
Ranunculus sceleratus as a Model Species to Decrypt the Role of Ethylene in Plant Adaptation to Salinity

Abstract
:
1. Introduction
2. Materials and Methods
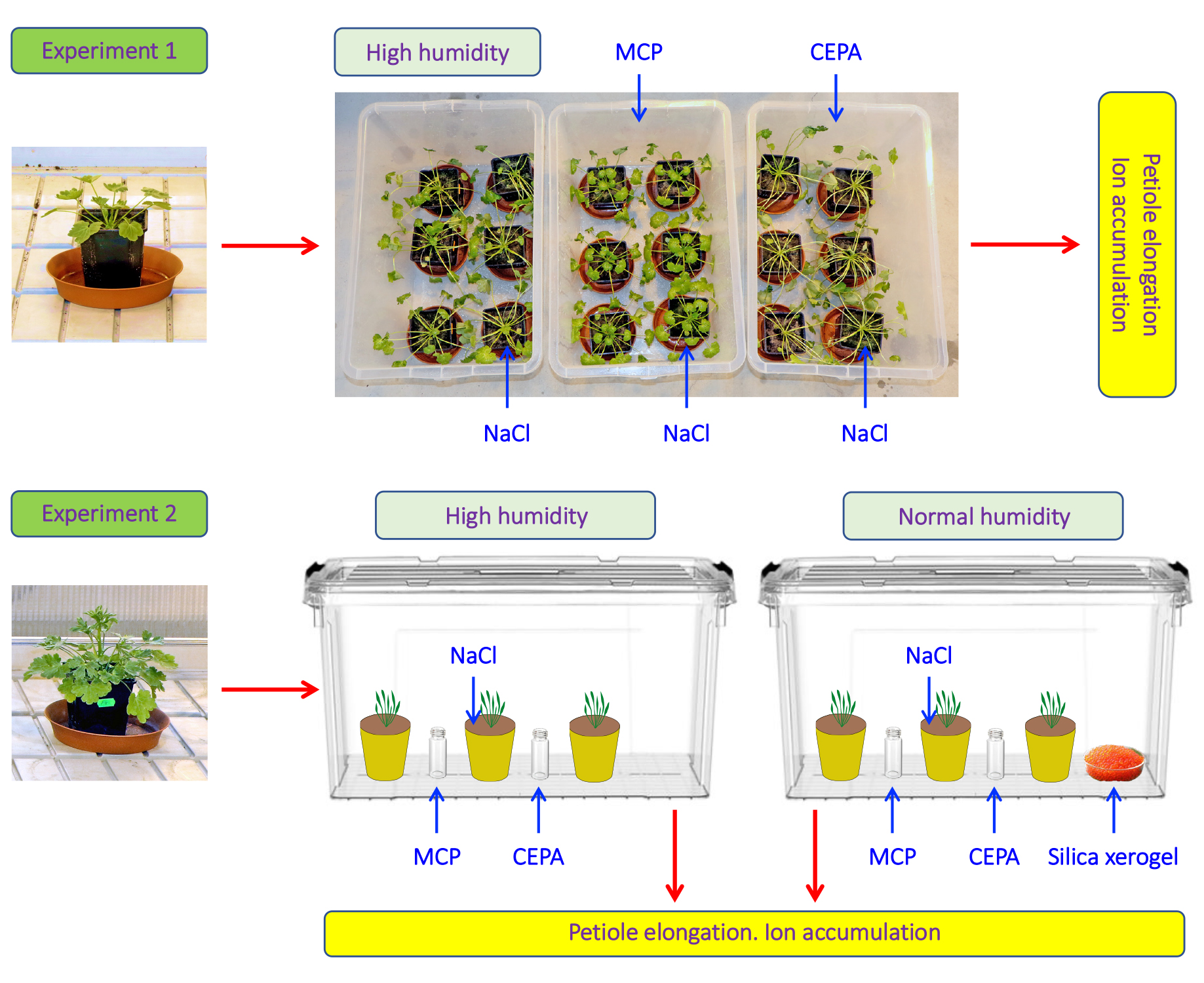
2.1. Experimental Setup
2.2. Plant Material and Cultivation Conditions
2.3. Treatments
2.4. Measurements
2.5. Data Analysis
3. Results
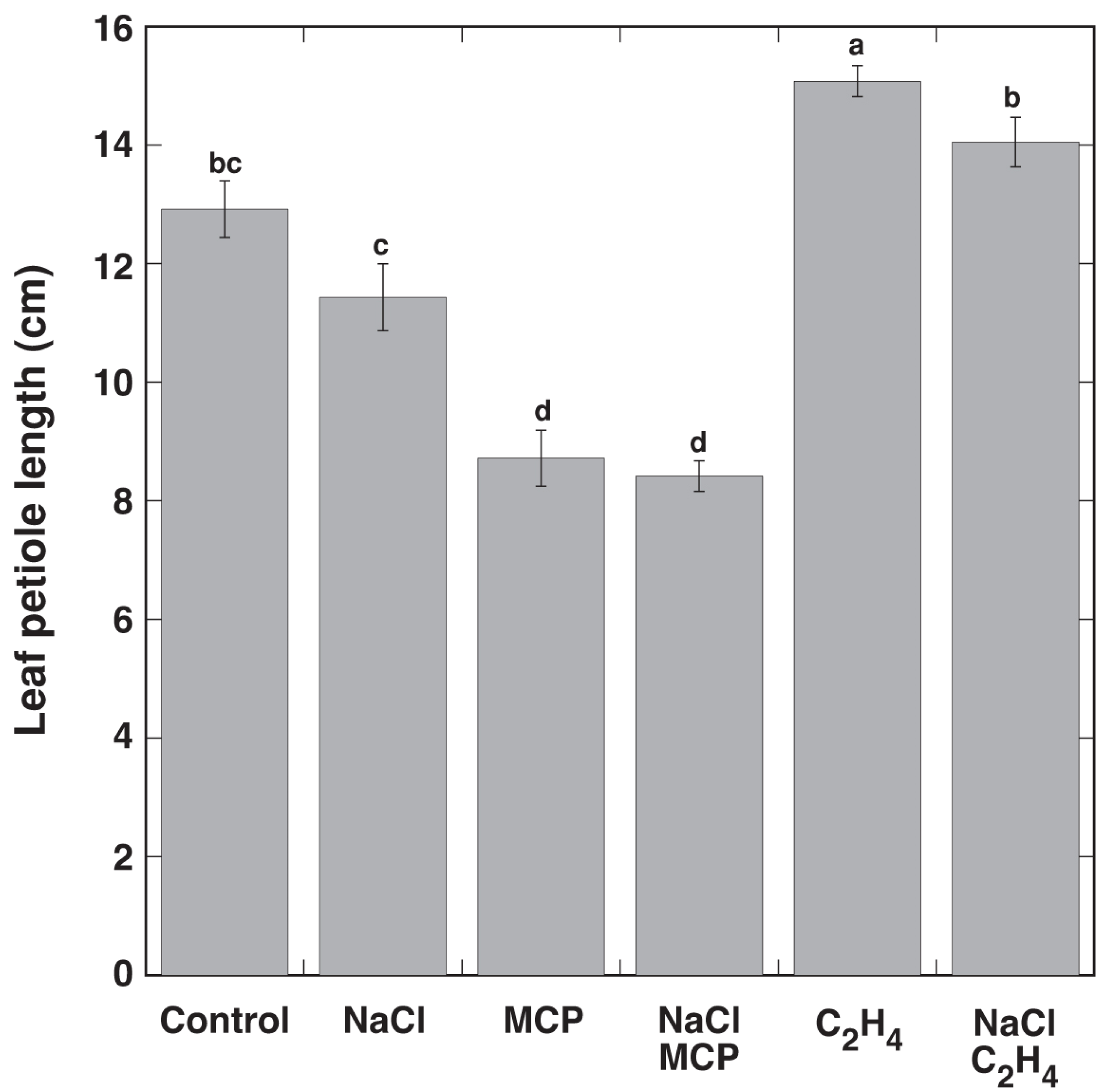
3.1. Experiment 1
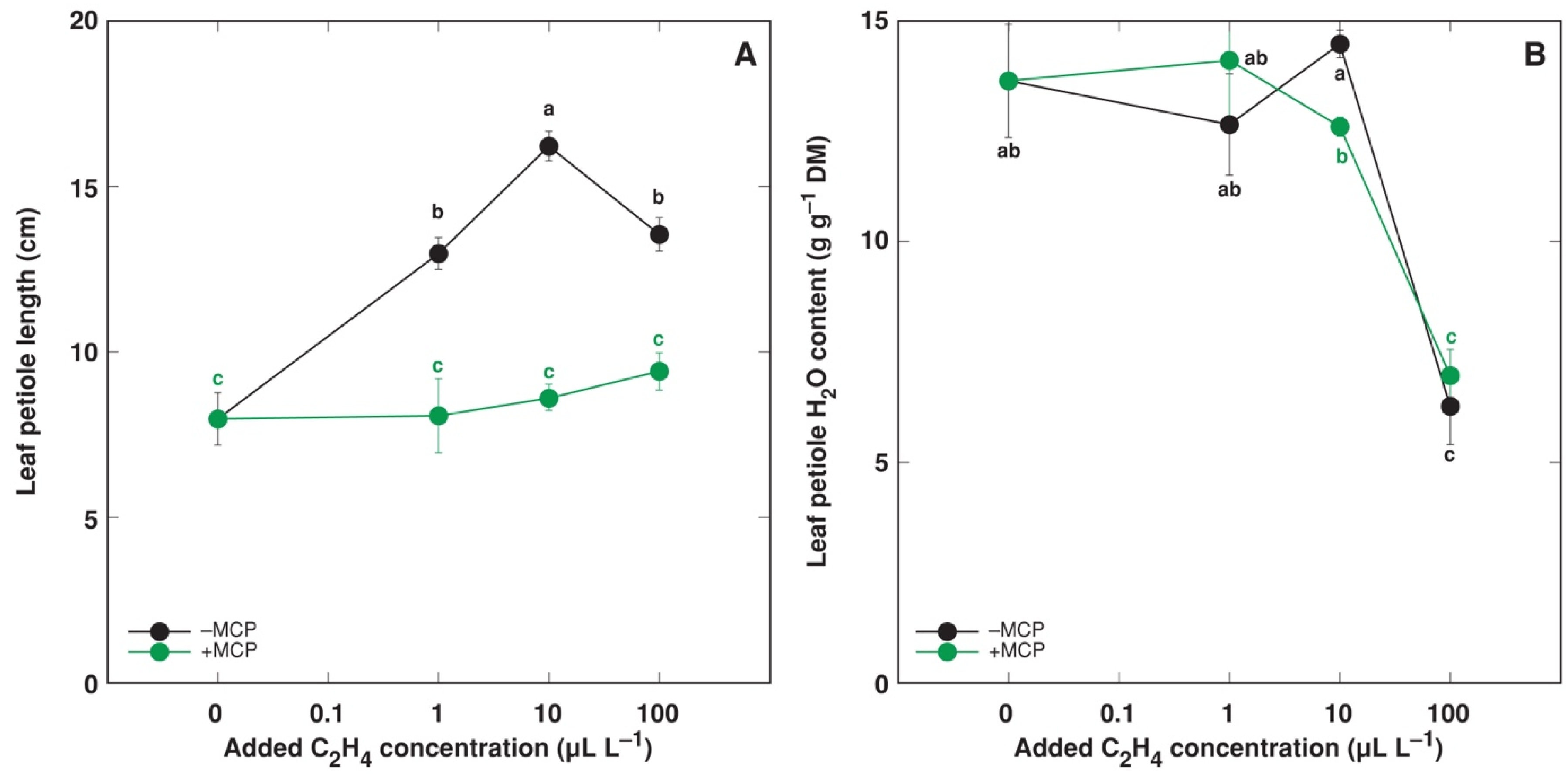
3.2. Calibration of the System
3.3. Experiment 2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, M.; Yang, Z.; Liu, J.; Zhu, T.; Wei, X.; Fan, H.; Wang, B. Adaptation Mechanism of Salt Excluders under Saline Conditions and Its Applications. Int. J. Mol. Sci. 2018, 19, 3668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.; Mostofa, M.G.; Keya, S.S.; Siddiqui, N.; Ansary, M.U.; Das, A.K.; Rahman, A.; Tran, L.S.-P. Adaptive Mechanisms of Halophytes and Their Potential in Improving Salinity Tolerance in Plants. Int. J. Mol. Sci. 2021, 22, 10733. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, F.J.; Reineke, D.; Leventini, D.; Chen, C.C.L.; Barrett-Lennard, E.G.; Colmer, T.D.; Dodd, I.C.; Shabala, S.; Brown, P.; Bazihizina, N. Plant responses to heterogeneous salinity: Agronomic relevance and research priorities. Ann. Bot. 2022, 129, 499–518. [Google Scholar] [CrossRef]
- Belkheiri, O.; Mulas, M. The effects of salt stress on growth, water relations and ion accumulation in two halophyte Atriplex species. Environ. Exp. Bot. 2013, 86, 17–28. [Google Scholar] [CrossRef]
- Dugardeyn, J.; Van Der Straeten, D. Ethylene: Fine-tuning plant growth and development by stimulation and inhibition of elongation. Plant Sci. 2008, 175, 59–70. [Google Scholar] [CrossRef]
- Chen, H.; Bullock, D.A.; Alonso, J.M.; Stepanova, A.N. To Fight or to Grow: The Balancing Role of Ethylene in Plant Abiotic Stress Responses. Plants 2021, 11, 33. [Google Scholar] [CrossRef]
- Tao, J.-J.; Chen, H.-W.; Ma, B.; Zhang, W.-K.; Chen, S.-Y.; Zhang, J.-S. The Role of Ethylene in Plants Under Salinity Stress. Front. Plant Sci. 2015, 6, 1059. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, N.; Umar, S.; Per, T.S.; Khan, N.A. Ethephon increases photosynthetic-nitrogen use efficiency, proline and antioxidant metabolism to alleviate decrease in photosynthesis under salinity stress in mustard. Plant Signal. Behav. 2017, 12, e1297000. [Google Scholar] [CrossRef]
- Pan, Y.-J.; Liu, L.; Lin, Y.-C.; Zu, Y.-G.; Li, L.-P.; Tang, Z.-H. Ethylene Antagonizes Salt-Induced Growth Retardation and Cell Death Process via Transcriptional Controlling of Ethylene-, BAG- and Senescence-Associated Genes in Arabidopsis. Front. Plant Sci. 2016, 7, 696. [Google Scholar] [CrossRef] [Green Version]
- Shibli, R.A.; Kushad, M.; Yousef, G.G.; Lila, M.A. Physiological and biochemical responses of tomato microshoots to induced salinity stress with associated ethylene accumulation. Plant Growth Regul. 2007, 51, 159–169. [Google Scholar] [CrossRef]
- Zapata, P.J.; Serrano, M.; García-Legaz, M.F.; Pretel, M.T.; Botella, M.A. Short Term Effect of Salt Shock on Ethylene and Polyamines Depends on Plant Salt Sensitivity. Front. Plant Sci. 2017, 8, 855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ellouzi, H.; Ben Hamed, K.; Hernández, I.; Cela, J.; Müller, M.; Magné, C.; Abdelly, C.; Munné-Bosch, S. A comparative study of the early osmotic, ionic, redox and hormonal signaling response in leaves and roots of two halophytes and a glycophyte to salinity. Planta 2014, 240, 1299–1317. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Charles, T.C.; Glick, B.R. Amelioration of high salinity stress damage by plant growth-promoting bacterial endophytes that contain ACC deaminase. Plant Physiol. Biochem. 2014, 80, 160–167. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Shelke, G.M.; Kumar, A.; Jha, P.N. Biochemistry and genetics of ACC deaminase: A weapon to “stress ethylene” produced in plants. Front. Microbiol. 2015, 6, 937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, W.-H.; Liu, J.; He, X.-J.; Mu, R.-L.; Zhou, H.-L.; Chen, S.-Y.; Zhang, J.-S. Modulation of Ethylene Responses Affects Plant Salt-Stress Responses. Plant Physiol. 2006, 143, 707–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Zu, Y.-G.; Tang, Z.-H. Ethylene improves Arabidopsis salt tolerance mainly via retaining K+ in shoots and roots rather than decreasing tissue Na+ content. Environ. Exp. Bot. 2013, 86, 60–69. [Google Scholar] [CrossRef]
- Zhang, W.; Wen, C.-K. Preparation of ethylene gas and comparison of ethylene responses induced by ethylene, ACC, and ethephon. Plant Physiol. Biochem. 2010, 48, 45–53. [Google Scholar] [CrossRef]
- Tassoni, A.; Watkins, C.B.; Davies, P.J. Inhibition of ethylene response by 1-MCP in tomato suggests that polyamines are not involved in delaying ripening, but may moderate the rate of ripening or over-ripening. J. Exp. Bot. 2006, 57, 3313–3325. [Google Scholar] [CrossRef]
- Watkins, C.B. The use of 1-methylcyclopropene (1-MCP) on fruits and vegetables. Biotechnol. Adv. 2006, 24, 389–409. [Google Scholar] [CrossRef]
- Acuña, M.G.V.; Biasi, W.V.; Mitcham, E.J.; Holcroft, D. Fruit temperature and ethylene modulate 1-MCP response in ‘Bartlett’ pears. Postharvest Biol. Technol. 2011, 60, 17–23. [Google Scholar] [CrossRef]
- Hays, D.B.; Do, J.H.; Mason, R.E.; Morgan, G.; Finlayson, S.A. Heat stress induced ethylene production in developing wheat grains induces kernel abortion and increased maturation in a susceptible cultivar. Plant Sci. 2007, 172, 1113–1123. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Prasad, P.; Al-Khatib, K. Ethylene perception inhibitor 1-MCP decreases oxidative damage of leaves through enhanced antioxidant defense mechanisms in soybean plants grown under high temperature stress. Environ. Exp. Bot. 2011, 71, 215–223. [Google Scholar] [CrossRef]
- Dubois, V.; Moritz, T.; Garcia-Martinez, J.L. Comparison of the role of gibberellins and ethylene in response to submergence of two lowland rice cultivars, Senia and Bomba. J. Plant Physiol. 2011, 168, 233–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, M.C.H.; Benschop, J.J.; Vreeburg, R.A.M.; Wagemaker, C.A.M.; Moritz, T.; Peeters, A.J.M.; Voesenek, A.C.J. The roles of ethylene, auxin, abscisic acid, and gibberellin in the hyponastic growth of submerged Rumex palustris petioles. Plant Physiol. 2004, 136, 2948–2960. [Google Scholar] [CrossRef] [Green Version]
- Bufler, G. Exogenous ethylene inhibits sprout growth in onion bulbs. Ann. Bot. 2008, 103, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, G.; Chang, J.; Liu, J.; Cai, J.; Rao, X.; Zhang, L.; Zhong, J.; Xie, J.; Zhu, S. Effects of 1-MCP and ethylene on expression of three CAD genes and lignification in stems of harvested Tsai Tai (Brassica chinensis). Food Chem. 2010, 123, 32–40. [Google Scholar] [CrossRef]
- Cheema, M.; Rees, D.; Colgan, R.; Taylor, M.; Westby, A. The effects of ethylene, 1-MCP and AVG on sprouting in sweetpotato roots. Postharvest Biol. Technol. 2013, 85, 89–93. [Google Scholar] [CrossRef]
- Laime, B. 1310 Salicornia and other annuals colonising mud and sand. In European Union Protected Habitats in Latvia. Interpretation Manual; Auniņš, A., Ed.; Latvian Fund for Nature; Ministry of Environmental Protection and Regional Development: Riga, Latvia, 2013; pp. 52–54. [Google Scholar]
- Rove, I. 1640 Boreal Baltic sandy beaches with perennial vegetation. In European Union Protected Habitats in Latvia. Interpretation Manual; Auniņš, A., Ed.; Latvian Fund for Nature; Ministry of Environmental Protection and Regional Development: Riga, Latvia, 2013; pp. 58–61. [Google Scholar]
- Smulders, M.J.M.; Horton, R.F. Ethylene Promotes Elongation Growth and Auxin Promotes Radial Growth in Ranunculus sceleratus Petioles. Plant Physiol. 1991, 96, 806–811. [Google Scholar] [CrossRef] [Green Version]
- Ievinsh, G.; Landorfa-Svalbe, Z.; Andersone-Ozola, U.; Karlsons, A.; Osvalde, A. Salinity and heavy metal tolerance, and phytoextraction potential of Ranunculus sceleratus plants from a sandy coastal beach. Life 2022, 12, 1959. [Google Scholar] [CrossRef]
- Nanthachai, N.; Ratanachinakorn, B.; Kosittrakun, M.; Beaudry, R.M. Absorption of 1-MCP by fresh produce. Postharvest Biol. Technol. 2007, 43, 291–297. [Google Scholar] [CrossRef]
- Blankenship, S.M.; Dole, J.M. 1-Methylcyclopropene: A review. Postharvest Biol. Technol. 2003, 28, 1–25. [Google Scholar] [CrossRef]
- Pierik, R.; Tholen, D.; Poorter, H.; Visser, E.J.; Voesenek, L.A. The Janus face of ethylene: Growth inhibition and stimulation. Trends Plant Sci. 2006, 11, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Kende, H.; van der Knaap, E.; Cho, H.-T. Deepwater Rice: A Model Plant to Study Stem Elongation. Plant Physiol. 1998, 118, 1105–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukao, T.; Bailey-Serres, J. Ethylene—A key regulator of submergence responses in rice. Plant Sci. 2008, 175, 43–51. [Google Scholar] [CrossRef]
- Voesenek, L.A.C.J.; Benschop, J.J.; Bou, J.; Cox, M.C.H.; Groeneveld, H.W.; Millenaar, F.F.; Vreeburg, R.A.M.; Peeters, A.J.M. Interactions between plant hormones regulate submergence-induced elongation in the flooding tolerant dicot Rumex palustris. Ann. Bot. 2003, 91, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Musgrave, A.; Walters, J. Ethylene-stimulated growth and auxin transport in Ranunculus sceleratus petioles. New Phytol. 1973, 72, 783–789. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Matloob, A.; Khan, F.A.; Khaliq, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.; Ullah, N.; et al. Phytohormones and plant responses to salinity stress: A review. Plant Growth Regul. 2014, 75, 391–404. [Google Scholar] [CrossRef]
- Gharbi, E.; Martínez, J.-P.; Benahmed, H.; Lepoint, G.; Vanpee, B.; Quinet, M.; Lutts, S. Inhibition of ethylene synthesis reduces salt-tolerance in tomato wild relative species Solanum chilense. J. Plant Physiol. 2017, 210, 24–37. [Google Scholar] [CrossRef]
- Cuin, T.A.; Betts, S.A.; Chalmandrier, R.; Shabala, S. A root’s ability to retain K+ correlates with salt tolerance in wheat. J. Exp. Bot. 2008, 59, 2697–2706. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Wang, H.; Han, B.; Wang, B.; Guo, A.; Zheng, D.; Liu, C.; Chang, L.; Peng, M.; Wang, X. Sodium instead of potassium and chloride is an important macronutrient to improve leaf succulence and shoot development for halophyte Sesuvium portulacastrum. Plant Physiol. Biochem. 2012, 51, 53–62. [Google Scholar] [CrossRef]
- Kronzucker, H.J.; Coskun, D.; Schulze, L.M.; Wong, J.R.; Britto, D.T. Sodium as nutrient and toxicant. Plant Soil 2013, 369, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Ievinsh, G.; Ieviņa, S.; Andersone-Ozola, U.; Samsone, I. Leaf sodium, potassium and electrolyte accumulation capacity of plant species from salt-affected coastal habitats of the Baltic Sea: Towards a definition of Na hyperaccumulation. Flora 2020, 274, 151748. [Google Scholar] [CrossRef]
- He, J.B.; Bögemann, G.M.; Van De Steeg, H.M.; Rijnders, J.G.H.M.; Voesenek, L.A.C.J.; Blom, C.W.P.M. Survival tactics of Ranunculus species in river floodplains. Oecologia 1999, 118, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Park, W.J.; Hertel, R.; Kang, B.G. Enhancement of auxin sensitivity in Ranunculus sceleratus by ethylene: A mechanism to escape from hypoxia under temporary submergence. Environ. Exp. Bot. 2011, 72, 266–271. [Google Scholar] [CrossRef]
- Shiono, K.; Takahashi, H.; Colmer, T.D.; Nakazono, M. Role of ethylene in acclimations to promote oxygen transport in roots of plants in waterlogged soils. Plant Sci. 2008, 175, 52–58. [Google Scholar] [CrossRef]
- Schaller, G.E.; Binder, B.M. Inhibitors of ethylene biosynthesis and signaling. In Ethylene Signaling: Methods and Protocols; Binder, B.M., Schaller, G.E., Eds.; Methods in Molecular biology; Springer Science+Business Media: Berlin/Heidelberg, Germany, 2017; Volume 1573, pp. 223–234. [Google Scholar]
- Mortensen, L.M.; Giselrød, H.R. Effects of air humidity and supplementary lighting on foliage plants. Sci. Hortic. 1990, 44, 301–308. [Google Scholar] [CrossRef]
- Rosenvald, K.; Tullus, A.; Ostonen, I.; Uri, V.; Kupper, P.; Aosaar, J.; Varik, M.; Sõber, J.; Niglas, A.; Hansen, R.; et al. The effect of elevated air humidity on young silver birch and hybrid aspen biomass allocation and accumulation—Acclimation mechanisms and capacity. For. Ecol. Manag. 2014, 330, 252–260. [Google Scholar] [CrossRef]
- Koch, K.; Hartmann, K.D.; Schreiber, L.; Barthlott, W.; Neinhuis, C. Influences of air humidity during the cultivation of plants on wax chemical composition, morphology and leaf surface wettability. Environ. Exp. Bot. 2006, 56, 1–9. [Google Scholar] [CrossRef]
- Sellin, A.; Alber, M.; Kupper, P. Increasing air humidity influences hydraulic efficiency but not functional vulnerability of xylem in hybrid aspen. J. Plant Physiol. 2017, 219, 28–36. [Google Scholar] [CrossRef]
- Calvo-Polanco, M.; Ibort, P.; Molina, S.; Ruiz-Lozano, J.M.; Zamarreño, A.M.; García-Mina, J.M.; Aroca, R. Ethylene sensitivity and relative air humidity regulate root hydraulic properties in tomato plants. Planta 2017, 246, 987–997. [Google Scholar] [CrossRef]
- Fanourakis, D.; Hyldgaard, B.; Giday, H.; Bouranis, D.; Körner, O.; Nielsen, K.L.; Ottosen, C.-O. Differential effects of elevated air humidity on stomatal closing ability of Kalanchoë blossfeldiana between the C3 and CAM states. Environ. Exp. Bot. 2017, 143, 115–124. [Google Scholar] [CrossRef]
- Georgii, E.; Jin, M.; Zhao, J.; Kanawati, B.; Schmitt-Kopplin, P.; Albert, A.; Winkler, J.B.; Schäffner, A.R. Relationships between drought, heat and air humidity responses revealed by transcriptome-metabolome co-analysis. BMC Plant Biol. 2017, 17, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, W.; Yang, Z.; Huang, L.; Sun, C.; Yu, X.; Zhao, M. Fuzzy comprehensive evaluation of the effects of relative air humidity on the morpho-physiological traits of Pakchoi (Brassica chinensis L.) under high temperature. Sci. Hortic. 2018, 246, 971–978. [Google Scholar] [CrossRef]
- Rodrigues, C.R.F.; Silveira, J.A.G.; Viégas, R.A.; Moura, R.M.; Aragão, R.M.; Silva, E.N. Combined effects of high relative humidity and K+ supply mitigates damage caused by salt stress on growth, photosynthesis and ion homeostasis in J. curcas plants. Agric. Waste Manag. 2016, 163, 255–262. [Google Scholar] [CrossRef]
- Backhausen, J.E.; Klein, M.; Klocke, M.; Jung, S.; Scheibe, R. Salt tolerance of potato (Solanum tuberosum L. var. Desirée) plants depends on light intensity and air humidity. Plant Sci. 2005, 169, 229–237. [Google Scholar] [CrossRef]
- Carvalho, D.R.; Vasconcelos, M.W.; Lee, S.; Vreugdenhil, D.; Heuvelink, E.; Carvalho, S.M. Moderate salinity improves stomatal functioning in rose plants grown at high relative air humidity. Environ. Exp. Bot. 2017, 143, 1–9. [Google Scholar] [CrossRef]
- Poorter, H.; Fiorani, F.; Stitt, M.; Schurr, U.; Finck, A.; Gibon, Y.; Usadel, B.; Munns, R.; Atkin, O.; Tardieu, F.; et al. The art of growing plants for experimental purposes: A practical guide for the plant biologist. Funct. Plant Biol. 2012, 39, 821–838. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Hernández, M.C.; Garmendia, I. Age of plant influences the effect of salinity in yield and mineral content in ice plants. J. Appl. Bot. Food Qual. 2022, 95, 94–99. [Google Scholar]
- Samarakoon, A.B.; Hotron, R.F. Flowering and stem growth in the celery-leaved buttercup, Ranunculus sceleratus. Can. J. Bot. 1981, 59, 1386–1392. [Google Scholar] [CrossRef]
- Ashraf, A.M.; Begam, S.N.; Ragavan, T. Defoliants Harvest-aid Chemicals: Cost Effective Technology to Facilitate Synchronized Maturity for Mechanical Harvesting in Cotton: A Review. Agric. Rev. 2021, R-2081. [Google Scholar] [CrossRef]
- Rutkowski, K.; Łysiak, G.P. Thinning Methods to Regulate Sweet Cherry Crops—A Review. Appl. Sci. 2022, 12, 1280. [Google Scholar] [CrossRef]
- Zhang, W.; Hu, W.; Wen, C.-K. Ethylene preparation and its application to physiological experiments. Plant Signal. Behav. 2010, 5, 453–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, F.; Mahfouz, S. Effect of 1-methylcyclopropene (1-MCP) on the postharvest senescence of coriander leaves during storage and its relation to antioxidant enzyme activity. Sci. Hortic. 2012, 141, 69–75. [Google Scholar] [CrossRef]
- Tiwari, K.; Paliyath, G. Microarray analysis of ripening-regulated gene expression and its modulation by 1-MCP and hexanal. Plant Physiol. Biochem. 2011, 49, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Mata, C.I.; Magpantay, J.; Hertog, M.L.A.T.M.; Van de Poel, B.; Nicolaï, B.M. Expression and protein levels of ethylene receptors, CTRs and EIN2 during tomato fruit ripening as affected by 1-MCP. Postharv. Biol. Technol. 2021, 179, 111573. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment/Treatment | NaCl | MCP | CEPA | Silica Xerogel | Container Volume (L) | Replicates (Boxes/ Plants) |
|---|---|---|---|---|---|---|
| Experiment 1 (high humidity) | ||||||
| Control | − | − | − | − | 48 | 2/6 |
| NaCl | + | − | − | − | 48 | 2/6 |
| MCP | − | + | − | − | 48 | 2/6 |
| NaCl MCP | + | + | − | − | 48 | 2/6 |
| C2H4 | − | − | + | − | 48 | 2/6 |
| NaCl C2H4 | + | − | + | − | 48 | 2/6 |
| Experiment 2 (high humidity) | ||||||
| Control | − | − | − | − | 48 | 2/6 |
| NaCl | + | − | − | − | 48 | 2/6 |
| NaCl C2H4 | + | − | + | − | 48 | 2/6 |
| NaCl MCP | + | + | − | − | 48 | 2/6 |
| NaCl MCP C2H4 | + | + | + | − | 48 | 2/6 |
| Experiment 2 (normal humidity) | ||||||
| Control | − | − | − | + | 24 | 3/6 |
| NaCl | + | − | − | + | 24 | 3/6 |
| NaCl C2H4 | + | − | + | + | 24 | 3/6 |
| NaCl MCP | + | + | − | + | 24 | 3/6 |
| NaCl MCP C2H4 | + | + | + | + | 24 | 3/6 |
| Treatment | High Humidity | Normal Humidity | ||
|---|---|---|---|---|
| In Respect to Control (%) | Statistical Significance (p) | In Respect to Control (%) | Statistical Significance (p) | |
| Leaf petiole length | ||||
| NaCl | 75 | <0.05 | 80 | <0.05 |
| NaCl C2H4 | 112 | ns | 102 | ns |
| NaCl MCP | 81 | <0.05 | 75 | <0.05 |
| NaCl MCP C2H4 | 78 | <0.05 | 89 | ns |
| Leaf petiole dry mass | ||||
| NaCl | 109 | ns | 100 | ns |
| NaCl C2H4 | 147 | <0.05 | 141 | ns |
| NaCl MCP | 113 | ns | 101 | ns |
| NaCl MCP C2H4 | 137 | <0.05 | 134 | ns |
| Leaf blade dry mass | ||||
| NaCl | 132 | ns | 116 | ns |
| NaCl C2H4 | 123 | ns | 117 | ns |
| NaCl MCP | 137 | ns | 119 | ns |
| NaCl MCP C2H4 | 175 | <0.05 | 149 | <0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prokopoviča, V.; Ievinsh, G. Ranunculus sceleratus as a Model Species to Decrypt the Role of Ethylene in Plant Adaptation to Salinity. Plants 2023, 12, 370. https://doi.org/10.3390/plants12020370
Prokopoviča V, Ievinsh G. Ranunculus sceleratus as a Model Species to Decrypt the Role of Ethylene in Plant Adaptation to Salinity. Plants. 2023; 12(2):370. https://doi.org/10.3390/plants12020370
Chicago/Turabian StyleProkopoviča, Veronika, and Gederts Ievinsh. 2023. "Ranunculus sceleratus as a Model Species to Decrypt the Role of Ethylene in Plant Adaptation to Salinity" Plants 12, no. 2: 370. https://doi.org/10.3390/plants12020370