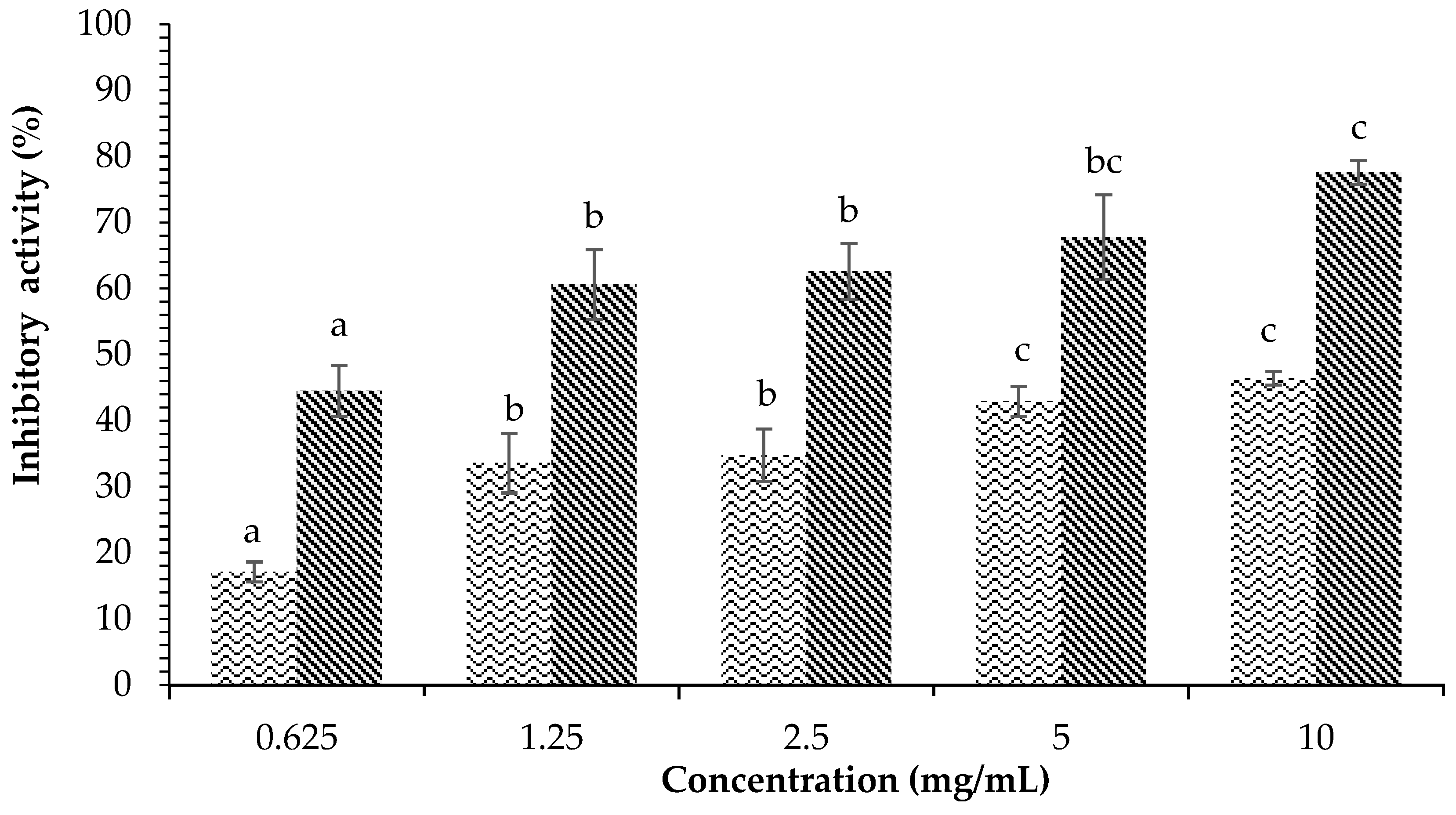
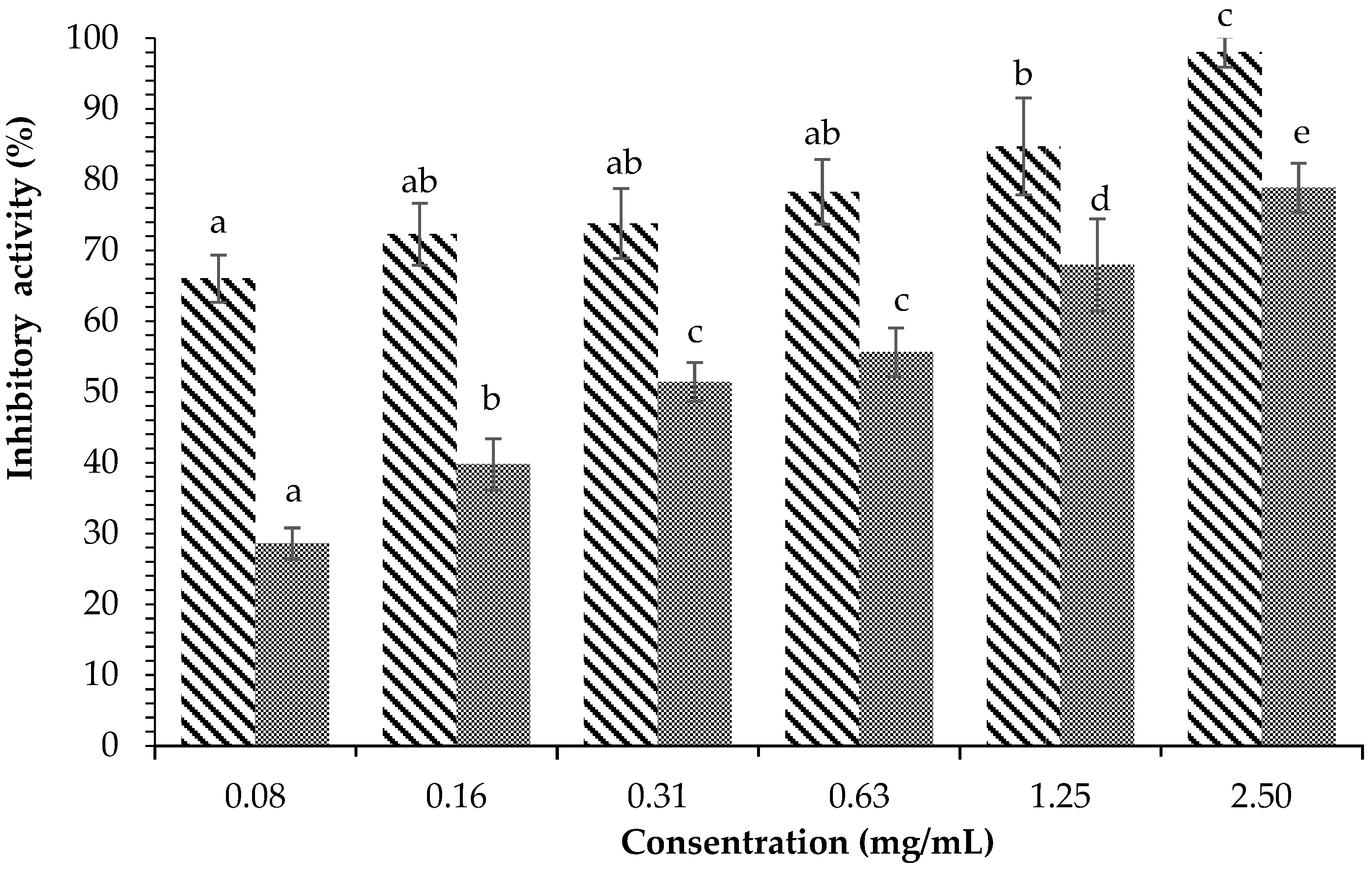
) and acarbose ( ) on α-amylase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–c) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
) and acarbose () on α-amylase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–c) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
) on α-amylase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–c) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
) and acarbose () on α-amylase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–c) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
 ) and acarbose (
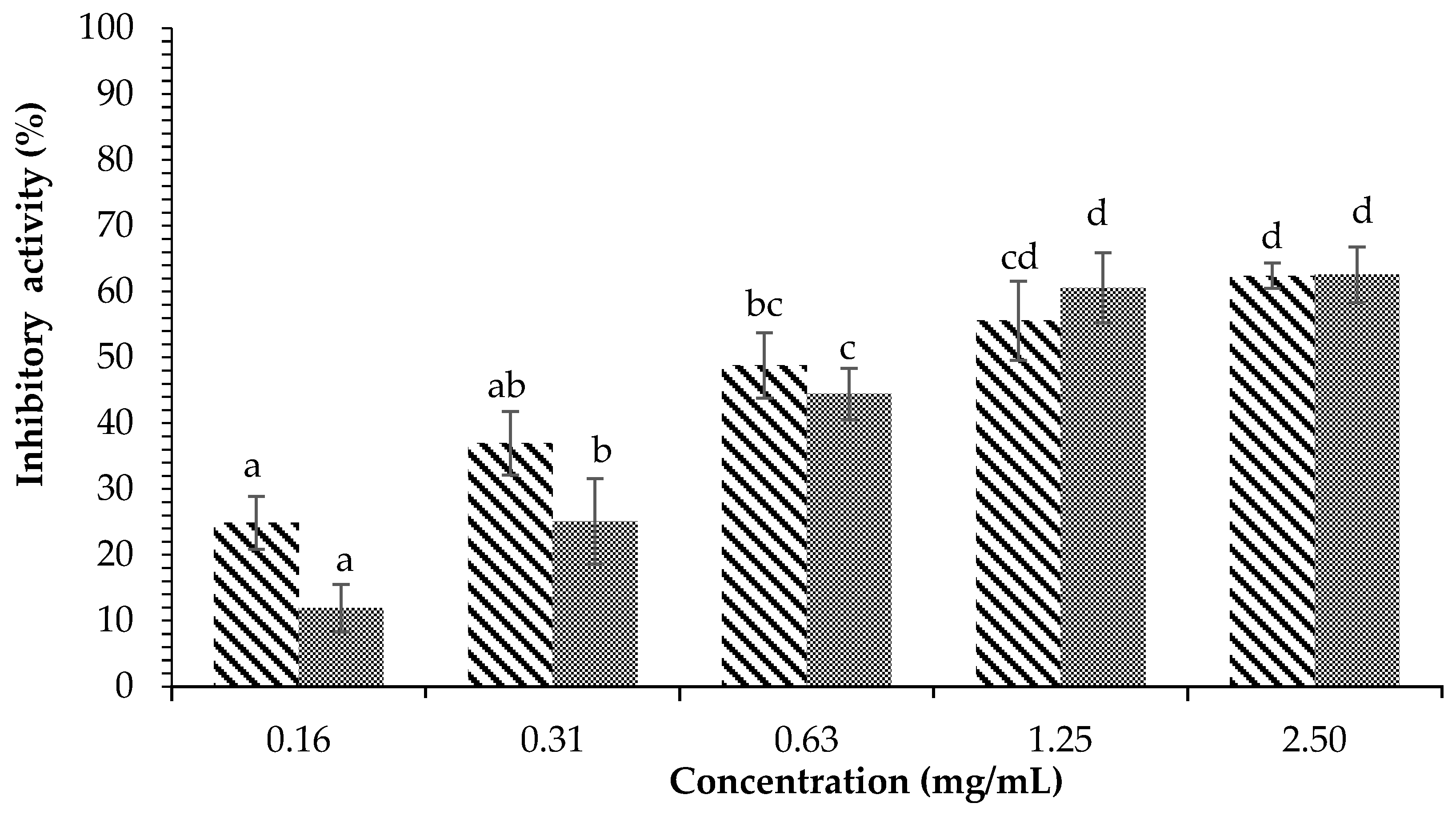
) and acarbose ( ) on α-amylase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–d) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
) and acarbose () on α-amylase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–d) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
) on α-amylase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–d) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
) and acarbose () on α-amylase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–d) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
 ) and Acarbose (
) and Acarbose ( ) on α-amylase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–d) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
) and Acarbose () on α-amylase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–d) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
) on α-amylase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–d) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
) and Acarbose () on α-amylase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–d) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
 ) and Acarbose (
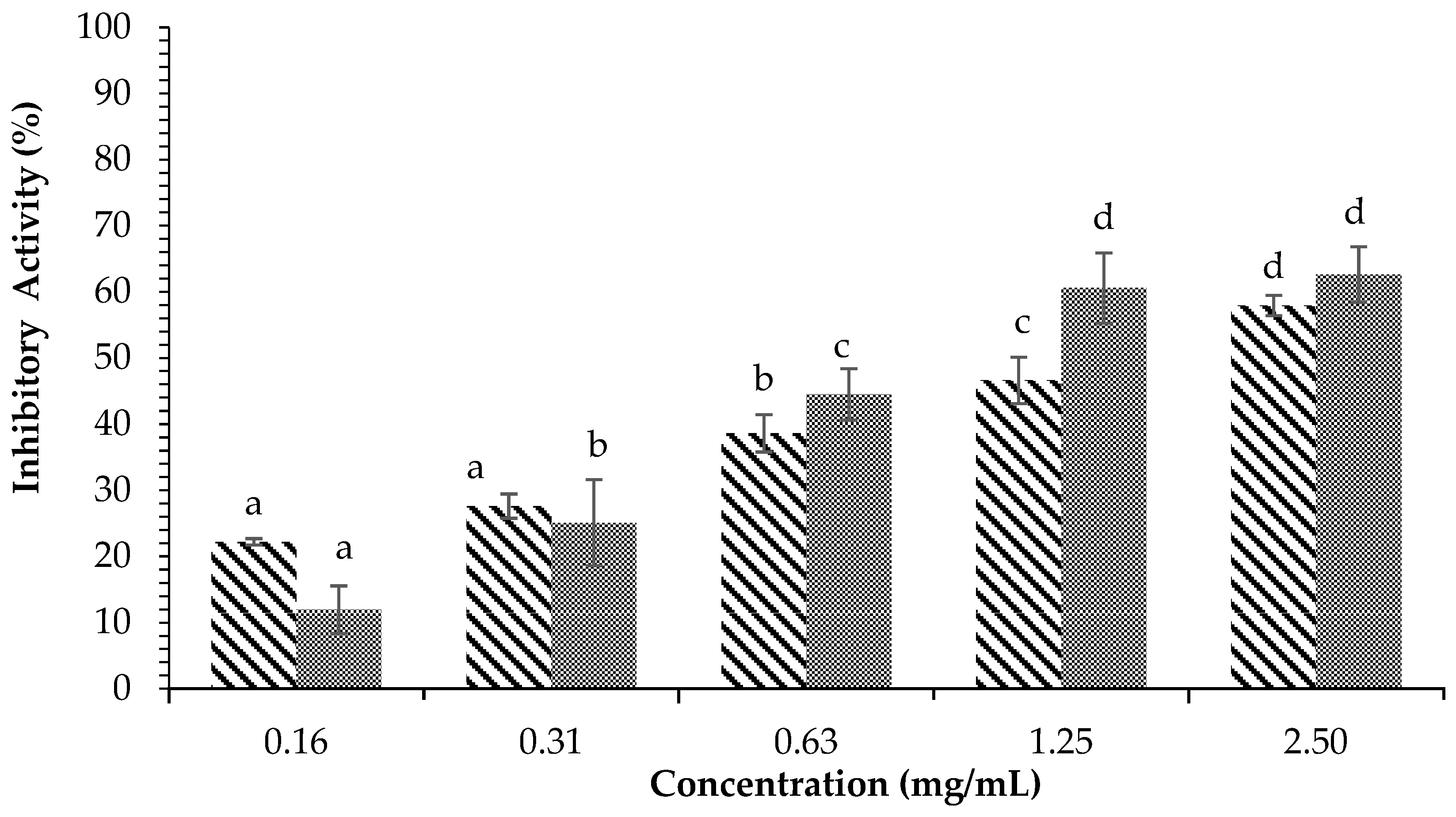
) and Acarbose ( ) on α-glucosidase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–d) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
) and Acarbose () on α-glucosidase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–d) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
) on α-glucosidase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–d) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
) and Acarbose () on α-glucosidase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–d) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
 ) and Acarbose (
) and Acarbose ( ) on α-glucosidase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–e) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
) and Acarbose () on α-glucosidase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–e) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
) on α-glucosidase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–e) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
) and Acarbose () on α-glucosidase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–e) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
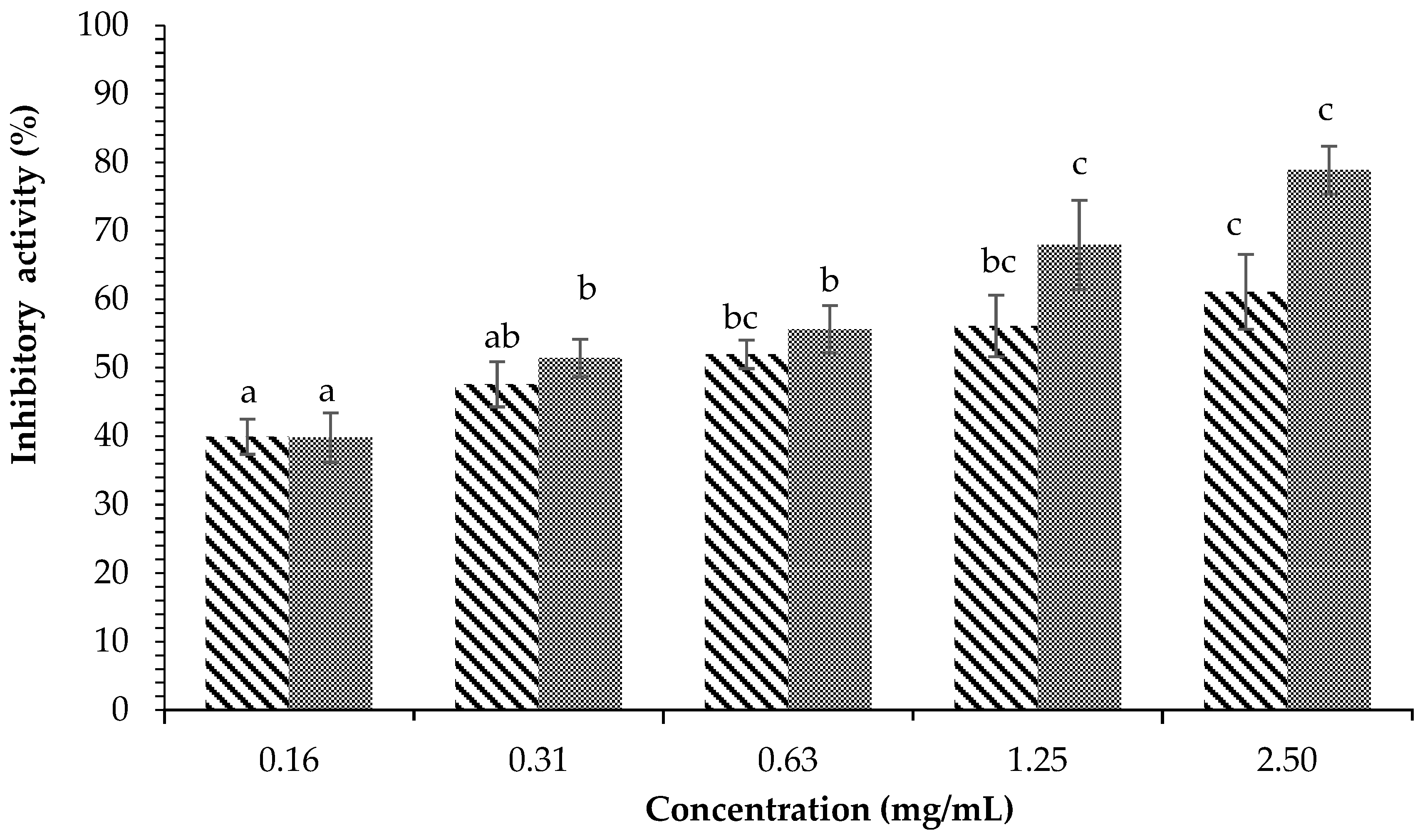
 ) and Acarbose (
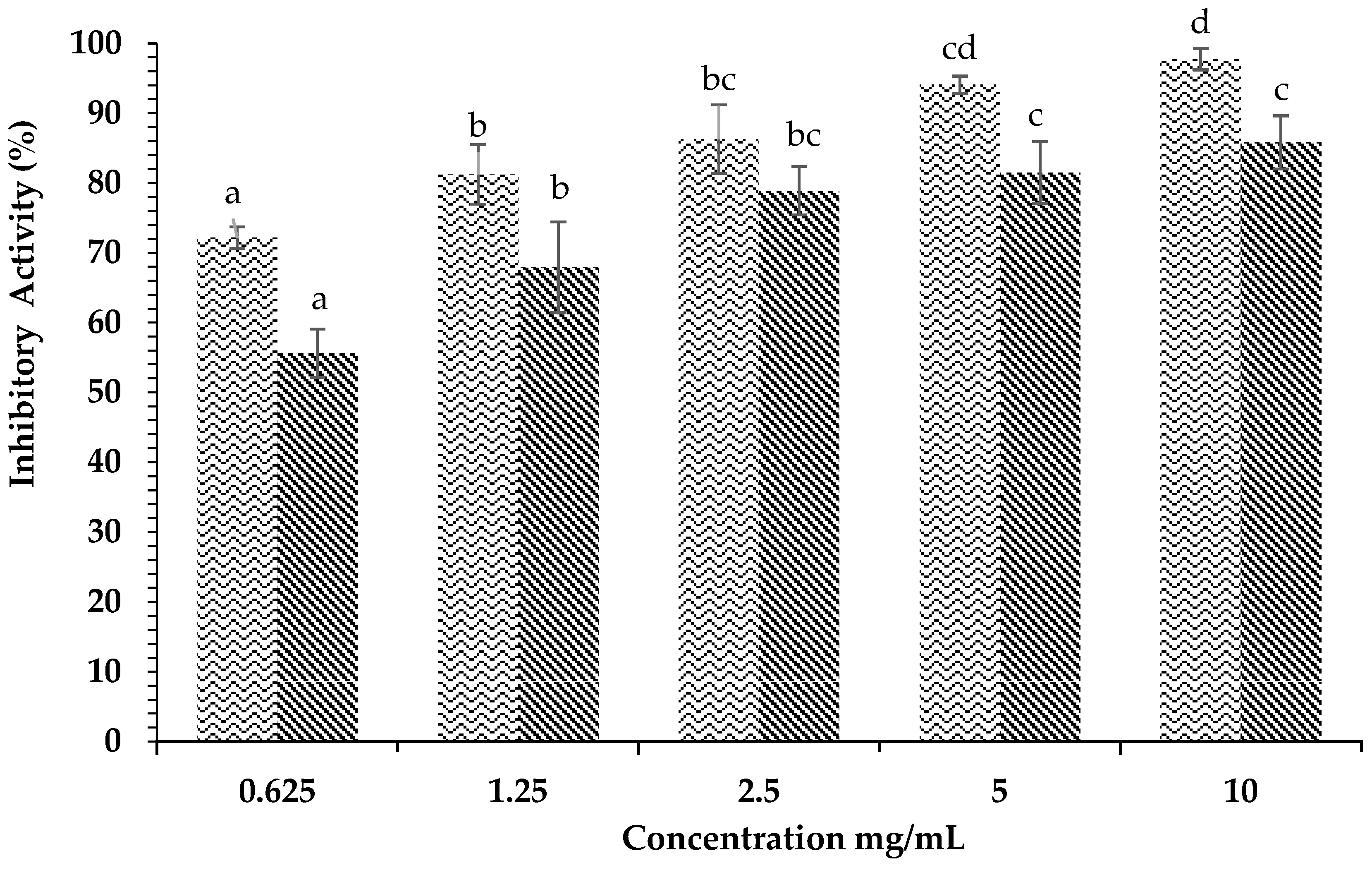
) and Acarbose ( ) on α-glucosidase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–c) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
) and Acarbose () on α-glucosidase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–c) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
) on α-glucosidase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–c) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.
) and Acarbose () on α-glucosidase inhibition. Each value is expressed as mean ± SD in the triplicate experiment. Values (a–c) with different alphabet letters indicate significant differences among treatments at p < 0.05, which was analyzed using Tukey’s HSD.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inhibitor | α-Amylase (mg/mL) | α-Glucosidase (mg/mL) |
|---|---|---|
| Methanol extract | 11.58 ± 0.38 a | 0.05 ± 0.01 ab |
| Water fraction | 1.50 ± 0.13 b | 0.55 ± 0.12 c |
| Ethyl acetate fraction | 0.87 ± 0.20 c | 0.01 ± 0.00 a |
| Acarbose | 0.76 ± 0.04 c | 0.27 ± 0.13 b |
| Test | Methanol Extract | Ethyl Acetate Fraction | Standard (Color) |
|---|---|---|---|
| Alkaloids | |||
| Dragendorff | + | ++ | Red or orange precipitate |
| Mayer | - | - | Yellowish precipitate |
| Wagner | - | - | Brown precipitate |
| Steroids | + | ++ | Green/blue |
| Triterpenoids | - | - | Red/purple |
| Saponins | - | - | Stable foam forms |
| Flavonoids | ++ | + | Yellow-orange |
| Phenol hydroquinone | + | + | Green/blue to red |
| Tannins | - | - | Dark blue color |
| No. | RT | % Area | Compound | Activity |
|---|---|---|---|---|
| 1 | 18.010 | 5.03 | 1-Docosanol | Antidiabetic activity [17] |
| 2 | 20.441 | 27.41 | 2-Nonanol, 5-ethyl- | Anticancer activity [18] |
| 3 | 20.833 | 6.78 | Stigmasta-7,22-dien-3-ol, acetate, (3.beta., 5.alpha.,22E)- | Antiulcerogenic and antithrombotic activities [19] |
| 4 | 21.065 | 20.73 | Octadecanoic acid, 2-oxo-, methyl ester | Antibacterial [20] and antidiabetic activities [21] |
| 6 | 13.516 | 41.41 | Neophytadiene | Antipyretic, analgesic and anti-inflammatory, antimicrobial, antioxidant [22], and antidiabetic activities [23] |
| 7 | 13.520 | 42.43 | Phytol | Anxiolytic activity, metabolic modulation, cytotoxic activity, antioxidant activity, induces apoptosis, antinociceptive activity, anti-inflammatory activity, immune modulation, antimicrobial effect [24], and antidiabetic activity [22,25] |
| No. | RT | % Area | Compound | Activity |
|---|---|---|---|---|
| 1 | 19.057 | 3.06 | Octadecyl vinyl ether | Antisepsis activity [26] |
| 2 | 19.380 | 7.78 | n-Tetratetracontane | Antioxidant, anti-inflammatory, antibacterial, and antiulcerogenic activities [27] |
| 3 | 19.508 | 8.32 | 3-Ethyl-5-(2′-ethylbutyl) octadecane | Antioxidant effect and anti-inflammatory activity [28] |
| 4 | 19.555 | 11.30 | n-Dotriacontane | Anticonvulsant activity [29], antioxidant activity, and stomach cramp reliever [30] |
| 5 | 23.220 | 69.53 | n-Nonadecane | Antidiabetic activity [31] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gazali, M.; Jolanda, O.; Husni, A.; Nurjanah; Majid, F.A.A.; Zuriat; Syafitri, R. In Vitro α-Amylase and α-Glucosidase Inhibitory Activity of Green Seaweed Halimeda tuna Extract from the Coast of Lhok Bubon, Aceh. Plants 2023, 12, 393. https://doi.org/10.3390/plants12020393
Gazali M, Jolanda O, Husni A, Nurjanah, Majid FAA, Zuriat, Syafitri R. In Vitro α-Amylase and α-Glucosidase Inhibitory Activity of Green Seaweed Halimeda tuna Extract from the Coast of Lhok Bubon, Aceh. Plants. 2023; 12(2):393. https://doi.org/10.3390/plants12020393
Chicago/Turabian StyleGazali, Mohamad, Odi Jolanda, Amir Husni, Nurjanah, Fadzilah Adibah Abd Majid, Zuriat, and Rina Syafitri. 2023. "In Vitro α-Amylase and α-Glucosidase Inhibitory Activity of Green Seaweed Halimeda tuna Extract from the Coast of Lhok Bubon, Aceh" Plants 12, no. 2: 393. https://doi.org/10.3390/plants12020393